Орлов Д.С. (отв. ред.) Гуминовые вещества в биосфере
Подождите немного. Документ загружается.

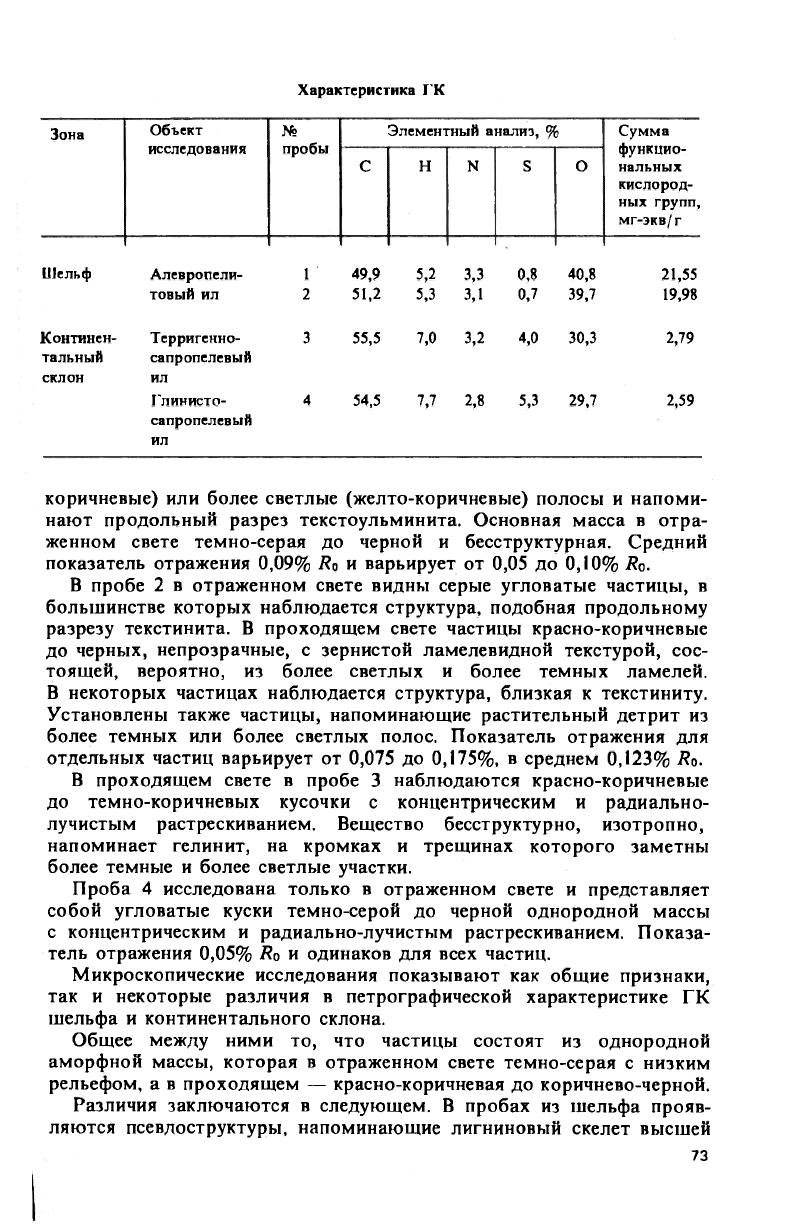
Характеристика Т'К
Зона
Объект
исследования
№
пробы
Элементный анализ, % Сумма
функцио-
нальных
кислород-
ных групп,
мг-экв/г
Зона
Объект
исследования
№
пробы
С
Н N S О
Сумма
функцио-
нальных
кислород-
ных групп,
мг-экв/г
Шельф
Алевропели- 1 49,9 5,2 3,3 0,8 40,8
21,55
товый ил 2 51,2
5,3
3,1
0,7 39,7
19,98
Континен- Терригенно-
3 55,5
7,0
3,2 4,0 30,3 2,79
тальный сапропелевый
склон ил
Глинисто- 4
54,5 7,7 2,8 5,3
29,7
2,59
сапропелевый
ил
коричневые) или более светлые (желто-коричневые) полосы и напоми-
нают продольный разрез текстоульминита. Основная масса в отра-
женном свете темно-серая до черной и бесструктурная. Средний
показатель отражения 0,09% /?о и варьирует от 0,05 до 0,10% R
0
.
