Страйер Л. Биохимия. Том 3
Подождите немного. Документ загружается.


ковые пробирки с образцами протыкали
снизу и собирали фракции по каплям для
дальнейшего анализа.
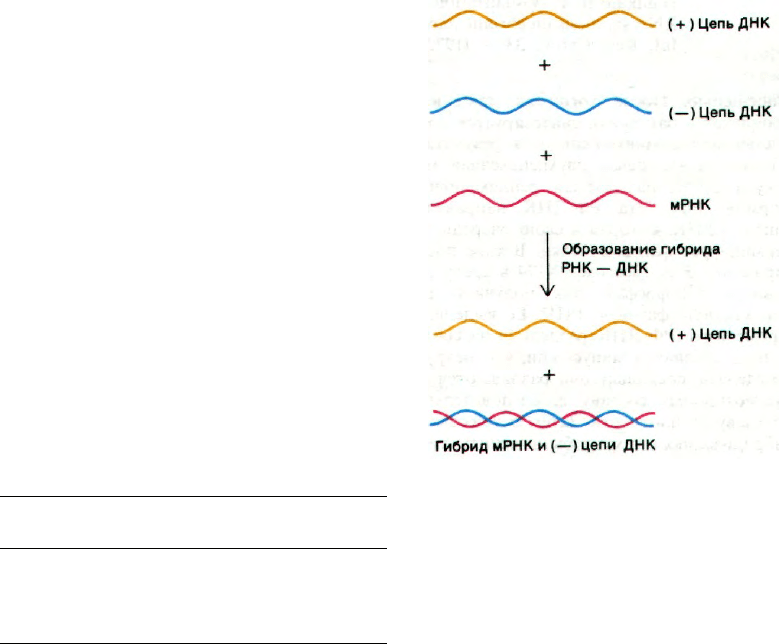
В результате были обнаружены три по-
лосы (рис. 25.6). Полоса с наибольшей
плотностью соответствовала одноцепочеч-
ной РНК. Вторая полоса соответствовала
двухспиральной ДНК. Третья располага-
лась вблизи от полосы ДНК и состояла из
двухцепочечных гибридных молекул
ДНК—РНК. Итак, Т2-мРНК образовыва-
ла гибрид с Т2-ДНК. В отличие от этого
Т2-РНК не гибридизовалась с ДНК мно-
гих бактерий и неродственных вирусов, да-
же если их нуклеотидный состав был подо-
бен Т2-ДНК. Последующие эксперименты
показали, что фракция мРНК из незара-
женных клеток гибридизуется с ДНК
именно того организма, из которого она
была выделена, но не с ДНК нерод-
ственных организмов. Эти убедительные
эксперименты продемонстрировали, что
последовательность оснований мРНК ком-
плементарна последовательности ДНК-ма-
трицы. К тому же был разработан
мощный метод, с помощью которого мож-
но было исследовать поток генетической
информации в клетках и выяснять, сходны
ли две молекулы нуклеиновой кислоты.
25.6. Рибосомные РНК и транспортные
РНК также синтезируются на
ДНК-матрице
Метод гибридизации был использован за-
тем, чтобы выяснить, синтезируются ли
рРНК и тРНК также на ДНК-матрицах.
Образование гибридов РНК—ДНК выяв-
ляли с помощью фильтров, а не центрифу-
гированием в градиенте плотности, так как
этот метод проще, чувствительнее и тре-
бует меньше времени. Одноцепочечные
РНК проходят через нитроцеллюлозный
фильтр, а двухспиральные ДНК и гибриды
РНК—ДНК задерживаются на фильтре.
РНК Е. coli пометили изотопом
32
Р и сме-
шали с немеченой ДНК Е. coli. Эту смесь
нагревали, медленно охлаждали и затем
отфильтровывали через нитроцеллюлозу.
Радиоактивность, задержанную на филь-
тре, просчитывали. Результаты экспери-
ментов не вызывали сомнений: гибриды
РНК—ДНК образовывались со всеми
рРНК (5S, 16S и 23S) и тРНК. Отсюда сле-
довало, что в геноме Е. coli имеются после-
довательности, комплементарные этим мо-
лекулам РНК.
Рис. 25.6. РНК, образовавшаяся после
заражения E. coli фагом Т2,
комплементарна вирусной
ДНК. В этих экспериментах по
гибридизации РНК метили
32
Р, а ДНК фага Т2 -
3
Н. Рас-
пределение радиоактивности
в градиенте плотности хлори-
стого цезия показывает, что
большая часть РНК, синтези-
рованной после заражения, по-
падает в одну полосу с ДНК
фага Т2. (Spiegelman S., Hybrid
nucleic acids, Sci. Amer., 1964.)
Рис. 25.7. Электронная микрофотогра-
фия РНК-полимеразы E. coli.
(Печатается с любезного разре-
шения д-ра Robley Williams
и д-ра Michael Chemberlin.)
25. Информационная РНК
и транскрипция
51

Рис. 25.8. Механизм реакции элонгации
цепи, катализируемой РНК-по-
лимеразой.
