Наумов А.Д. Двустворчатые моллюски Белого моря. Опыт эколого-фаунистического анализа
Подождите немного. Документ загружается.


Глава 5. ДИНАМИКА ОБИЛИЯ
90
должительность более точно не представляется возможным, так как времени
проведения наблюдений для этого явно недостаточно. Можно только отметить, что с
1987 г. (начало наблюдений) до 1990 г. биомасса мии в среднем возрастала, затем
начала падать, достигла минимума в 1999 г., а в настоящее время снова растет. Если
считать, что циклическое изменение биомассы этого
вида имеет синусоидальный
характер, то продолжительность цикла можно с некоторой осторожностью оценить
приблизительно в 18–20 лет. Сезонный тренд (рис. 39) выявляется довольно
уверенно, однако его минимумы и максимумы не имеют регулярной привязки к
определенным сезонам. Скорее всего, это говорит о том, что скорость роста молоди
зависит в значительной степени от условий конкретного года
.
Что касается динамики плотности поселения Mya arenaria в Сельдяной губе
(рис. 40), то максимумы и минимумы основного тренда этого показателя опережают
таковые биомассы приблизительно на 5 лет, что не должно вызывать удивления. В
общем можно сказать, что и в этом случае с некоторой долей вероятности
предсказывается 18 – 20-летний цикл. Помимо этого в последние годы
удается
проследить и другой, более короткий цикл, продолжающийся около 3 лет. Насколько
он надежен, покажут будущие исследования. Что же касается сезонного цикла
плотности поселения (рис. 41), то он достаточно регулярен и прослеживается на
протяжении всего времени проведения наблюдений. Его минимумы и максимумы
весьма стабильно приходятся на гидрологическую весну и гидрологическую осень
так
же, как это имеет место у Macoma balthica (см. рис. 41). Надо полагать, что и
вызывается это сходными причинами.
В Медвежьей губе основной тренд динамики биомассы не демонстрирует
заметной изменчивости (рис. 42), зато достаточно надежно выявляется цикл,
приблизительно трехлетней периодичности (рис. 43).
Плотность поселения этого вида монотонно, хотя и весьма незначительно, растет
на протяжении всего
времени наблюдений (рис. 44), и выявить какие-либо
циклические колебания этого показателя, долговременные или краткосрочные, не
удается. Зато без труда обнаруживается достаточно регулярный сезонный цикл,
сходный с таковым для поселения в Сельдяной губе (рис. 45). Таким образом, в
названных поселениях Mya arenaria преобладают несколько иные демографические
процессы, чем те, что описаны Н. В.
Максимовичем и А. В. Герасимовой для
популяций из заливов Сухая салма и Лебяжья губа (Maximovich, Guerassimova, 2003).
Динамика, сходная с той, что отмечена в губах Сельдяная и Медвежья, обнаружена в
эстуарии р. Хэмптон-Брук (штат Нью-Гэмпшир), причем там наблюдается четкая 14-
летняя периодичность в оседании молоди (Kinner, Shipman, 1985).
Модель динамики показателей обилия в
плотном поселении двустворчатых моллюсков
Хотя у нас нет фактического материала, который позволил бы однозначно
ответить на вопрос, почему в ряде случаев мы сталкиваемся с аномальным
демографическим вектором, можно построить модель развития плотного поселения
двустворчатых моллюсков, которая учитывала бы возможность снижения плотности
спата пропорционально обилию взрослых, а работу модели проверить фактическими
данными, пусть и немногочисленными.

МОДЕЛЬ ДИНАМИКИ ОБИЛИЯ
91
Рассмотрим произвольно выбранную точку в поселении двустворчатых моллю-
сков. Демографический вектор в момент времени t + 1 представляет собой произве-
дение демографического вектора в момент времени t и переходной матрицы Лесли
(Leslie, 1945, 1948; Уильямсон, 1975):
⎥
⎥
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎢
⎢
⎣
⎡
⎥
⎥
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎢
⎢
⎣
⎡
=
⎥
⎥
⎥
⎥
⎥
⎥
⎦
⎤
⎢
⎢
⎢
⎢
⎢
⎢
⎣
⎡
−
−
+
+
+
+
m,t
3,t
2,t
1,t
1m
2
1
m1m21
1m,t
13,t
12,t
11,t
n
...
n
n
n
P...
