Наумов А.Д. Двустворчатые моллюски Белого моря. Опыт эколого-фаунистического анализа
Подождите немного. Документ загружается.


Глава 5. ДИНАМИКА ОБИЛИЯ
100
взрослых особей полностью подавляет оседание молоди, иначе и быть не может, при-
чем, чем продолжительнее цикл развития поселения, тем меньше вероятность найти
в нем ювенильные экземпляры. Следовательно, надо ожидать, что мы редко будем
встречать молодь и часто будем наблюдать аномальный демографический вектор в
плотных поселениях долго живущих видов двустворчатых моллюсков. Собственно
говоря, только такие поселения, в которых закономерно возникает подобный
демографический вектор, и следует называть плотными.
Построенная модель наглядно показывает, что возникновение циклических коле-
баний показателей обилия не обязательно связано с воздействием факторов внешней
среды. Действительно, описаны разнообразные циклические сукцессии, основанные
на биологических особенностях доминантных видов. Из них наиболее известны пи-
рогенные сукцессии чапарраля (Одум, 1975) и хвойно-таежных лесов (Максимов,
Ердаков, 1985).

101
É·‚‡ 6
çÖäéíéêõÖ ÄëèÖäíõ ÅàéñÖçéíàóÖëäàï
éíçéòÖçàâ ÑÇìëíÇéêóÄíõï åéããûëäéÇ
ë ÑêìÉàåà Ñéççõåà éêÉÄçàáåÄåà
Влияние некоторых двустворчатых моллюсков
на распределение сессильного бентоса
Бóльшая часть поверхности дна Белого моря сложена илистыми грунтами, и по-
этому данные организмы в основном представлены инфаунными формами. Жесткие
грунты, благоприятствующие развитию сессильного бентоса, относительно широко
распространены только в Онежском заливе и Северной части моря. Последняя аква-
тория, однако, изучена недостаточно подробно, поэтому влияние двустворчатых
моллюсков на других обитателей
морского дна рассматривалось на материале Онеж-
ского залива. В анализ были включены относительно массовые формы: Modiolus
modiolus, Mytilus edulis, Chlamys islandica и Elliptica elliptica.
Раковины исследованных видов двустворчатых моллюсков обрастают видами
Lithothamnion, различными Porifera, Hydrozoa (в основном Hydrallmania falcata), се-
дентарными Polychaeta (виды рода Spirorbis, а также Chitinopoma fabricii рассматри-
вались отдельно), видами рода Balanus (главным образом В. balanus), Verruca stroe-
mia, видами рода Heteranomia, Bryozoa, Hemithyris psittacea и Ascidia. Оказалось, что
встречаемость видов рода Lithothamnion, полихет (кроме серпулид) и Hemithyris psit-
tacea на створках половозрелых моллюсков на принятом уровне достоверности
(Р = 95% ) не отличается от нуля, поэтому в дальнейшем они не рассматриваются.
Несмотря на выраженную вертикальную зональность биоценозов Онежского за-
лива (Иванова, 1957), биомасса отдельных групп обрастателей оказывается мало свя-
занной с глубиной. Во всяком случае проведенный дисперсионный анализ не выявил
достоверного влияния глубины на обилие седентарной фауны. То же самое можно
сказать и о самих моллюсках, раковины которых подвергаются обрастанию.
Следует отметить, что кажущаяся противоречивость предыдущего абзаца легко
объяснима. Вертикальная зональность биоценозов в Онежском заливе строго соблю-
дается только в верхних отделах сублиторали, где она вызвана влиянием освещенно-
сти на распространение макрофитов. Что же касается зообентоса, то его батиметри-
ческое распределение в условиях Белого моря в основном определяется летним тер-
мическим режимом. При этом гомотермия рассматриваемого залива обеспечивает
для обитающих в нем видов равно благоприятные условия на любых глубинах.