25.7. Все клеточные РНК синтезирует
РНК-полимеразa
Концепция мРНК стимулировала поиски
фермента, который синтезирует РНК в со-
ответствии с последовательностью ДНК-
матрицы. Стратегия эксперимента была
такой же, как и при поиске ДНК-полиме-
разы I. В 1960г. Джерард Хёрвиц и Сэ-
мюэл Вейсс (Jerard Hurwitz, Samuel Weiss)
независимо открыли такой фермент. Они
назвали его РНК-полимеразой. Ферменту
из клеток Е. соli (рис. 25.7) нужны были
для синтеза РНК следующие компоненты.
1. Матрица. Предпочтительная матри-
ца - двухцепочечная ДНК. Одноцепочечная
ДНК также может служить матрицей. Ни
РНК (как одноцепочечная, так и двухцепо-
чечная), ни гибриды РНК—ДНК не могут
служить эффективными матрицами.
2. Активированные предшественники. Не-
обходимы все четыре рибонуклеозидтри-
фосфата - ATР, GTP, UTP и СТР.
3. Двухвалентные ионы металлов. Фер-
мент активен в присутствии Mg
2+
или
Mn
2+
. In vivo эту потребность фермента
удовлетворяет Mg
2+
.
РНК-полимераза катализирует инициа-
цию и элонгацию цепей РНК. Фермент ка-
тализирует следующую реакцию:
(РНК)
n
остатков
+
Рибонуклеозидтрифос-
фат
(РНК)
n
+ 1
остаток
+
PP
i
.
52
Часть IV.
Информация
Синтез РНК во многих отношениях
подобен синтезу ДНК (рис. 25.8). Во-
первых, как будет вскоре показано более
подробно, синтез идет в направлении 5'—>
—>3'. Во-вторых, по-видимому, механизм
элонгации сходен. Происходит нуклео-
фильная атака внутреннего фосфата оче-
редного нуклеозидтрифосфата 3'-ОН-груп-
пой на конце растущей цепи. В-третьих,
движущая сила синтеза - гидролиз пиро-
фосфата.
Однако по некоторым важным особен-
ностям синтез РНК отличается от синтеза
ДНК. Во-первых, РНК-полимеразе не нуж-
на затравка. Во-вторых, ДНК-матрица при
синтезе РНК полностью сохраняется, тог-
да как при синтезе ДНК она сохраняется
лишь наполовину. В-третьих, РНК-полиме-
раза, насколько известно, не обладает ни-
какими нуклеазными активностями.
Все три типа клеточной РНК Е.
coli - мРНК, тРНК и рРНК - синтезируют-
ся одной РНК-полимеразой в соответствии
с инструкциями, заданными ДНК-матри-
цей. В клетках млекопитающих имеет ме-
сто разделение труда между несколькими
видами РНК-полимераз. Кроме того, необ-
ходимо отметить, что некоторые вирусы
кодируют РНК-синтезирующие ферменты,
совершенно отличные от ферментов клет-
ки-хозяина. Таковы, например, РНК-поли-
мераза, кодируемая ДНК-содержащим фа-
гом Т7, и РНК-репликаза, кодируемая
РНК-содержащим фагом Qβ. Репликаза
фага Qβ является РНК-зависимой РНК-
полимеразой, так как она синтезирует
РНК не по ДНК-матрице, а по РНК
(гл. 30). Клеточные же ферменты, синтези-
5'-GCGGCGACGCGCAGUUAAUCCCACAGCCGCCAGUUCCGCUGGCGGCAUUUU-3' мРНК
3'-CGCCGCTGCGCGTCAATTAGGGTGTCGGCGGTCAAGGCGACCGCCGTAAAA-5'
ДНК
5'-GCGGCGACGCGCAGTTAATCCCACAGCCGCCAGTTCCGCTGGCGGCATTTT-3'
Рис. 25.9. Последовательность основа-
ний мРНК комплементарна
последовательности матрич-
ной ДНК. Показана часть по-
следовательности триптофано-
вого оперона.
рующие РНК, наоборот, представляют со-
бой ДНК-зависимые РНК-полимеразы.
25.8. РНК-полимераза получает инструкции
от ДНК-матрицы
Подобно ДНК-полимеразам, описанным
в предыдущей главе, РНК-полимераза ис-
пользует информацию, содержащуюся
в ДНК-матрице. Первым доводом в поль-
зу этого утверждения послужили данные
о том, что нуклеотидный состав новосин-
тезированной РНК комплементарен соста-
ву матричной цепи ДНК. Если в качестве
матрицы использовать синтетический по-
лидезоксирибонуклеотид poly(dT), содержа-
щий только остатки тимидиловой кислоты,
то в синтезируемую полирибонуклеотид-
ную цепь включается только один рибону-
клеозидтрифосфат - АТР. Продукт реак-
ции - полирибоаденилат [сокращенное обо-
значение poly(rA)]. Если в качестве ма-
трицы для РНК-полимеразы используется
сополимер с чередующимися основаниями
poly(dA-dT), то включаются UTP и АТР,
а в качестве продукта реакции образуется
poly(rA-rU). Нуклеотидный состав РНК,
синтезированной с использованием в каче-
стве матрицы одноцепочечной ДНК фага
φХ174, также свидетельствует о наличии
комплементарности между РНК-продук-
том и ДНК-матрицей (табл. 25.2). Экспери-
менты по гибридизации показывают, что
РНК, синтезированная РНК-полимеразой,
Таблица 25.2. Нуклеотидный состав РНК, синтезиро-
ванной на вирусной ДНК в качестве матрицы
ДНК-матрица (плюс-цепь фага
φХ174)
А
Т
G
С
0.25
0,33
0,24
0,18
РНК-продукт
0,25
0,32
0,23
0,20
U
А
С
G
комплементарна матричной ДНК. Наибо-
лее убедительный довод в пользу надежно-
сти копирования ДНК при транскрипции
получен при изучении последовательности
оснований; оказалось, что последователь-
ность мРНК в точности комплементарна
последовательности матричной ДНК
(рис. 25.9).