В пробе 2 в отраженном свете видны серые угловатые частицы, в
большинстве которых наблюдается структура, подобная продольному
разрезу текстинита. В проходящем свете частицы красно-коричневые
до черных, непрозрачные, с зернистой ламелевидной текстурой, сос-
тоящей, вероятно, из более светлых и более темных ламелей.
В некоторых частицах наблюдается структура, близкая к текстиниту.
Установлены также частицы, напоминающие растительный детрит из
более темных или более светлых полос. Показатель отражения для
отдельных частиц варьирует от 0,075 до 0,175%, в среднем 0,123% Ro.
В проходящем свете в пробе 3 наблюдаются красно-коричневые
до темно-коричневых кусочки с концентрическим и радиально-
лучистым растрескиванием. Вещество бесструктурно, изотропно,
напоминает гелинит, на кромках и трещинах которого заметны
более темные и более светлые участки.
Проба 4 исследована только в отраженном свете и представляет
собой угловатые куски темно-серой до черной однородной массы
с концентрическим и радиально-лучистым растрескиванием. Показа-
тель отражения 0,05% Ro и одинаков для всех частиц.
Микроскопические исследования показывают как общие признаки,
так и некоторые различия в петрографической характеристике ГК
шельфа и континентального склона.
Общее между ними то, что частицы состоят из однородной
аморфной массы, которая в отраженном свете темно-серая с низким
рельефом, а в проходящем — красно-коричневая до коричнево-черной.
Различия заключаются в следующем. В пробах из шельфа прояв-
ляются псевдоструктуры, напоминающие лигниновый скелет высшей
73

растительности и соответствующие угольным материалам (текстинит,
тесктоульминит) (Штах и др., 1978). Эти ГК отличаются зернистым
или ламелевидным строением и характерными трещинами. Средний
показатель отражения выше — 0,09% Ro для пробы 1 и 0,12%
для пробы 2. Эти ГК, вероятно, продуцированы из детритных,
коллоидально-водорастворимых форм ОВ, привнесенных с континента.
ГК континентального склона представлены бесструктурным изо-
тропным веществом, напоминающим гелинит с концентрическим и
радиально-лучистым растрескиванием. Показатель отражения для
пробы 4 совсем невысок — 0,05% Ro. На основании литологической
характеристики осадков склона, соответственно терригенно- и гли-
нисто-сапропелевых илов, мы можем допустить, что основной про-
дуцент для этих ГК — останки зоо- и фитопланктона.
ЛИТЕРАТУРА
Велев В.. Маркова К. Исследования гуминовых кислот, выделенных из шельфовых,
склоновых и абиссальных осадков в западной части Черноморской впадины // Петрол-
геохим 88,Сцолнок, ВНР. Секция А. Геохимия: Тезисы. Сцолнок, 1988. С. 137—145.
Маркова К., Петрова Р. Хуминови киселини в съвременните морски утайки
от Българското черноморско крайбрежие // Год. СУ. ГГФ. 1985. Т. 75, кн. I. С. 77—98.
Штах Э., Маковски М., Тейхмюллер М. и др. Петрология углей. M.: Мир,
1978. 554 с.
Kononova M M. Soil organic matter. Oxford: Pergamon press, 1966. 544 p.
Rashid M. Geochemistry of marine humic compounds. N.Y. etc.: Springer, 1985. 300 p.