...............
...P
...P
FF...FF
n
...
n
n
n
000
000
000
, (13)
где n
i,t
– численность особей i-го возраста в момент времени t, F
i
– плодовитость сам-
ки i-го возраста, P
i
– вероятность самки i-го возраста дожить до возраста i + 1, t – те-
кущий возраст, m – предельный возраст.
Отсюда
n
i+1,t+1
= P
i,t
n
i,t
. (14)
Для массовых беломорских двустворчатых моллюсков личиночный пул (или ко-
личество молоди, готовой осесть на подходящий субстрат, что для построения моде-
ли не имеет значения) заведомо избыточен (Максимович, 2004) и в общем случае не
зависит от пополняемого поселения, поэтому нас не будут интересовать величины F.
Положим их равными нулю для неполовозрелых особей
и равными единице для по-
ловозрелых.
Займемся вычислением величины P.
До сих пор функции, описывающие дифференциальную смертность живых орга-
низмов, не разработаны, поэтому в биологической литературе для этой цели чаще
всего используется распределение Вейбулла, которое описывает интенсивность отка-
зов различных устройств (см., например, Барлоу, Прошан, 1969). Это распределение
достаточно удобно и широко
используется в технике, но его теоретическое обосно-
вание не базируется на биологических предпосылках. Параметры функции, описы-
вающей распределение Вейбулла, затруднительно вычислить аналитически из фак-
тических данных, но главный ее недостаток заключается том, что они не объяснимы
с точки зрения биологических особенностей видов.
Эти соображения положены в основу попытки подобрать другую
функцию, кото-
рая описывала бы вероятность экземпляра определенного возраста дожить до сле-
дующего сезона размножения.
Представим себе, что общая дифференциальная смертность M складывается из
двух противоположных по смыслу процессов. Один из них – снижение с возрастом
детской дифференциальной смертности M
j
из-за увеличения сопротивляемости орга-
низма, которая возрастает за счет увеличения размеров, приобретения иммунитета и
др. Второй процесс – увеличение случайной дифференциальной смертности M
r
за
счет случайных причин. Половым созреванием и старением с их функциональными и
биохимическими перестройками пренебрежем. Так как оба процесса несовместны,
M = M
j
+ M
r
. (15)
Предположим, что первый процесс идет так, что за каждый год смертность падает
в определенное число раз. В этом случае снижение дифференциальной смертности с
возрастом будет равно
M
j,t+1
= M
j,t
/μ, (16)
где μ – коэффициент ежегодного снижения детской дифференциальной смертности.

Глава 5. ДИНАМИКА ОБИЛИЯ
92
Предположим далее, что удельный прирост случайной дифференциальной смерт-
ности постоянен. Тогда рост случайной смертности будет описываться логистиче-
ским законом. Поскольку его асимптота равна единице, то в рекуррентной форме
()
tl
tr
tr
Μe
eM
M
,
1
,
1,
1
−
+
+
=
λ
λ
, (17)
где λ – коэффициент удельного прироста случайной дифференциальной смертности.
При этом значение M
r,t
при t = 1 будет означать вероятность гибели в момент рожде-
ния или (в нашем конкретном случае) оседания.
Так как вероятность дожить до следующего возраста P равна 1 – M,
()
⎥
⎥
⎦
⎤
⎢
⎢
⎣
⎡
+
+−=
−
+
tr
tr
tj
t
Me
eM
M
P
,
1
,
,
1
1
1
λ
λ
μ
, (18)
где 0 ≤ P ≤ 1, поэтому при M
r
= 1 M
j
= 0. Значения P будем подставлять в диагональ
переходной матрицы Лесли.
Полученная функция достаточно гибка и удобна. Она базируется на обоснован-
ных биологических допущениях, и ее параметры имеют биологическое объяснение.
Аналитическое нахождение ее параметров тоже затруднено, но это ведь не останав-
ливает использование распределения Вейбулла. В случае же предложенной функции
ее параметры
несложно подобрать эмпирически, исходя из биологических особенно-
стей моделируемого вида.