25.9. Обычно в данном участке генома
транскрибируется только одна цепь ДНК
Транскрибируются ли обе цепи двухспи-
ральной матричной ДНК, или только од-
на? А priori кажется маловероятным, что
в одном и том же участке ДНК обе цепи
кодируют функциональные белки. Один из
первых ответов на этот вопрос был полу-
чен при использовании метода гибридиза-
ции в опытах с Е. coli, зараженной фагом
φХ174. Частицы фага φХ174 содержат
одноцепочечную ДНК, которую называют
Рис. 25.10. Как показывает этот гибриди-
зационный эксперимент, толь-
ко одна цепь РФ-ДНК фага
φХ174 используется в качестве
матрицы при транскрипции.
25. Информационная РНК
и транскрипция
53

Рис. 25.11. Электронная микрофотогра-
фия, на которой виден процесс
транскрипции. [Miller О. L.,
Hamkalo В. A., Visualization of
RNA synthesis on chromosomes,
Int. Rev. Cytol., 33, 1 (1972).]
плюс-цепью. После того как плюс-цепь
проникает в бактерию, синтезируется ком-
плементарная минус-цепь, и в результате
образуется кольцевая двухцепочечная мо-
лекула ДНК, называемая репликативной
формой (РФ). Эта РФ-ДНК направляет
синтез мРНК, которая в свою очередь де-
терминирует фаговые белки. Вскоре после
заряжения E. coli фагом φХ174 в среду до-
бавляли
32
Р-фосфат; так получили ра-
диоактивную фаговую РНК. Ее выделили.
Кроме того, РФ-ДНК разделили на соста-
вляющие плюс- и минус-цепи, что нетруд-
но сделать, поскольку они различаются по
нуклеотидному составу и, следовательно,
по плавучей плотности. Затем осуществили
гибридизацию, с тем чтобы выяснить, ком-
плементарна ли новосинтезированная
мРНК плюс-цепи, или минус-цепи, или же
обеим цепям. Был получен однозначный
результат: только минус-цепь РФ-ДНК
образовывала гибрид с меченой мРНК.
Итак, только одна цепь РФ-ДНК фага
54
Часть IV.
Информация
φХ174 служит in vivo матрицей для тран-
скрипции.
Точно так же только одна цепь таких ви-
русов, как Т7, SP8 и α, транскрибируется in
vivo. В случае таких вирусов, как фаги Т4
или λ, ситуация сложнее. В одних участках
генома матрицей служит одна цепь, в дру-
гих - другая. Такая же транскрипция с пере-
меной матрицы в различных группах генов
происходит и в клетках Е. coli.
25.10. РНК-полимераза Е. coli состоит из
субъединиц
РНК-полимераза Е. coli - очень большой
и сложный фермент. Масса целого фермен-
та равна примерно 500 кДа. РНК-полиме-
раза состоит из отдельных субъединиц
(табл. 25.3); это можно доказать, вызвав
диссоциацию фермента в концентрирован-
ном растворе мочевины. Субъединичный
состав целого фермента, называемого го-
лоферментом,- α
2
ββ'σ. Как это видно из
Таблица 25.3. Субъединицы РНК-полимеразы E.coli
Обозначение
субъединицы
α
β
β'
σ
Число
субъединиц
2
1
1
1
Масса, кДа .
40
155
165
95

Рис. 25.12. Электронная микрофотогра-
фия РНК-полимеразы-голо-
фермента, связанного со мно-
гими промоторными участка-
ми фрагмента ДНК фага Т7.
[Williams R.C., Ргос. Nat. Acad.
Sci., 74, 2313 (1977).]
последующего текста, после того как ини-
циация синтеза РНК произошла, σ-субъе-
диница отделяется от фермента. РНК-по-
лимераза без σ-субъединицы называется
минимальным ферментом или кор-фермен-
том (α
2
ββ'). Каталитический участок РНК-
полимеразы находится в кор-ферменте.
Установлено, что β'-субъединица участвует
в связывании с ДНК-матрицей, а β-субъе-
диница - в связывании субстратов - рибону-
клеозидтрифосфатов. σ-Субъединица голо-
фермента участвует в выборе участка
инициации транскрипции.