УДК 615.32:547.992.2/015.11
З.Ф. Косьянова, Д.С. Орлов, Я.М. Аммосова
ГУМИНОВЫЕ КИСЛОТЫ ПЕЛОИДОВ
Актуальность изучения ГК пелоидов вызывается необходимостью
исследования их строения и свойств, механизма действия на организм
в целях расширения предпосылок к получению новых лекарственных
препаратов.
Известно, что ГК почв и торфов обладают высокой биологической
активностью: оказывают стимулирующее действие на брожение дрож-
жей, активность ферментов, прорастание семян; снимают поражения
у растений, вызванные пестицидами; повышают адаптацию к герби-
цидам; способствуют заживлению кожных дефектов; обладают бакте-
рицидным действием; оказывают профилактическое действие при
болезнях, вызванных авитаминозами и нарушением обмена веществ,
и т.д. Выявлена полная безвредность ГК в опытах по определению
"острой" и "хронической" токсичности (Соловьева и др., 1982).
Объектами исследования были выбраны пелоиды трех типов:
© З.Ф. Косьянова, Д.С. Орлов, Я.М. Аммосова, 1993
74

1) тамбуканская иловая высокоминерализованная сильносульфидная
лечебная грязь; 2) чокракская иловая, насыщенная солями, сильно-
сульфидная лечебная грязь; 3) высокоминерализованная слабосуль-
фидная лечебная грязь вулкана Бабазанан (сопочная).
ГК были выделены по методике Д.С. Орлова г соавт. ( Орлов,
Гришина, 1981) из средних проб лечебных грязей, а для тамбу-
канской — также из сезонных и послойных образцов лечебных
слоев (черный и темно-серый ил). Очистку ГК проводили путем
неоднократного переосаждения, центрифугирования и электродиализа.
Впервые для изучения строения и свойств ГК пелоидов были
применены современные методы исследования: электронные и моле-
кулярные спектры поглощения, рентгендифрактограммы, термограм-
мы, ЭПР-спектры, электронная растровая микроскопия, молекулярно-
массовое распределение и др.
В результате изучения элементного состава ГК установлена отрица-
тельная степень окисленности ГК пелоидов; иными словами, ГК
пелоидов имеют восстановленный характер и в макромолекулах ГК
атомы водорода преобладают над атомами кислорода.
На диаграмме атомных отношений Н:С — 0:С по Ван-Кревелену
ГК пелоидов расположены в одной области с ГК сапропелей и
контактируют с областями расположения ГК и гиматомелановых
кислот (ГМК) почв. Такое расположение ГК пелоидов на диаграмме
атомных отношений Н:С — 0:С обусловлено более высокой мети-
лированностью и меньшей окисленностью ГК пелоидов в сравнении
с почвенными.
Для осенних образцов ГК тамбуканской грязи наблюдалось
повышение содержания углерода и кислорода, а водорода, наоборот,
понижение.
Методом гель-хроматографии показана высокомолекулярная и
полидисперсная природа ГК. Часть макромолекул ГК пелоидов
имеет молекулярные массы (ММ) свыше 150000, но меньше
200 000. Результаты изучения представлены в табл. 1.
Химическими методами показано присутствие кислотных функцио-
нальных групп в ГК пелоидов, т.е. карбоксильных групп и феноль-
ных гидроксилов, водород которых способен к реакциям замещения.
Для сравнительной характеристики ГК пелоидов определены также
пороги агрегации (табл. 2).
Исходя из полученных результатов определения порогов агрегации,
можно предположить большую гидрофильность ГК тамбуканской
и чокракской лечебных грязей вследствие преобладания в их молекулах
боковых цепей, несущих гидрофильные группы, чем таковую ГК
сопочной грязи, в молекулах которых должны преобладать сетки
ароматического углерода.