Изучение материала показало, что ход изменений общих биомасс как сессильных
организмов, так и моллюсков вполне идентичен (рис. 48). Проведенный дисперсион-
ный анализ выявил, что значения биомасс исследованных групп обрастателей в дон-
ных биоценозах Онежского залива существенно зависят от присутствия в том или
ином месте моллюсков-субстратов. При этом в отдельных случаях эта связь объясня-
Глава 6. ВЗАИМООТНОШЕНИЯ ДВУСТВОРОК И ДРУГИХ ОРГАНИЗМОВ
102
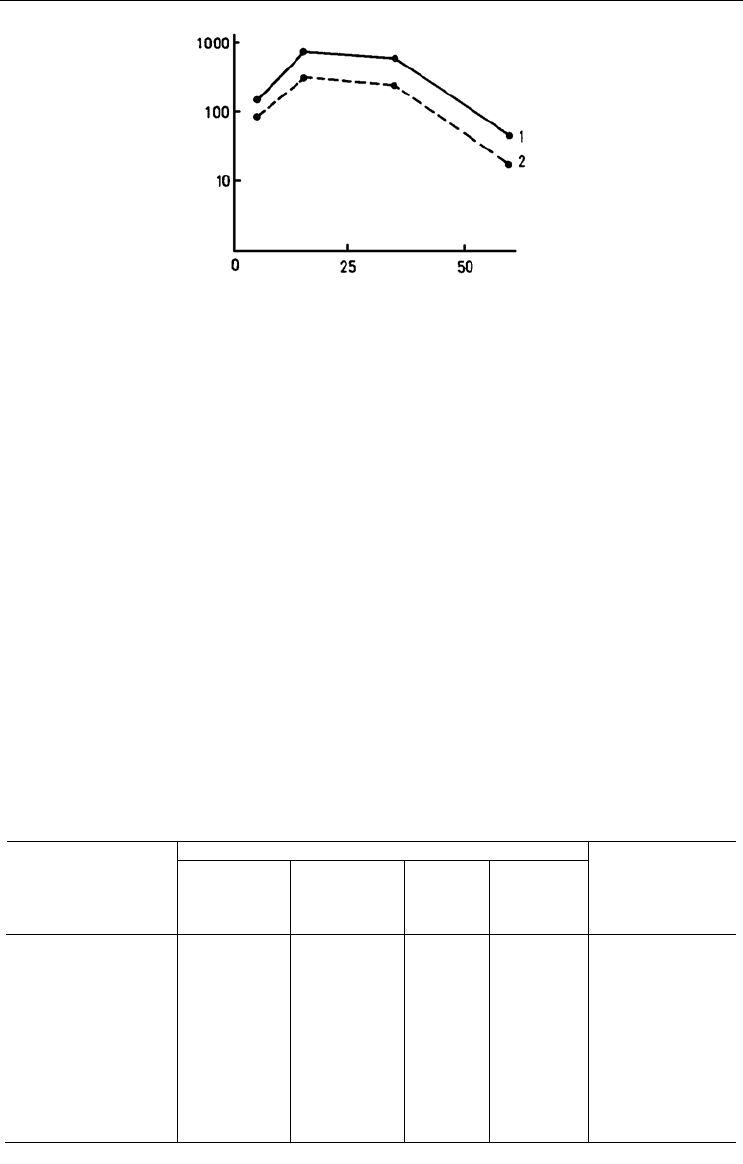
Рис. 48. Зависимость общей биомассы крупных двустворчатых моллюсков (1) и сессильных
организмов (2) от глубины.
По оси абсцисс – глубина, м; по оси ординат – биомасса, г/м
2
. По: Наумов, Федяков, 1985, а.
Fig. 48. Biomass of large bivalves (1) and sessile organisms (2) plotted against depth.
X-axis – depth (m), Y-axis – biomass (g/m2). After Naumov, Fedyakov, 1985, а.
ет значительную долю дисперсии биомассы (табл. 10). Интересно отметить, что оби-
лие той или иной группы обрастателей по большей части достоверно зависит от при-
сутствия лишь одного из видов моллюсков. Исключение составляют только виды
родов Balanus и Heteranomia. Биомасса первых заметно связана с наличием в биоце-
нозе как Modiolus modiolus, так и Мytilus edulis, а вторых – Modiolus modiolus и Еllip-
tica elliptica. В последнем случае значимым оказывается и влияние соотношения обо-
их видов.
Обращает на себя внимание то обстоятельство, что наличие или отсутствие в био-
ценозе Mytilus edulis и Elliptica elliptica не вызывает изменения биомассы подавляю-
щего большинства рассмотренных групп седентарных животных. Обилие обрастате-
лей, определяемое этими моллюсками, в то же самое время существенно зависит и от
Modiolus modiolus.
Таблица 10
Сила влияния крупных двустворчатых моллюсков на биомассу видов-обрастателей в
биоценозах Онежского залива по результатам дисперсионного анализа
По: Наумов, Федяков, 1985, а
Fraction of fouling organisms biomass dispersion (%) explained by the influence of bivalves
computed using ANOVA in bottom communities of Onega Bay.
After Naumov, Fedyakov, 1985, a.
Наличие в сообществе
Группа обрастателей
Chlamys
islandica
Modiolus
modiolus
Mytilus
edulis
Elliptica
elliptica
Соотношение в
сообществе
Modiolus modiolus
и Elliptica
elliptica
Porifera 21.0 ± 2.8 — — — —
Hydrozoa — 24.1 ± 2.6 — — —
Spirorbis spp. 17.3 ± 2.9 — — — —
Chitinopoma fabricii 25.4 ± 2.7 — — — —
Balanus spp. — 11.0 ± 3.1 9.4 ±
3.1
— —
Verruca stroemia — 39.8 ± 2.1 — — —
Heteranomia spp. — 27.8 ± 2.6 — 14.0 ± 3.1 13.1 ± 3.1
Bryozoa — 38.2 ± 2.1 — — —
Ascidia — 15.2 ± 3.0 — — —
Примечание. Знаком (—) обозначены недостоверные влияния.
ВЛИЯНИЕ ДВУСТВОРОК НА СЕССИЛЬНЫЙ БЕНТОС
103
Chlamys islandica и Modiolus modiolus в Онежском заливе часто встречаются со-
вместно. Несмотря на это, каждый из них оказывает воздействие на биомассу раз-
личных групп сессильных животных. Что же касается соотношения этих моллюсков
в биоценозе, то оно не имеет значения для обрастателей.