Синтез РНК РНК-полимеразой Е. coli
происходит в три этапа, называемых
1) инициация, 2) элонгация и 3) термина-
ция. С этими процессами мы познакомим-
ся несколько ниже.
25.11. Транскрипция инициируется на промо-
торных участках матричной ДНК
Транскрипция начинается в определенных
участках матричной ДНК, называемых
промоторами. Как РНК-полимераза нахо-
дит эти участки? Один из подходов к ре-
шению этой проблемы состоит в том,
чтобы выделить те фрагменты, которые
голофермент РНК-полимеразы защищает
от расщепления панкреатической дезокси-
рибонуклеазой, и определить в этих фраг-
ментах последовательность нуклеотидов.
Другой подход сводится к тому, чтобы ис-
следовать ряд мутантов с повышенной или
пониженной интенсивностью транскрипции
определенных генов. В результате таких
исследований было установлено, что про-
моторные участки состоят примерно из со-
рока пар оснований, что соответствует
фрагменту ДНК длиной примерно 140 А.
Как показал анализ различных промото-
ров Е. coli и фагов, функцию сигнала узна-
вания в основном выполняет последова-
тельность семи пар оснований, середина
которой находится на расстоянии пример-
но 10 нуклеотидов перед точкой, с которой
начинается кодирование мРНК (рис. 25.13).
Второй участок узнавания расположен на
расстоянии около 35 нуклеотидов перед
точкой начала мРНК; он также участвует
в первоначальном связывании РНК-поли-
меразы.
25.12. σ-Субъединица обеспечивает
узнавание промоторных участков РНК-
полимеразой
Кор-фермент (α
2
ββ') РНК-полимеразы не
может начинать транскрипцию в промо-
торных участках. Для специфической ини-
циации необходим голофермент α
2
ββ'σ
Роль сигма-субъединицы была открыта
следующим образом. Если РНК-полимера-
зу очищали хроматографией на колонке
с фосфоцеллюлозой, то она была практи-
Рис. 25.13. Промоторные последователь-
ности лактозного (А), галактоз-
ного (Б) и триптофанового (В)
оперонов E.coli, фагов λ (Г)
и φХ174 (Д) и вируса SV-40 (Е).
Предполагаемая область го-
мологии, так называемая по-
следовательность Прибнова
(Pribnow), показана зеленым.
25. Информационная РНК
и транскрипция
55
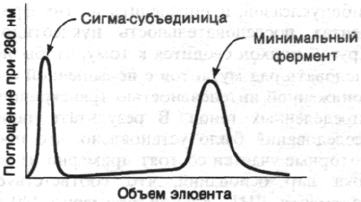
Рис. 25.14. Разделение РНК-полимеразы
на σ-субъединицу и кор-фер-
мент (минимальный фермент;
α
2
ββ')
на
колонке
с
фосфоцел-
люлозой.
чески лишена активности при использова-
нии в качестве матрицы ДНК фага Т4, но
сохраняла активность в опытах с ДНК из
тимуса теленка. Наряду с этим та же РНК-
полимераза, очищенная центрифугирова-
нием в градиенте концентрации глицерола,
была, напротив, весьма активна на обеих
матрицах. Из этого наблюдения можно
было сделать вывод, что в препарате РНК-
полимеразы, очищенной на фосфоцеллю-
лозе, не хватает какого-то фактора. Так оно
и было (рис. 25.14). Активность фермента,
очищенного на фосфоцеллюлозе, значи-
тельно увеличивается при добавлении дру-
гой фракции, элюирующейся с колонки.
Этот стимулирующий фактор сам по себе
не обладает каталитической активностью.
Он был назван сигма (σ)-фактором. После-
дующие эксперименты показали, что фер-
мент, очищенный на фосфоцеллюлозе,
лишен σ-субъединицы, тогда как фермент,
очищенный в глицероле, содержит ее. Та-
ким образом, препарат, проявлявший мак-
симальную активность при использовании
в качестве матрицы ДНК фага Т4, предста-
влял собой голофермент α
2
ββ'σ а кор-фер-
мент α
2
ββ' был не в состоянии транскриби-
ровать эту ДНК. Добавление σ-субъеди-
ницы к кор-ферменту приводило к рекон-
струкции вполне активного голофермента.
Кор-фермент обладал способностью тран-
скрибировать ДНК тимуса теленка, так
как эта матрица содержит много одноце-
почечных разрывов. РНК, синтезированная
кор-полимеразой in vitro, не соответствует
транскриптам, образующимся in vivo.
В частности, кор-фермент транскрибирует
56
Часть IV.
Информация
обе цепи фаговых ДНК-матриц, тогда как
голофермент транскрибирует асимметрич-
но одну цепь, как он это делает in vivo.
Сигма-субъединица обеспечивает специ-
фическую инициацию, снижая в 10
4
раз
сродство РНК-полимеразы к ДНК, не
содержащей промоторов. Кроме того, сиг-
ма-субъединица активирует узнавание про-
моторных последовательностей РНК-поли-
меразой. Наконец, сигма-субъединица уча-
ствует в раскрывании двойной спирали
ДНК, так чтобы одна из цепей могла слу-
жить матрицей. При связывании голофер-
мента РНК-полимеразы расплетается при-
мерно один виток спирали ДНК. Так
происходит подготовка фермента к обра-
зованию первой фосфодиэфирной связи но-
вой цепи РНК.