Для изучения строения ГК пелоидов применен метод кислотного
гидролиза (табл. 3). После мягкого гидролиза ГК в гидролизатах
идентифицированы моносахариды (глюкоза и галактоза) и дисахариды
(сахароза, мальтоза и лактоза), а также 15 аминокислот. Не обнару-
жены валин, лейцин, изолейцин, фенилаланин, изатин, метионин,
75

Таблица 1
Молекулярные массы ГК пелоидов
Источник ГК Молекулярные массы фракций
М
I II I III
IV V VI VII
VIII
Тамбукан- >150000 26060 6000 65600
екая грязь
Чокракская >150000 54 700 38500 15400 50000
грязь
Грязь вул- >150000 52600 29300 15400 9300 5100 3100 1600 39600
кана Баба-
занан
Таблица 2
Функциональные группы и порот агрегации ГК пелоидов
ИСТОЧНИК ГК
Функциональные группы, мг-экв/г
соон+он соон он
ЕМКОСТЬ об
мена, мг-экв/
100 г ГК
Порог агрега-
ции, мг-экв
СаОЬ/л гума-
та натрия
Тамбуканская
грязь
Чокракская
грязь
Г
рязь вулкана
Бабазанан
7,10 2,70
4,40
270,0
4
5,70
2,08 3,67
203,0
4
3,40
2,24
1,16 224,0 3
T аблица 3
Характеристика гидролизатов ГК пелоидов
Источник ГК
Выход ГВ*, % Азот аминный, %
от N
0
g
m
Углеводы гидроли-
зата, %
2%-ная
НС1
6 н. НС1
2%-ная
НС1
6 н. НС1
2%-ная
НС1
6 н. НС1
Тамбуканская 19,45 23,75 18,20 39,63
1,09
6,30
грязь
Чокракская 18,40 23,66 21,08
52,41 2,02
5,44
грязь
Грязь вулкана
Бабазанан
9,40 17,67
29,58
30,21 1,59
11,02
* Г'В — гидролизуемые вещества.
76

Таблица 4
Выход оксалатоа и бетолполикярбоновых кислот (БПК) при окислении
Источник ГК Выход, % Источник ГК
оксалатов
БПК
Тамбуканская грязь 36,10
14,20
Чокракская грязь
60,66
22,68
Грязь вулкана Бабазанан
68,78
22,71
триптофан (триптофан мог разложиться в кислой среде). В гидро-
лизатах ГК пелоидов 6 н. HCI идентифицированы те же моносахариды
и дисахариды, что и при гидролизе 2%-ной НС1, а также ксилоза.
Помимо Сахаров, в этих гидролизатах ГК идентифицировано 20 ами-
нокислот. Не обнаружены изатин и триптофан, который мог разло-
житься в кислой среде.
ГК сопочной грязи как по общему содержанию веществ в гидро-
лизате, так и по выходу аминного азота отличаются меньшей
степенью гидролиза по сравнению с ГК иловых минеральных суль-
фидных лечебных грязей. В остатках после гидролиза ГК пелоидов
растет содержание углерода, уменьшается содержание водорода,
азота и кислорода, а также снижаются атомные отношения Н:С,
что отражает рост ароматичности негидролизуемых остатков в резуль-
тате отрыва легкогидролизуемых частей макромолекул.
Для изучения строения и свойств ГК пелоидов был применен
также метод щелочного окисления перманганатом калия (табл. 4).
Более высокий выход оксалатов для ГК сопочной грязи в какой-то
мере согласуется с наибольшим выходом углеводов при мягком
гидролизе ГК сопочной грязи. Известно, что вещества, не имеющие
бензоидной структуры, дают при окислении в подобных условиях
преимущественно уксусную и щавелевую кислоты, а также воду,
что было показано, например, при окислении целлюлозы.
По степени ароматичности (сложности структуры) ГК пелоидов
можно расположить в следующий ряд: ГК тамбуканской грязи < ГК
чокракской грязи < ГК вулкана Бабазанан.