Биомасса обрастателей повышается в местах скопления Bivalvia (рис. 49–52). Оче-
видно, что такое повышение биомасс может быть вызвано тем, что требования к
окружающей среде седентарных животных аналогичны таковым исследованных
моллюсков и (или) тем, что обрастатели привлекаются метаболитами видов-субтра-
тов. Наконец, это обстоятельство может
объясняться увеличением пригодной для
поселения поверхности за счет крупных раковин. На нашем материале невозможно
решить, какая из названных причин превалирует; во всяком случае, последняя имеет
немаловажное значение.
В табл. 11 представлены данные об экстенсивности обрастания половозрелых эк-
земпляров изученных видов моллюсков. Видно, что, хотя все они обладают сходной
по видовому составу
фауной, имеются значительные различия в частоте встречаемо-
сти отдельных групп. Наиболее часто в составе обрастания раковин мидий встреча-
ются виды рода Balanus и различные мшанки. На значительной части створок посе-
ляются также Verruca stroemia и виды рода Heteranomia. Раковины Modiolus
modiolus, обитающего, в отличие от предыдущего вида, на больших глубинах, глав-
ным образом обрастают
Verruca stroemia и мшанками. Приблизительно половина
этих моллюсков служит субстратом для поселения Heteranomia.
Обрастания раковины Chlamys islandica, особенно ее верхней створки, очень на-
поминают то, что мы видели у Modiolus modiolus, с той лишь разницей, что здесь
снижается частота встречаемости мшанок и асцидий. Что же касается морских желу-
дей и, в особенности, полихет сем. Serpulidae,
то экстенсивность обрастания ими зна-
чительно возрастает. Встречаемость всех организмов-обитателей, за исключением
видов рода Spirorbis, на нижней створке гребешка заметно падает по сравнению с
верхней. Это в первую очередь относится к усоногим ракам и гидроидам.
Еlliptica elliptica, в отличие от всех предыдущих видов, благодаря инфаунному
образу жизни почти не подвержена
обрастанию. В сколько-нибудь заметных количе-
ствах здесь встречаются лишь различные мшанки, Verruca stroemia и, иногда, асци-
дии. В противоположность остальным рассмотренным моллюскам, где обрастанию
подвергается почти вся поверхность раковины, в этом случае сессильные формы по-
селяются лишь на ее торчащем из грунта заднем конце.
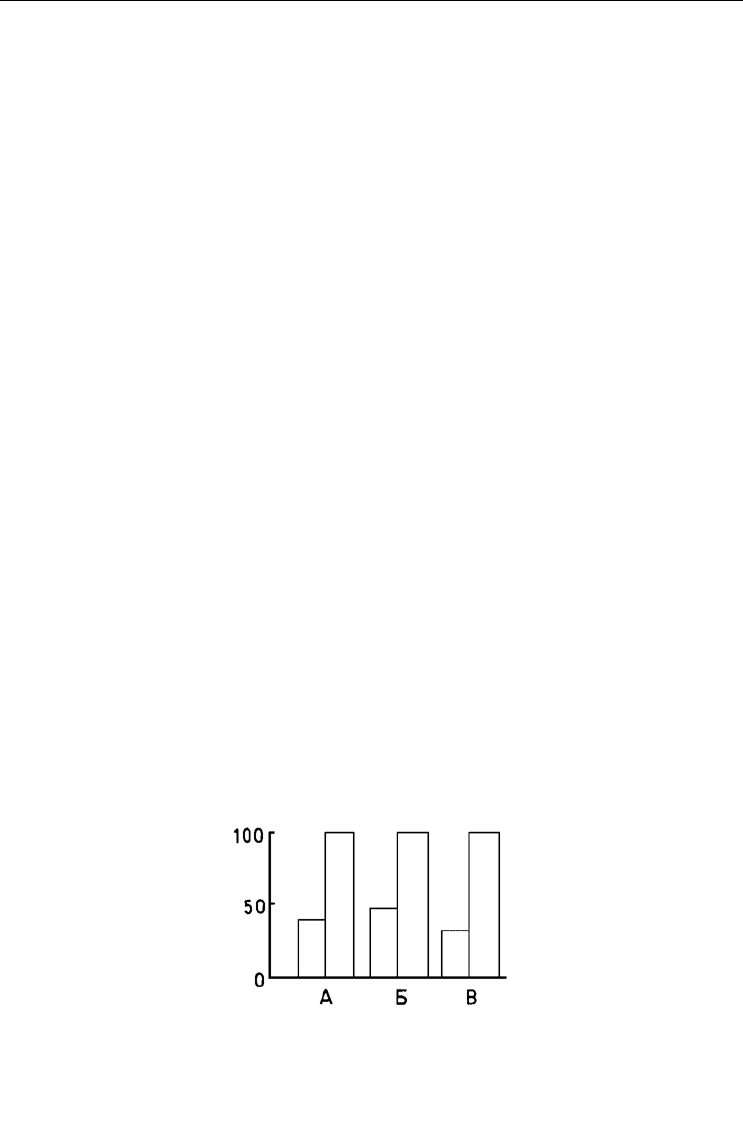
Рис. 49. Встречаемость Porifera (А), Spirorbis (Б) и Chitinopoma fabricii (В) в сообществах, где
отсутствует Chlamys islandica (левые столбцы) и где он присутствует (правые столбцы).