После того как начинается синтез новой
цепи РНК, сигма-субъединица отделяется
от голофермента. Кор-фермент продол-
жает транскрибировать матрицу ДНК. Та-
ким образом, функция голофермента —
поиск промотора и инициация, а функция
кор-фермента - элонгация. Отделившаяся
сигма-субъединица присоединяется к дру-
гой молекуле кор-полимеразы, чтобы уча-
ствовать в инициации нового цикла тран-
скрипции.
Инициация новых цепей РНК регули-
руется многими способами. Некоторые
промоторные последовательности обеспе-
чивают высокую эффективность инициа-
ции, другие менее эффективны. Кроме то-
го, эффективность инициации регулируется
белками, которые связываются с самим
промоторным участком или рядом с ним.
Репрессоры блокируют транскрипцию,
препятствуя связыванию РНК-полимеразы,
а факторы положительной регуляции спо-
собствуют инициации, облегчая связывание
РНК-полимеразы. Эти важные регуля-
торные механизмы мы рассмотрим под-
робнее в гл. 28.
25.13. Цепи РНК начинаются с pppG или
рррА
Большинство новосинтезированных цепей
РНК содержат своего рода ярлычок, ко-
торый показывает, с чего начался их син-
тез. Новые цепи РНК имеют строго опре-
деленную структуру 5'-конца: молекула
начинается либо с рррА, либо с pppG. В от-
личие от синтеза ДНК затравка в этом
случае не нужна. Цепи РНК могут обра-
зовываться de novo.
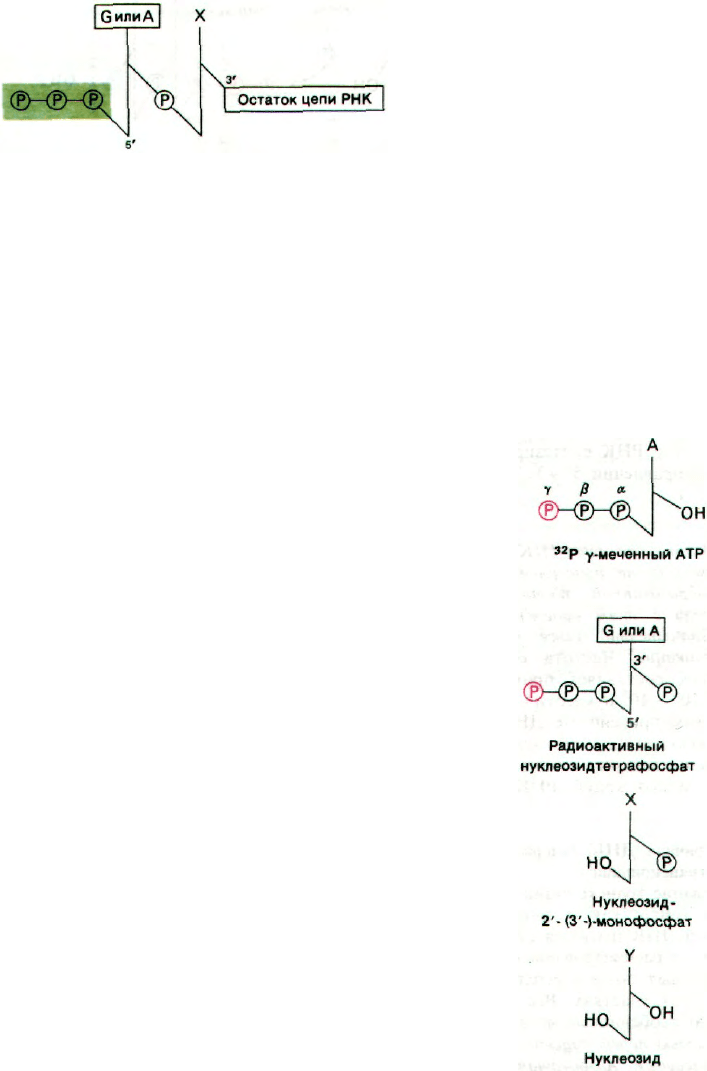
Этот ярлычок на 5'-конце был открыт
двумя способами. Было обнаружено, что
цепи РНК включают
32
Р, если инкуба-
ционная смесь содержит меченный по
γ-атому
32
Р-АТР. Очевидно, что включен-
ная метка должна быть локализована на
конце, так как только α-атом фосфора ну-
клеозидтрифосфата может участвовать
в образовании внутренних фосфодиэ-
фирных мостиков РНК. Кроме того, при
щелочном гидролизе новосинтезированной
РНК образуются продукты трех типов: ну-
клеозиды, нуклеозид-2'-монофосфаты (или
нуклеозид-3'-монофосфаты) и нуклеозид-
тетрафосфаты (рис. 25.15). Если в ка-
честве субстрата использовали γ-
32
Р-АТР,
образовывался аденозин-3'-фосфат-5'-три-
фосфат. γ-Атом фосфора этого нуклео-
зидтетрафосфата содержал метку. Анало-
гичный результат был получен с γ-
32
P-GTP.