Электронные спектры поглощения ГК показали аналогичный ха-
рактер для всех ГК пелоидов: поглощение света убывает по мере
увеличения длины волны. Отличительной особенностью ГК тамбу-
канской грязи является присутствие в их спектрах в видимой
области максимума при длине волны около 660 нм, обусловленного
фракцией P
t
.
Получены коэффициенты экстинкции 4651 ом
,для
тамбуканской
грязи — 0,03—0,05, ГК чокракской грязи — 0,03, ГК вулкана
Бабазанан — 0,04.
Анализ ИК-спектров исходных ГК и остатков после гидролиза
свидетельствует о значительном вкладе гидролизуемых структур в
построение макромолекул ГК пелоидов. При гидролизе отщепляется
77

значиительная часть структурных единиц по эфирным и пептидным
связям, что ведет к повышению интенсивности полос, характерных
для СООН-групп, причем при более жестком гидролизе происходит
более полное отщепление. Исчезновение полосы 1650 см ' после
жесткого гидролиза также свидетельствует об отщеплении перифери-
ческой алифатической части макромолекул ГК.
На рентгендифрактограммах ГК пелоидов присутствуютмаксимумы-
гало с вершинами 4,2 А (для ГК тамбуканской грязи), 4,2—4,4 А
(для ГК чокракской грязи) и 4,4 А (для ГК вулкана Бабазанан),
а также максимумы с вершинами 3,4 А, характерные для упорядочение
расположенных ароматических структур.
Характер рентгендифрактограмм ГК пелоидов указывает на отсут-
ствие кристаллического строения их макромолекул, на сходство в
строении ГК различных пелоидов, на присутствие в их структуре
как ароматических, так и алифатических компонентов, а также на
относительно большую упорядоченность периферической части в
макромолекулах ГК вулкана Бабазанан. Полученные результаты
подтверждаются результатами элементного анализа, гель-хромато-
графии, кислотного гидролиза, определения степени ароматичности
ГК пелоидов.
Электронная микроскопия ГК пелоидов свидетельствует о слоис-
том, текстурированном сложении обломков ГК. Для ГК тамбуканской
грязи характерно проявление неравномерности стяжения ассоциатов
макромолекул при высыхании (волнистая ребристость). Для всех
ГК проявился тип упаковки ассоциатов, названный нами "чешуй-
чатостью".
Анализ термограмм ГК пелоидов свидетельствует о наибольшем
содержании термонестабильных группировок в составе ГК тамбу-
канской грязи и о наименьшем — в составе ГК вулкана Бабазанан.
В спектрах ЭПР ГК регистрируются три типа парамагнитных
центров с ^-факторами 4,3; 2,1 и 2,003. Можно считать, что сигнал
с ^-фактором 2,003 является сигналом органических свободных
радикалов (CP), а сигналы с g-факторами 4,3 и 2,1 обусловлены
органо-минеральными комплексами.
Таким образом, установлено сходство строения ГК разных типов
пелоидов. Показано, что они являются высокомолекулярными
( М — 40—66 тыс.), полидисперсными (3—8 фр.) соединениями,
содержат от 18 до 24% гидролизуемых (6 н. НС1) веществ, близки по
термической стабильности и характеру рентгендифрактограмм, элек-
тронных и ИК-спектров, по содержанию функциональных групп
(СО — 3,4—7,1 мг-экв/г, СООН — 2,2—2,7 мг-экв/г, ОН —
1,2—4,4 мг-экв/г), емкости обмена ГК (224—270 мг-экв/100 г),
величинам порогов агрегации (3—4 мг-экв СаС1
2
на. 1 л гума-
та натрия), углеводному и аминокислотному составу гидролиза-
тов; обладают парамагнитными свойствами (концентрация ПМЦ
3—6- 10
17
спин/г).