По оси ординат – встречаемость, %. По: Наумов, Федяков, 1985, а.
Fig. 49. Occurrence of Porifera (А), Spirorbis (Б) и Chitinopoma fabricii (В) in communities where
Chlamys islandica is absent (left bars) and where it is present (right bars).
Y-axis – occurrence (%). After Naumov, Fedyakov, 1985, а.
Глава 6. ВЗАИМООТНОШЕНИЯ ДВУСТВОРОК И ДРУГИХ ОРГАНИЗМОВ
104
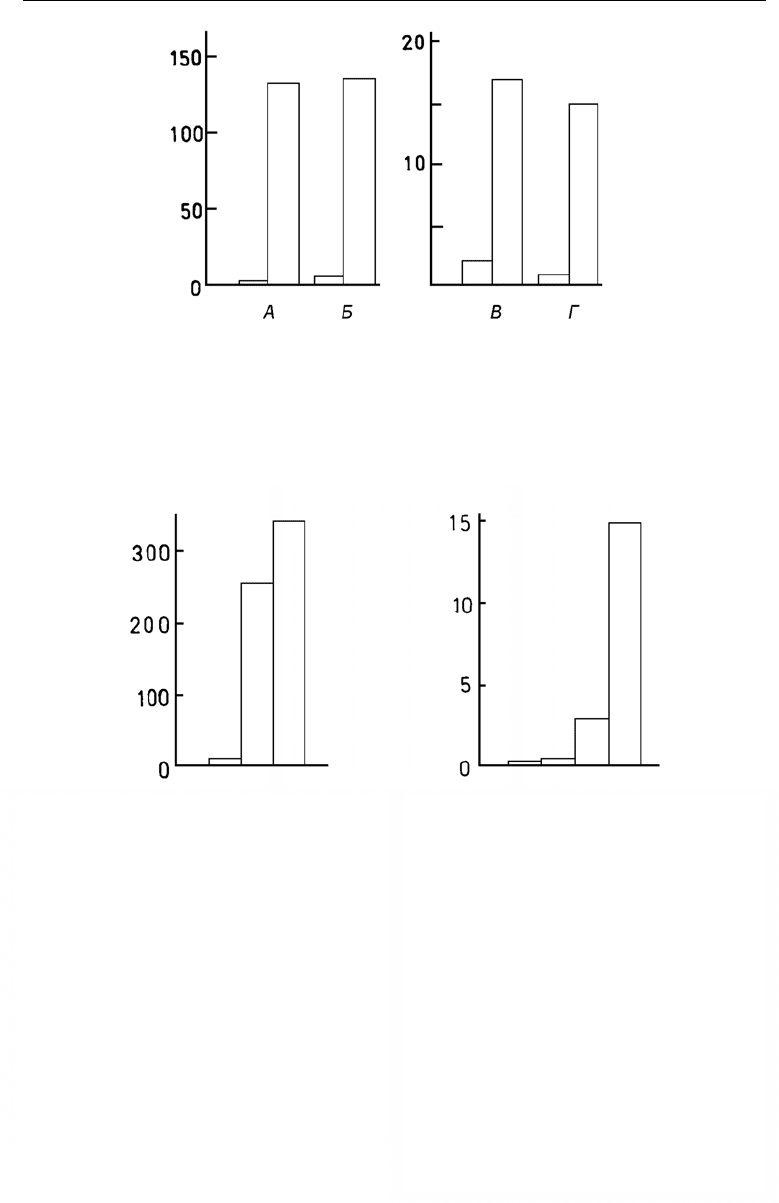
Рис. 50. Биомасса (г/м
2
, по оси ординат) Hydrozoa (А), Verruca stroemia (Б), видов рода Balanus
(В) и Ascidia (Г) в сообществах, где отсутствует Modiolus modiolus (левые столбцы) и где он
присутствует (правые столбцы).
По: Наумов, Федяков, 1985, а.
Fig. 50. Biomass (g/m
2
) of Hydrozoa (A), Verruca stroemia (Б), Balanus species (В) and Ascidia (Г)
in communities where Modiolus modiolus is absent (left bars) and where it is present (right bars).
After Naumov, Fedyakov, 1985, а.
Рис. 51. Биомасса видов рода Balanus (г/м
2
, по
оси ординат) в сообществах, где нет ни Modio-
lus modiolus, ни Mytilus edulis (левый столбец),
где присутствует Mytilus edulis (средний стол-
бец) и где встречен Mod. modiolus (правый
столбец).
По: Наумов, Федяков, 1985, а.
Fig. 51. Balanus species biomass (g/m
2
, X-axis) in
communities where neither Modiolus modiolus nor
Mytilus edulis are present (left bar); where
M. edulis (medium bar) and Mod. modiolus (right
bar) are present.
After Naumov, Fedaykov, 1985, a.