Однако, если взять γ-меченный СТР или
UTP, радиоактивность не включается.
Следовательно, новообразованная цепь
РНК имеет трифосфатную группу на
5'-конце и свободную ОН-группу на 3'-конце.
25.14. Цепи РНК синтезируются
в направлении 5'—>3'
В каком направлении синтезируется РНК?
В направлении 5'—>3' или 3'—>5'? Два
противоположных механизма роста цепи
проиллюстрированы на рис. 25.16. При ро-
сте в направлении 5'—>3' трифосфатный
конец образуется в начале роста цепи,
а при росте 3'—>5' трифосфатный конец
появляется с последним включенным
остатком. Кинетика включения радиоак-
тивности в РНК, синтезированную из
γ-
32
P-GTP (или АТР), указывает, какая из
этих возможностей имеет место. Если в ка-
честве субстрата использовать γ-
32
P-GTP,
то отношение включения
32
Р к общему
включению нуклеотидов в продукт дости-
гает максимума очень быстро после сме-
шивания компонентов, а затем постепенно
снижается во времени. При этом общая ра-
диоактивность уже помеченной РНК не
снижается при последующем добавлении
большого избытка нерадиоактивного GTP
в инкубационную смесь. Таким образом,
32
Р включается в молекулу РНК в начале
синтеза, а не в конце. Следовательно, рост
цепи РНК идет в направлении 5'—>3', как
при синтезе ДНК.
Кор РНК-полимеразы движется вдоль
цепи матричной ДНК в направлении 3'—>
—>5', так как матричная цепь антипарал-
лельна новосинтезированной цепи ДНК.
Одна и та же молекула РНК-полимеразы
синтезирует весь транскрипт - иными сло-
вами, транскрипция процессивна. По мере
того как расплетается очередной участок
ДНК, транскрибированный участок восста-
навливает свою двухспиральную конфор-
мацию. Максимальная скорость элонгации
составляет примерно 50 нуклеотидов
в секунду.
Необходимо подчеркнуть, что РНК-по-
лимераза не обладает нуклеазной актив-
Рис. 25.15. Продукты щелочного гидроли-
за цепи РНК, меченной с по-
мощью γ-
32
Р-АТР.
25. Информационная РНК
и транскрипция
57

Рис. 25.16. Предполагаемое положение
32
Р-метки в случае роста в на-
правлении 5'—>3' и 3'—>5'. На-
блюдаемое в действительности
положение метки показывает,
что РНК синтезируется в на-
правлении 5'—>3'.
ностью. В отличие от ДНК-полимеразы
РНК-полимераза не проверяет правильно-
сти новообразованной полинуклеотидной
цепи. В связи с этим надежность тран-
скрипции значительно ниже, чем надеж-
ность репликации. Частота ошибок при
синтезе РНК составляет примерно одну
ошибку на 10
4
—10
5
нуклеотидов, что в 10
5
раз выше, чем при синтезе ДНК. Гораздо
более низкую надежность синтеза РНК
клетка обходит тем, что с одного гена син-
тезируется много копий РНК-транскрип-
тов.
25.15. Матричная ДНК содержит стоп-сиг-
налы для транскрипции
Терминирование транскрипции регулирует-
ся так же тонко, как и инициирование.
В матричной ДНК имеются стоп-сигналы,
которые были расшифрованы путем срав-
нения различных последовательностей ос-
нований в этих участках. Все они имеют
одну общую особенность: вслед за GC-бо-
гатой областью перед участком термина-
ции располагается АТ-богатая последова-
тельность. Отличительная особенность
терминирующих последовательностей —
симметрия второго порядка этой GC-бога-
58
Часть IV.
Информация
той области (рис. 25.17). Следовательно,
РНК-транскрипт этой области самоком-
плементарен, а это значит, что он способен
к спариванию оснований, приводящему
к образованию структуры шпильки
(рис. 25.18). Кроме того, новообразованные
цепи РНК кончаются несколькими остат-
ками U, которые кодируются серией осно-
ваний А в АТ-богатой области ДНК-ма-
трицы. Одна или несколько подобных
структурных особенностей заставляют
РНК-полимеразу задержаться, сделать
паузу, когда она сталкивается с таким сиг-
налом. В некоторых участках терминации
новообразованные цепи РНК высвобо-
ждаются без участия дополнительных бел-
ков. В других местах для терминации цепи
необходимо участие белка ρ (ро).
25.16. Белок ρ участвует в терминировании
транскрипции
Тот факт, что терминирование транскрип-
ции в некоторых участках происходит
с участием белка ρ, подтверждается сле-
дующими данными: молекулы РНК, син-
тезированные in vitro в присутствии белка
ρ, короче молекул, полученных в его отсут-
ствие. Например, РНК, синтезированная на
ДНК фага fd в присутствии белка ρ, имеет
коэффициент седиментации 10S, тогда как
РНК, синтезированная в отсутствие бел-
ка ρ,- 23S. Дополнительные сведения о дей-
ствии белка ρ были получены при добавле-
нии этого фактора терминации в инкуба-
ционную смесь через различные промежут-
ки времени после начала синтеза РНК.