Предположим, что на единицу площади субстрата может осесть N метаморфизи-
ровавших личинок. Эта величина будет показывать максимально возможное количе-
ство осевшего спата. Предположим далее, что количество реально осевшего спата
есть функция биомассы взрослых особей в поселении. У нас нет никаких данных
,
которые позволили бы подобрать вид этой функции, поэтому несколько условно вос-
пользуемся для расчетов степенной функций вида
f
adt
cBNN −= , (19)
где N
t
≤ N – количество спата, способное осесть в данном поселении в момент време-
ни t, B
ad
– биомасса взрослых особей в поселении, т. е. таких, для которых мы поло-
жили F = 1, c и f – коэффициенты. Поскольку у нас нет фактических данных, которые
позволили бы оценить параметр f, примем его равным единице, т. е. будем считать,
что моделируемая нами зависимость линейна. При этом условии коэффициент c по-
казывает,
какому количеству личинок препятствует осесть 1 г биомассы взрослых.
Одна из немногих попыток установить, влияет ли поселение взрослых моллюсков на
оседание молоди, предпринята П. Мёллером (Möller, 1986). Из его крайне скудных
данных следует, что указанная функция действительно близка к линейной, хотя экс-
перимент был поставлен довольно грубо и не доведен до конца.
Рост линейных
размеров опишем уравнением Л. фон Берталанффи (Bertalanffy,
1957, 1960) в форме, употребляемой для описания увеличения линейных размеров:
L
t
= L
0
+ L
∞
(1 – e
–kt
), (20)
где L
t
– длина раковины в момент времени t, L
0
– длина протоконха, L
∞
– максималь-
ная теоретически достижимая длина раковины, k – коэффициент.

МОДЕЛЬ ДИНАМИКИ ОБИЛИЯ
93
Для определения массы моллюсков в момент времени t воспользуемся хорошо из-
вестным уравнением простой аллометрии
1
(Huxley, 1932):
W
t
= W
0
+ αL
t
β
, (21)
где W
t
– масса моллюска в момент времени t, W
0
– масса только что прошедшей ме-
таморфоз личинки, α и β – коэффициенты.
Строго говоря, для определения массы было бы корректнее воспользоваться
уравнением весового роста Л. фон Берталанффи (Мина, Клевезаль, 1976; Алимов,
1981), однако в литературе, как правило, не приводятся значения его коэффициентов,
так как исследователей чаще интересует моделирование линейного роста, что не дает
возможности использовать опубликованные данные. В то же время параметры урав-
нения линейного роста и аллометрической зависимости хорошо известны для многих
видов.
Понятно, что величины L
0
и W
0
почти не влияют на работу модели из-за того, что
они крайне невелики численно. Не оказывают они влияния на результаты и при по-
лучении фактических данных, поэтому на практике ими обычно пренебрегают. Од-
нако мы сочли необходимым ввести их в модель для строгости.
Построенная модель позволяет легко рассчитывать демографический вектор
по-
селения двустворчатых моллюсков для любого произвольного момента времени и на
его основании получать значения плотности и биомассы как любой произвольно вы-
бранной возрастной когорты, так и всего поселения в целом. При желании модель
несложно дополнить расчетом продукционных характеристик, однако это не входило
в нашу задачу.
Для проверки работы построенной модели
на конкретных видах ряд данных был
почерпнут из литературы. Удалось найти данные только для четырех из тех видов,
которые обсуждались в начале этой главы – Modiolus modiolus, Mytilus edulis,
Macoma balthica и Mya arenaria. Параметры уравнения Л. фон Берталанффи были
взяты из следующих работ: Anwar et al., 1990 (Modiolus modiolus); Maximovich,
Guerassimova, 2003 (Mya arenaria); Максимович и др., 1992; Hummel et al. 1998
(Macoma balthica); Голиков и др., 1992 (Mytilus edulis).