Рис. 52. Биомасса видов рода Heteranomia
(г/м
2
, по оси ординат) в сообществах, где нет
ни Modiolus modiolus, ни Elliptica elliptica
(крайний левый столбец), где имеется
E. elliptica и нет Mod. modiolus (второй слева
столбец), где есть Mod. modiolus, но нет
E. elliptica (третий слева столбец) и где
встречены оба вида моллюсков (крайний пра-
вый столбец).
По: Наумов, Федяков, 1985, а.
Fig. 52. Heteranomia species biomass (g/m
2
, X-
axis) in communities where neither Modiolus
modiolus nor Elliptica elliptica are present (left
bar); where E. elliptica is present, but
Mod. modiolus is absent (second bar); where
Mod. modiolus is present, but E. elliptica is ab-
sent (third bar) and where the both species are
present (right bar).
After Naumov, Fedaykov, 1985, a.

ВЛИЯНИЕ ДВУСТВОРОК НА СЕССИЛЬНЫЙ БЕНТОС
105
Очевидно, что различия в обрастании разных видов обусловлены избирательной
способностью обрастателей. Предпочтение той или иной раковины в качестве суб-
страта может быть обусловлено комплексом причин. К ним следует отнести различ-
ные условия среды в местах обитания моллюсков, ориентацию в пространстве и
скульптуру поверхности их створок, выделяемые ими в воду метаболиты и др. По-
видимому, первой причиной можно объяснить бедность обрастания Elliptica elliptica,
а также предпочтение раковины мидий мелководными усоногими ракообразными.
Как уже было сказано, практически все обрастатели неохотно поселяются на нижней,
соприкасающейся с грунтом створке Chlamys islandica, и только мелкие Serpulidae
довольствуются небольшим пространством между ребрами. Остальные же обраста-
тели располагаются кольцом вдоль наружного, слегка приподнятого края. Различия в
обрастании Modiolus modiolus верхней и створки Chlamys islandica объясняются, по-
видимому, другими из перечисленных причин, поскольку оба вида в исследованном
районе повсюду встречаются совместно.
Из табл. 12 хорошо видно, что большинство групп обрастателей поселяются
предпочтительно на одном-двух видах моллюсков. Так, например, Chitinopoma fab-
ricii наиболее часто встречается на створках Chlamys islandica, а виды рода Balanus –
на Mytilus edulis. Это утверждение остается в силе и для губок, гидроидов, мшанок и
асцидий, представленных значительным числом видов с весьма различными требо-
ваниями к среде обитания.
Сопоставление данных табл. 11 и 12 показывает, что биомасса той или иной груп-
пы обрастателей, как правило, существенно зависит от наличия в биоценозе именно
предпочитаемого ею вида моллюска (см. табл. 13), однако имеются и исключения.
Например, биомасса глубоководных поселений видов рода Balanus определяется на-
личием в донных биоценозах Modiolus modiolus, в то же время на раковинах гребеш-
ков эти ракообразные поселяются почти вдвое чаще. Для объяснения этого и других
подобных фактов требуются дальнейшие исследования.
Таблица 11
Экстенсивность обрастания раковин некоторых видов
двустворчатых моллюсков, %
По: Наумов, Федяков, 1985, а
Extensiveness of some clam species fouling (%)
After Naumov, Fedyakov, 1985, a
Clamys islandica
Группа обрастателей
Mytilus
edulis
Modiolus
modiolus
верхняя
створка
нижняя
створка
Elliptica
elliptica
Lithothamnoin spp. 0–1 3 ± 2 0–1 0–1 0–1
Porifera 7 ± 2 26 ± 6 28 ± 6 13 ± 4 1 ± 1
Hydrozoa 9 ± 2 26 ± 6 32 ± 6 6 ± 3 4 ± 1
Spirorbis spp. 3 ± l 15 ± 5 34 ± 6 41 ± 6 0–1
Chitinopoma fabricii 1 ± 1 26 ± 6 64 ± 6 58 ± 6 0–1
Прочие Polychaeta 0–1 4 ± 2 5 ± 3 0–1 0–1
Balanus spp. 80 ± 3 29 ± 6 48 ± 6 4 ± 2 8 ± 1
Verruca stroemia 32 ± 3 87 ± 6 74 ± 6 32 ± 6 37 ± 2
Heteranomia spp. 28 ± 3 54 ± 6 55 ± 6 40 ± 6 1 ± 1
Bryozoa 86 ± 2 89 ± 6 58 ± 6 34 ± 6 23 ± 2
Hemithyris psittacea 0–1 2 ± 2 0–1 0–1 0–1
Ascidia 3 ± 1 28 ± 6 9 ± 4 7 ± 3 13 ± 1

Глава 6.ВЗАИМООТНОШЕНИЯ ДВУСТВОРОК И ДРУГИХ ОРГАНИЗМОВ
106
Таблица 12
Взаимоотношения между видами-субстратами и организмами обрастателями
По: Наумов, Федяков, 1985, а
Mutual relations between substrate species and fouling organisms
After Naumov, Fedyakov, 1985, a
Группа обрастателей Предпочитаемый субстрат
Моллюски, влияющие на
биомассу обрастателей
Balanus (мелководные поселения) Mytilus edulis Mytilus edulis
Balanus (глубоководные поселения) Chlamys islandica
Ascidia Modiolus modiolus
Bryozoa Modiolus modiolus, Mytilus edulis
Hydrozoa, Verruca stroemia
Modiolus modiolus
Heteramonia spp.