Молекулярная структура ГК пелоидов характеризуется присутствием
ароматических и алифатических компонентов. Высокие атомные отно-
шения Н:С, низкие коэффициенты экстинкции, слабовыраженные
78

полосы С=С-связей ароматических колец в ИК-спектрах, характер
реитгендифрактограмм и термограмм, содержание функциональных
групп, результаты кислотного гидролиза и щелочного окисления
позволяют считать, что в структуре ГК пелоидов, особенно в ГК
тамбуканской лечебной грязи, преобладают алифатические цепочечные
фрагменты, несущие углевод-белковые комплексы, а ароматические
выражены слабо.
ГК сопочной лечебной грязи отличаются от ГК иловых мине-
ральных сульфидных грязей меньшим содержанием азота и боль-
шим содержанием углерода. Для ГК сопочной грязи характерна
большая устойчивость к кислотному гидролизу как по общему
содержанию веществ в гидролизате, так и по содержанию амин-
ного азота, что может быть связано с более высокой степенью
гумификации ОВ сопочной грязи, с большей степенью ароматичности
и меньшим содержанием алифатических фрагментов в структуре ГК,
с наличием в молекулах ГК преимущественно циклических форм
азота, что подтверждается результатами рентгендифрактометрии,
термографии, элементного состава, порогов агрегации, определения
степени ароматичности.
Таким образом, физико-химическое изучение молекулярного стро-
ения ГК различных типов пелоидов позволило предположить наличие
биологической активности ГК. Для доказательства этого предположе-
ния проведено изучение биологической активности ГК табуканской
грязи.
В результате изучения биологической активности ГК тамбуканской
лечебной грязи установлена их высокая противовоспалительная актив-
ность, они оказывают действие на обе фазы воспалительного процесса:
снижают процессы эксудациии на 44% в дозе 10 мг/кг и на 42%
в дозе 20 мг/кг, снижают процессы пролиферации — на 55 и 50%
соответственно.
Установлена выраженная способность ГК тамбуканской лечебной
грязи нормализовать содержание лейкоцитов и лейкоцитарную фор-
мулу у животных при экспериментальной лейкопении.
ЛИТЕРАТУРА
Соловьева В.П. и др. Перспективы использования торфа в медицине // Тр.
Междунар. симпоз. IV и II Комис. МТО. Минск, 1982. С. 229—231.
Орлов Д.С., Гришина Л.А. Практикум по химии гумуса. М.: Изд-во МГУ,
1981. 271 с.

ГУМИНОВЫЕ ВЕЩЕСТВА В БИОСФЕРЕ
УДК 631.41
В.И. Кефели
ФЕНОЛЬНЫЕ СОЕДИНЕНИЯ И ГУМУСООБРАЗОВАНИЕ
В СИСТЕМЕ РАСТЕНИЕ-ПОЧВА
Сопряжение биогеохимических циклов углерода и азота связано
с формированием в биосфере ряда полимеров, к которым следует
отнести нуклеиновые кислоты, белки и ГК. Если два первых класса
биоорганических соединений относятся к системе воспроизведения
жизни (генетический код, митозы, пролиферация), то последние
определяют жизнь почвы, той геодермы, без которой невозможно
существование биосферы (Ковда, Пачепский, 1989).
Наиболее актуальными сегодня являются проблемы: генезиса ГК,
их влияния на почвенную структуру, взаимодействия гуматов с
минеральными компонентами почвы, стимуляции ростовых и формо-
образовательных процессов растений с помощью гуматов.
Решения этих вопросов тесно связаны с проведением исследований
внутрипочвенной трансформации азота и углерода, низкомолеку-
лярных и предгумусовых компонентов, кинетики трансформации
и инактивации агрохимикатов в почвах и водах в зависимости от
их биохимической и физико-химической активности. В целом для
экосистемы важна и пока не выяснена мембранная и барьерная
роль почвы и фитомассы в процессах массопереноса биофилов и
биоцидов в системе почва—растение—вода.
Итак, образование гумуса и его компонентов связывается непре-
рывной метаболической цепью с процессами фиксации СО2 листьями
растений и с синтетической деятельностью корней.