Если белок ρ добавляли через несколько
секунд, через 2 и 10 мин после инициации,
получали РНК с коэффициентами седимен-
тации 13, 17 и 23S соответственно. Этот ре-
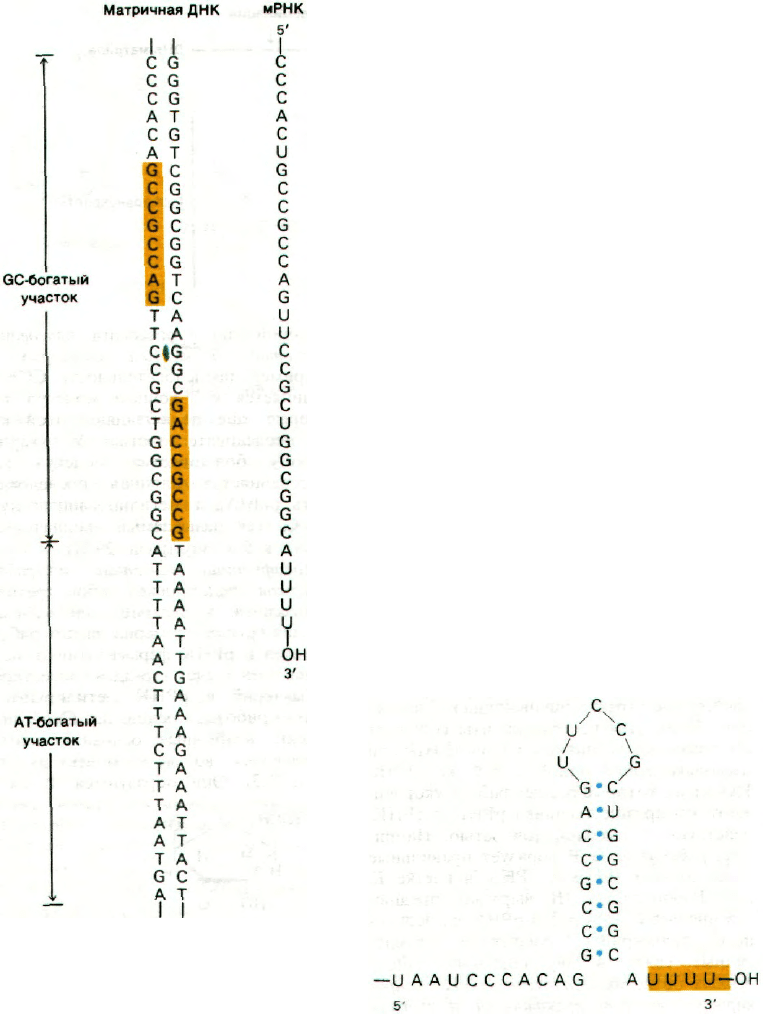
Рис. 25.17. Последовательность ДНК,
соответствующая 3'-концу
trp-мРНК E.coli. Последова-
тельности оснований, пока-
занные желтым цветом, обла-
дают симметрией второго по-
рядка относительно оси, обо-
значенной зеленым цветом.
зультат свидетельствует о том, что матри-
ца содержит но крайней мере три участка
терминации, чувствительных к белку ρ (они
дают 10S-, 13S- и 17S-PHK), и один участок
терминации, не требующий белка ρ (дает
23S-PHK). Следовательно, специфическая
терминация может происходить и в отсут-
ствие белка ρ. Однако белок ρ выявляет
дополнительные сигналы терминации, ко-
торые сама по себе РНК-полимераза узна-
вать не может.
Каким образом белок ρ вызывает терми-
нацию синтеза РНК по достижении опре-
деленных сигналов терминации? Одна из
возможностей состоит в том, что этот бе-
лок-тетрамер с массой 200 кДа - садится
на РНК и движется по направлению к мо-
лекуле РНК-полимеразы, останавливая ее
на участке терминации. Согласно этой мо-
дели, белок ρ вытесняет РНК-полимеразу
с 3'-конца РНК, что приводит к высвобож-
дению РНК-транскрипта. Интересно отме-
тить, что при терминировании транскрип-
ции белок ρ гидролизует АТР.
Как можно было предвидеть, термини-
рование транскрипции некоторых генов
контролируется. В ходе дальнейшего изло-
жения мы увидим, что бактериофаг λ син-
тезирует белки-антитерминаторы, обеспе-
чивающие возможность транскрипции
и экспрессии некоторых генов (разд. 28.11),
У Е. coli действует система регуляции с ис-
пользованием специализированных терми-
нирующих сигналов, называемых атте-
нюаторами, для обеспечения пищевых по-
требностей клетки (разд. 28.9).
Рис. 25.18. Последовательность основа-
ний 3'-конца мРНК, транскри-
бированной с триптофанового
оперона E.coli. Возможно обра-
зование стабильной структуры
шпильки.