Modiolus modiolus,
Elliptica elliptica
Porifera
Modiolus modiolus,
Chlamys islandica
Spirorbis spp., Chinopoma fabricii Chlamys islandica
Chlamys islandica
Макрообрастания раковин некоторых видов беломорских
двустворчатых моллюсков
Несмотря на то, что изучению бентоса Белого моря посвящена обширная литера-
тура, обрастание раковин моллюсков в этом водоеме практически не исследовано.
Нам известна лишь одна работа Е. А. Нинбурга (1977), в которой рассматривается
экология комменсалов Масота balthica и Portlandia arctica. В то же время раковины
двустворчатых моллюсков представляют собой превосходный субстрат для седен-
тарных видов бентоса. Массовость и относительно легкая доступность Bivalvia дела-
ют их чрезвычайно удобным объектом для изучения динамики сообществ обраста-
ния, играющих значительную роль в практической деятельности человека.
Важно отметить, что наибольшего развития обрастания достигают на раковинах
крупных двустворок, которые могут служить (а в некоторых местах уже служат) объ-
ектами искусственного разведения и промысла. Известно, что отрицательное воздей-
ствие отдельных сопутствующих видов создает серьезные трудности при организа-
ции марикультуры (Саранчова, Кулаковский, 1982).
В предыдущем разделе показана существенная роль некоторых видов двустворча-
тых моллюсков в распространении седентарных животных. Установлены также раз-
личия в обрастании раковин отдельных видов Bivalvia. Специфика обрастания мол-
люсков, по-видимому, объясняется целым комплексом причин: скульптура поверх-
ности и ориентация створок в пространстве, экологические особенности отдельных
видов и т. п. Характерное расположение седвизионтов на раковине двустворчатых
моллюсков показано на рис. 53.
Для оценки силы влияния указанных причин достаточно провести дисперсионный
анализ встречаемости седвазионтов на разных моллюсках. Однако, как было показа-
но В. В. Ошурковым (1982, 2000), значительное воздействие на последнюю оказыва-
ет время существования субстрата. В связи с этим представляется разумным вклю-
чить в анализ длину раковины, в определенной степени отражающую ее возраст
1
.
Полученные результаты двухфакторного дисперсионного анализа представлены в
1
Значительные различия максимальных размеров отдельных видов изучаемых моллюсков
вызывали необходимость пользоваться относительной длиной раковины (в процентах от мак-
симального размера вида в данной акватории).

ОБРАСТАНИЯ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ
107
Таблица 13
Доля дисперсии числа видов обрастателей и экстенсивности обрастания раковин дву-
створчатых моллюсков разными группами животных. По: Наумов, Федяков, 1985, б
Fraction of dispersion (%) of fouling organisms species number explained by the influence of
bivalves species, shell length and their mutual impact computed using ANOVA
After Naumov, Fedyakov, 1985, б
Доля дисперсии, объясняемая влиянием
Группа обрастателей
вида моллюсков длины раковины
совместного воздей-
ствия вида и длины
раковины
Spirorbis spp. 7.9 ± 0.3 8.0 ± 0.2 9.9 ± 0.9
Chitinopoma fabricii 9.9 ± 0.3 12.3 ± 0.2 9.3 ± 0.9
Balanus spp. 12.3 ± 0.3 21.7 ± 0.2 14.0 ± 0.9
Verruca stroemia 1.7 ± 0.3 5.4 ± 0.2 25.7 ± 0.8
Heteranomia spp. 9.0 ± 0.3 18.0 ± 0.2 6.6 ± 1.0
Bryozoa 11.5 ± 0.3 27.8 ± 0.2 8.4 ± 1.0
Число видов 6. 2 ± 0.3 41.3 ± 0.2 7.6 ± 1.0
Общая экстенсивность 13.3 ± 0.2 45.1 ± 0.2 3.4 ± 1.4
табл. 13. Как видно, воздействие характера субстрата, времени его существования и
их совместное влияние во всех случаях оказались достоверными, причем ведущая
роль обычно принадлежит длине раковины. Таким образом, сведения о возрасте мол-
люска дают бóльшую информацию об обросте, нежели знание его видовой принад-
лежности. Относительно низкая роль совместного воздействия рассматриваемых
факторов говорит о том, что в большинстве случаев обрастание различных раковин
одним и тем же видом протекает сходным образом. Лишь усоногие раки, в особенно-
сти Verruca stroemia, заселяют разные субстраты неодинаково. Наименьшее воздей-
ствие рассматриваемые факторы (в сумме около 26%) оказывают на встречаемость
видов рода Spirorbis. Таким образом, его присутствие в обросте раковин определяет-
ся в основном другими причинами.
Перейдем к рассмотрению развития обрастаний на раковинах двустворчатых
моллюсков. На рис. 54 и 55 показана динамика экстенсивности заселения поверхно-
сти створок. Бросается в глаза, что в отличие от остальных видов раковины Elliptica
elliptica и по достижении максимальных размеров могут быть лишены обрастаний.