Если путь углерода при фотосинтезе начинается в растении и
продолжается при трансформации биотических элементов в почве,
то равным образом в почвенный азот, поступая в растение, включается
в состав органических соединений и определяет функцию нуклеино-
вых кислот, белков, алкалоидов и ряда фитогормонов. При этом
скорости трансформации С и N в растении и почве часто резко
различаются.
© В.И. Кефели, 1993
80

Мы еще не знаем всей совокупности растений, особенно интенсивно
истощающих почвы, растений — обогатителей почвы и растений-
санитаров, интенсивно и избирательно очищающих почву от ток-
сических элементов и поллютантов.
Без знания генезиса ГК невозможно найти пути обогащения почвы
гумусом.
В процессе фотосинтеза в хлоропластах осуществляется биосинтез
основных полифенолов и флавоноидов (рис. 1).
Свет, как известно, неоднозначно влияет на биосинтез фенольных
соединений, активируя ряд фоторецепторов. Основные фоторецепторы
в регуляции уровня флавоноидов и фенилпропаноидов следующие:
фитохром, фоторецептор синего света, фоторецептор далекого крас-
ного света.
Под действием красного света увеличивается содержание антоцианов,
фенолкарбоновых кислот и некоторых флавонолов типа кверцетин-
производных. Вместе с тем некоторые классы фенолов не являются
фотозависимыми или, по крайней мере, не активируются системой
фитохрома, например некоторые флавоны и изофлавоны.
Следует так интерпретировать фотозависимый биосинтез феноль-
hv
, .
СО, *•
I
С|
I ,
OI седогеитулозо-фосфат *• хинная
хлор.
1
"
кислота шикимован кислота хоризмовая кислота
БИОГЕНЕЗ ФЕНОЛЬНЫХ КОМПОНЕНТОВ ГУМАТОВ
ll V
Фал-лиаза
фешшалашш
фитохром
коричная кислота н-кумарован
кислота
Рис. 1 . Шикиматный путь (от фотосинтеза к полифенолам)
hv
фенол-
карбоновые
кислоты
/1-кумарат
Дипсиды Халкопы Лигнин
(хлорогенопал Флпвононды
кислота)
Рис. 2. Путь от коричной кислоты к полифенолам
6. Гуминовые вещества в биосфере
81
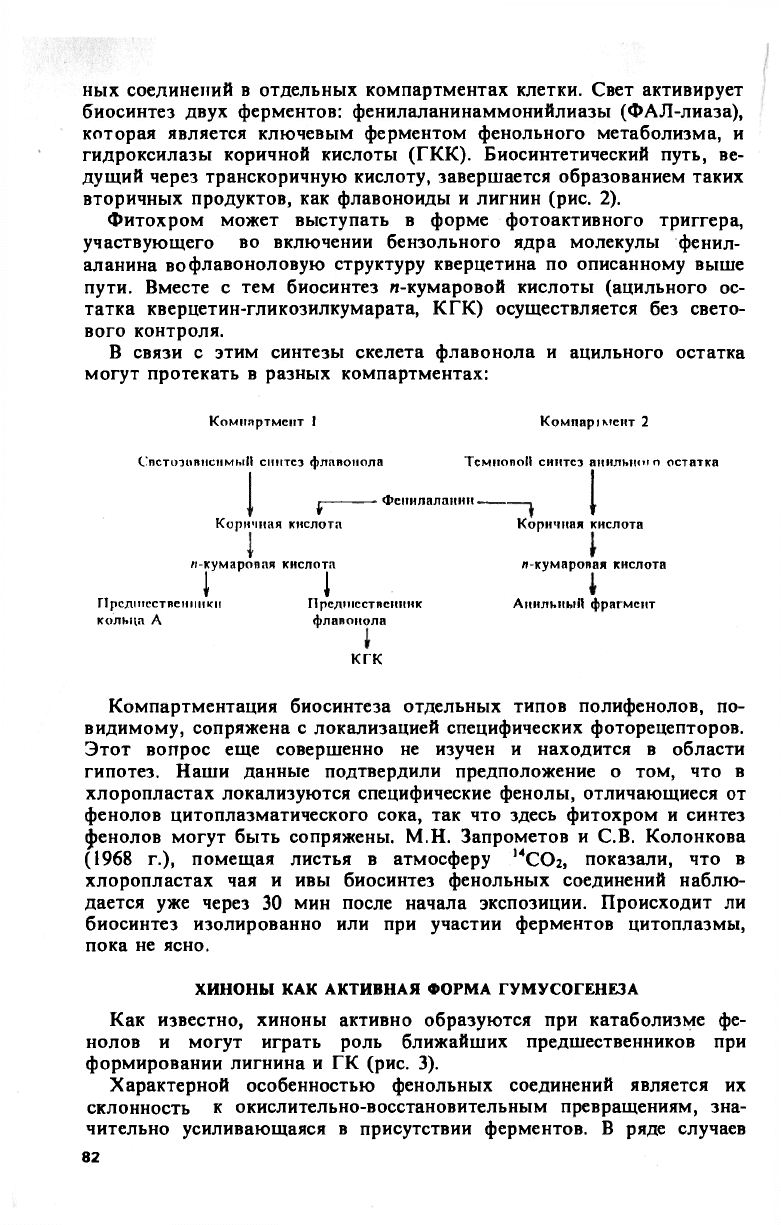
ных соединений в отдельных компартментах клетки. Свет активирует
биосинтез двух ферментов: фенилаланинаммонийлиазы (ФАЛ-лиаза),