25. Информационная РНК
и транскрипция
59
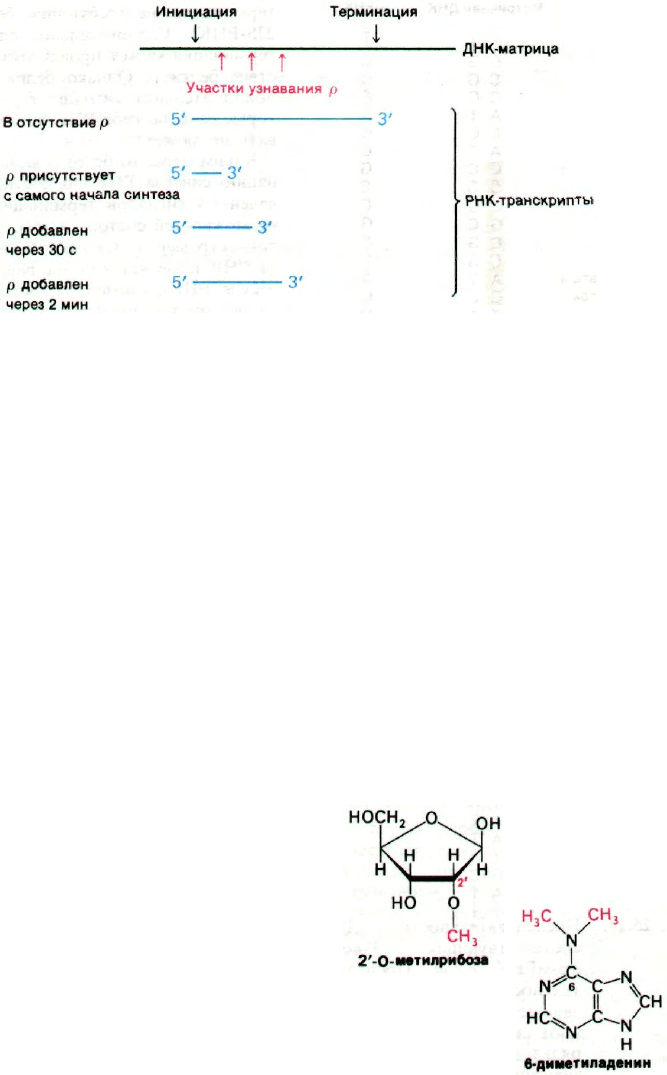
Рис. 25.19. Влияние фактора ρ на размер
транскрибируемых РНК.
25.17. Многие молекулы РНК
после транскрипции расщепляются
и химически модифицируются
Образование функционально активных мо-
лекул РНК (процессинг) продолжается пос-
ле завершения транскрипции. У прокариот
молекулы транспортных и рибосомных
РНК образуются путем расщепления и хи-
мической модификации определенных ново-
синтезированных цепей РНК. Например,
у E.coli три вида молекул рибосомных
РНК и одна молекула транспортной РНК
вырезаются из первичного РНК-тран-
скрипта, который, кроме того, содержит
спейсерные (разграничивающие) участки
(рис. 25.20). Другие транскрипты содержат
по несколько различных видов тРНК или
несколько копий одной и той же тРНК.
Нуклеазы, которые расщепляют и укорачи-
вают эти предшественники рРНК и тPHK,
действуют с высокой точностью. Напри-
мер, рибонуклеаза Р образует правильные
5'-концы всех молекул тРНК в клетке Е.
coli. Рибонуклеаза III вырезает предше-
ственники 5S-, 16S- и 23S-pPHK из первич-
ного транскрипта, расщепляя опреде-
ленные связи в двухспиральных шпи-
лечных областях. Молекулы мРНК у про-
кариот, наоборот, практически не претер-
певают модификации. Более того, многие
из них транслируются еще до того, как за-
канчивается их транскрипция. В то же вре-
мя некоторые вирусные мРНК (например,
мРНК фага Т7) расщепляются рибонуклеа-
зой III раньше, чем начинается трансляция.
60
Часть IV.
Информация
Второй тип процессинга - присоединение
нуклеотидов к концам некоторых РНК.
Например, последовательность ССА при-
соединяется к 3'-концам молекул тРНК.
которые еще не обладают этой конце-
вой последовательностью. У эукариот к
3'-концу большинства молекул мРНК
присоединяется длинная последователь-
ность poly(A), а метилированный нуклео-
тид G (так называемый «колпачок», или
«кеп») - к 5'-концу (разд. 29.22).
Модификации оснований и рибозных
остатков представляют собой третий тип
процессинга. У эукариот одна 2'-гидрок-
сильная группа примерно на сто рибозных
остатков в рРНК ферментативно метили-
руется за счет S-аденозилметионина.
У бактерий в рРНК метилируются не
остатки рибозы, а основания. Особенно ин-
тересны необычные основания, которые
встречаются во всех молекулах тРНК
(разд. 27.3). Они образуются путем фер-
ментативной модификации обычных рибо-
нуклеотидов, входящих в состав предше-
ственника тРНК. Например, псевдоуриди-
лат и риботимидилат образуются путем
модификации уридиловых остатков после
транскрипции.