Прочие моллюски обрастают однотипно, причем верхняя створка гребешка начинает
заселяться несколько раньше нижней. Обращает на себя внимание тот факт, что в
большинстве случаев зависимость экстенсивности обрастания от длины раковины
носит хорошо выраженный S-образный характер (см. рис. 54 – 55).
Обратимся теперь к динамике отдельных групп обрастателей на разных видах ис-
следованных моллюсков. Такие виды, как Portlandia arctica и Macoma calcarea, об-
растают всего одним видом седвазионтов (Нинбург, 1977), так что развитие обраста-
ния на их створках не может быть названо сукцессией. В остальных случаях мы име-
ем дело с сукцессионным процессом. При этом, как видно из рис. 56–60, смена руко-
водящих форм
1
наблюдается только в процессе развития обрастаний на раковинах
Chlamys islandicus. Заселение створок видов протекает следующим образом. В пер-
вую очередь на раковинах поселяются усоногие раки, причем на Mytilus edulis это –
виды рода Balanus, а на Modiolus modiolus и Elliptica elliptica – Verruca stroemia.
Здесь уместно отметить, что первые обрастания на мидии появляются уже по дости-
жении моллюсками 10% от дефинитивного размера, в то время как двум другим ви-
1
Применительно к использовавшейся методике руководящими формами будут именовать-
ся наиболее часто встречающиеся.
Глава 6.ВЗАИМООТНОШЕНИЯ ДВУСТВОРОК И ДРУГИХ ОРГАНИЗМОВ
108
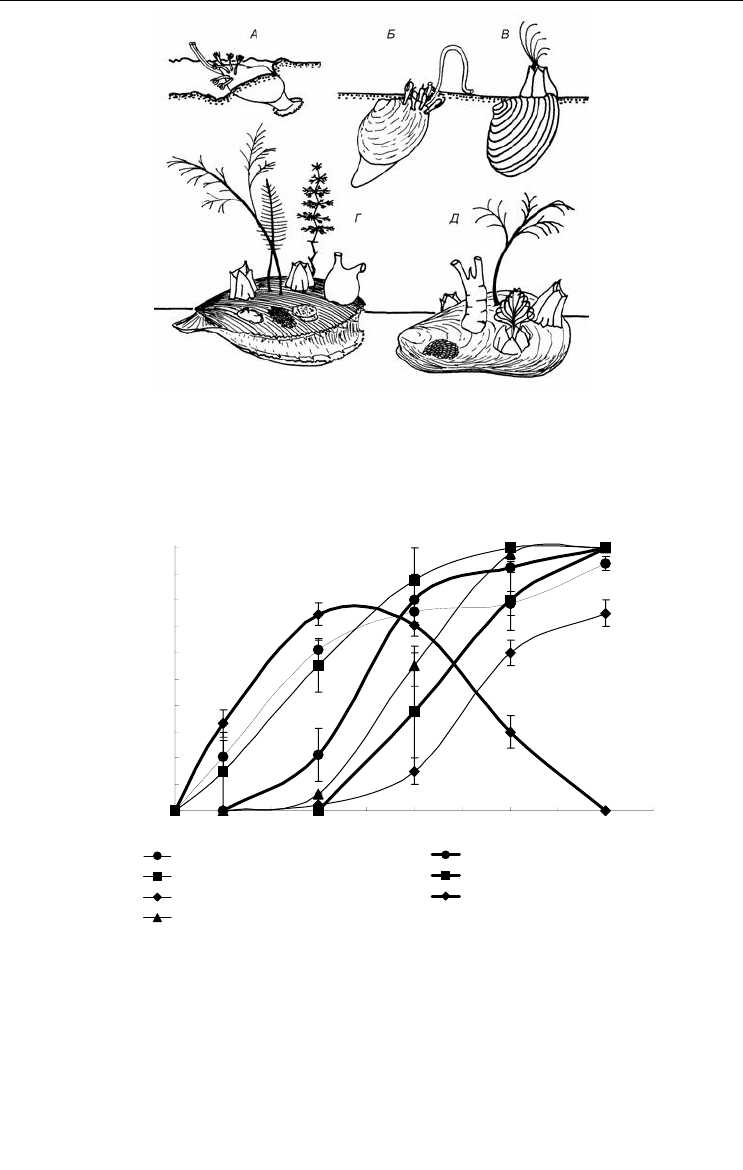
Рис. 53. Характерное размещение седвазинтов на раковинах некоторых
двустворчатых моллюсков. По: Наумов, Федяков, 1993.
А – Portlandia arctica, Б – Macoma calcarea, В – Elliptica elliptica, Г – Clamys islandica,
Д – Modiolus modiolus.
Fig. 53. Typical position of fouling organisms on shells of some bivalves.
After Naumov, Fedyakov, 1993.