которая является ключевым ферментом фенольного метаболизма, и
гидроксилазы коричной кислоты (ГКК). Биосинтетический путь, ве-
дущий через транскоричную кислоту, завершается образованием таких
вторичных продуктов, как флавоноиды и лигнин (рис. 2).
Фитохром может выступать в форме фотоактивного триггера,
участвующего во включении бензольного ядра молекулы фенил-
аланина вофлавоноловую структуру кверцетина по описанному выше
пути. Вместе с тем биосинтез л-кумаровой кислоты (ацильного ос-
татка кверцетин-гликозилкумарата, КГК) осуществляется без свето-
вого контроля.
В связи с этим синтезы скелета флавонола и ацильного остатка
могут протекать в разных компартментах:
Компартмепт I Комнаррмент 2
СпстоэивненмыН синтез флавонола ТсмпопоН синтез анильнто остатка
—
Фепилаланин -
i
Коричная кислота Коричная кислота
и-кумаропая кислота
I I
Предшественники Предшественник
кольца А флавонола
КГК
л-кумаровая кислота
АнильныП фрагмент
Компартментация биосинтеза отдельных типов полифенолов, по-
видимому, сопряжена с локализацией специфических фоторецепторов.
Этот вопрос еще совершенно не изучен и находится в области
гипотез. Наши данные подтвердили предположение о том, что в
хлоропластах локализуются специфические фенолы, отличающиеся от
фенолов цитоплазматического сока, так что здесь фитохром и синтез
фенолов могут быть сопряжены. М.Н. Запрометов и С.В. Колонкова
(1968 г.), помещая листья в атмосферу
14
С02, показали, что в
хлоропластах чая и ивы биосинтез фенольных соединений наблю-
дается уже через 30 мин после начала экспозиции. Происходит ли
биосинтез изолированно или при участии ферментов цитоплазмы,
пока не ясно.
ХИНОНЫ КАК АКТИВНАЯ ФОРМА ГУМУСОГЕНЕЗА
Как известно, хиноны активно образуются при катаболизме фе-
нолов и могут играть роль ближайших предшественников при
формировании лигнина и ГК (рис. 3).
Характерной особенностью фенольных соединений является их
склонность к окислительно-восстановительным превращениям, зна-
чительно усиливающаяся в присутствии ферментов. В ряде случаев
82