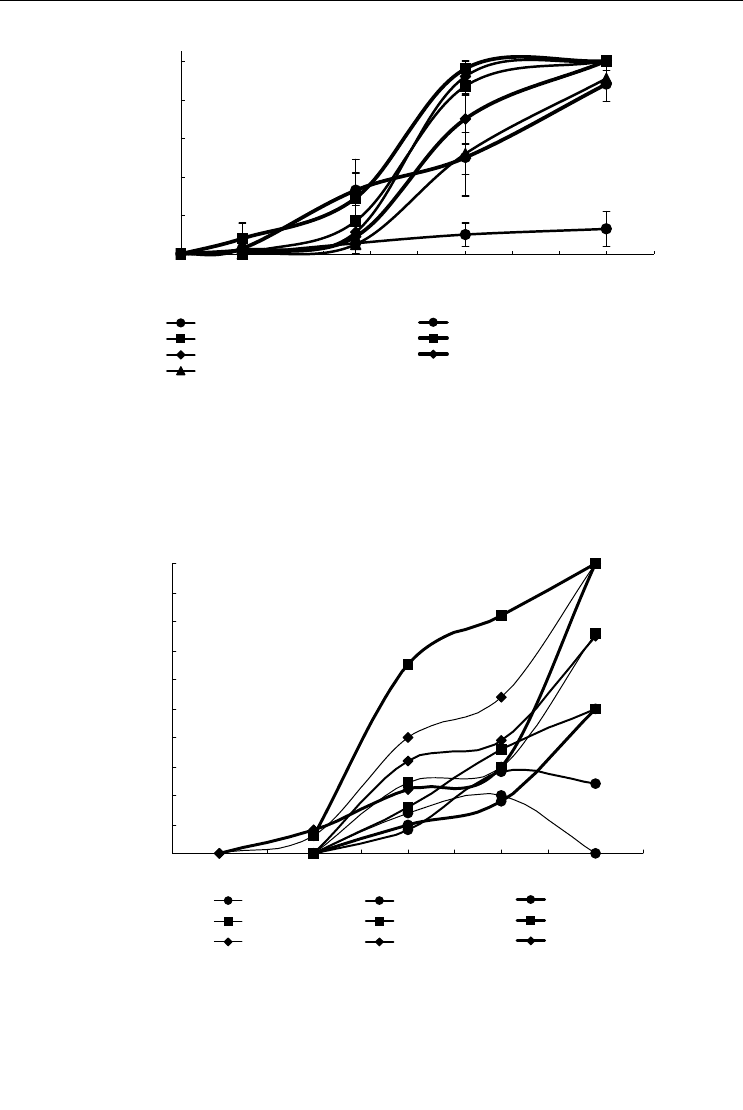
Рис. 54. Зависимость общей экстенсивности седвазии, % (по оси ординат) от относительной
длины раковины, % от максимальной длины (по оси абсцисс) на створках раковин некоторых
видов двустворчатых моллюсков в Онежском заливе.
Вертикальными линиями показана статистическая ошибка. Modiolus modiolus, Mytilus edulis, Chlamys is-
landica и Elliptica elliptica – по данным А. Д. Наумова и В. В. Федякова (1985, б), Portlandia arctica и
Macoma calcarea – по данным Е. А. Нинбурга (1977).
Fig. 54. Total extensiveness of fouling on shells in some bivalves species from Onega Bay
(%, Y-axis) plotted against the relative shell length (% of maximum length, X-axis).
Vertical bars – standard error. Modiolus modiolus, Mytilus edulis, Chlamys islandica and Elliptica elliptica ac-
cording data of Naumov and Fedyakov (1985, б), Portlandia arctica and Macoma calcarea according data of
Ninburg (1977).
0
10
20
30
40
50
60
70
80
90
100
0 102030405060708090100
Portlandia arctica Modiolus modiolus
Chlamys islandica, верхняя створка Chlamys islandica, нижняя створка
Elliptica elliptica Macoma calcarea
Mytilus edulis
ОБРАСТАНИЯ ДВУСТВОРЧАТЫХ МОЛЛЮСКОВ
109
Рис. 55. Зависимость общей экстенсивности седвазии на створках раковин Mytilus edulis в гу-
бах Чупе и Унский, а также в ряде районов Онежского залива.
Обозначения, как на рис. 54. По данным А. Д. Наумова и В. В. Федякова (1985, в) и А. Д. Наумова (1990).
Fig. 55. Total extensiveness of fouling on Mytilus edulis shells in Chupa Inlet, Ouna Inlet and some
regions of Onega Bay plotted against the shell length.
For legend see fig. 54. According data of Naumov and Fedyakov (1985, б) and Naumov (1990).
Рис. 56. Зависимость экстенсивности обрастания створок Modiolus modiolus в Онежском заливе
от длины раковины.
Обозначения по осям, как на рис. 54. По данным А. Д. Наумова и В. В. Федякова (1985, б).
Fig. 56. Extensiveness of fouling on Modiolus modiolus shells in Onega Bay plotted against the shell
length.
For legend see fig. 54. According data of Naumov and Fedyakov (1985, б).
0
20
40
60
80
100
0 102030405060708090100
Губа Чупа Восточная Соловецкая салма
Входная часть Онежского залива Центральная часть Онежского залива
Район Кондострова Кутовая часть Онежского залива
Унская губа
0
10
20
30
40
50
60
70
80
90
100
0 102030405060708090100
Porifera Hydrozoa Spirorbis spp.
Chitinopoma fabricii Balanus spp. Verruca sroemia
Heteranomia spp. Bryozoa Ascidia