Параметры уравнения простой
аллометрии: Максимович, 1978 (Mya arenaria); Максимович и др., 1993, а (Macoma
balthica); Кулаковский, Сухотин, 1986; Сухотин, 1989; Кулаковский и др., 1993
(Mytilus edulis). Дифференциальная смертность: Maximovich, Guerassimova, 2003
(Mya arenaria). Для Macoma balthica дифференциальная смертность была рассчитана
по данным, содержащимся в работе Н. В. Максимовича с соавторами (1993, а), а для
Mytilus edulis – в работах Э. Е. Кулаковского (2000) и Н. В.
Максимовича с соавтора-
ми (1993, б). Данных о дифференциальной смертности Modiolus modiolus не удалось
найти в литературе. Снижение смертности молодых особей было рассчитано на ос-
новании оригинальных данных (Flyachinskaya, Naumov, 2003), а рост случайной
смертности был подобран эмпирически таким образом, чтобы продолжительность
жизни соответствовала известным из литературы фактам (см. систематическую
часть). Параметры функции смертности подбирались эмпирически
таким образом,
чтобы она наилучшим способом описывала фактические данные. Максимально воз-
можная плотность спата Mya arenaria и Macoma balthica была взята из работ
1
Строго говоря, уравнение простой аллометрии было предложено для описания разной
скорости роста отдельных органов или частей тела по отношению к росту общей массы жи-
вотного. Однако и в случае увеличения массы с ростом линейных размеров аллометрические
зависимости отражаются на величине β, а именно: при β = 3 рост изометричен (пропорции тела
остаются
неизменными), при β < 3 аллометрия отрицательна (тело относительно удлинняется),
а при β > 3 – положительна (тело относительно укорачивается). Это обстоятельство позволяет
считать уравнение роста массы тела в зависимости от увеличения линейных размеров уравне-
нием простой аллометрии.

Глава 5. ДИНАМИКА ОБИЛИЯ
94
К.-П. Гюнтер (Günther, 1991, 1992, 1994). Эти данные были откорректированы на
основании наших наблюдений в губе Чупа (Кандалакшский залив). Для введения в
модель этой величины у Mytilus edulis и Modiolus modiolus использовались собствен-
ные наблюдения в Кандалакшском и Двинском заливах. Сведения о времени наступ-
ления половозрелости были взяты из работы А. Д. Наумова с соавторами (1987).
Длина
протоконха Modiolus modiolus взята из этой же работы, а протоконхов осталь-
ных видов – из работы Л. П. Флячинской (1999). Масса только что осевшего моллю-
ска – по нашим наблюдениям. Во всех тех случаях, когда в литературе приводились
данные по различным поселениям, использовались средние величины.
Коэффициент c функции (19) подбирался эмпирически, так как данные такого
ро-
да в литературе отсутствуют. В будущем, однако, можно надеяться, что после прове-
дения соответствующих исследований правильность его подбора будет проверена
фактическим материалом.
При тех параметрах, которые вводились в модель, она описывает поселения, об-
ладающие автоциклическими колебаниями плотности и биомассы. При исследовании
работы модели было установлено, что при других значениях вводимых
в нее пара-
метров она может описывать также поселения, в которых развиваются затухающие
циклы, или стабильные популяции, лишенные циклических изменений показателей
обилия. Так, например, попытка рассчитать коэффициент c функции (19) для Mya
arenaria из данных, приведенных П. Мёллером (Möller, 1986), дает значение числа
экземпляров молоди, которым препятствует осесть на квадратном метре 1 г биомас-
сы
взрослых, близкое к 9. При этом его значении модель предсказывает затухающие
колебания показателей обилия. Такой ход динамики возникает, если при низких зна-
чениях указанного коэффициента никогда не происходит полного прекращения оса-
ждения молоди в плотное поселение моллюсков.
Важно отметить, что даже весьма значительные изменения начального демогра-
фического вектора не сказываются на
результатах сколько-нибудь серьезно и изме-
няют только характеристики одного или нескольких начальных циклов. Ниже приве-
дены параметры, вводившиеся в модель (табл. 7), и полученные с ее помощью ре-
зультаты (табл. 8).
В табл. 9 приведены фактические данные, известные для рассматриваемых видов
из Белого моря по материалам автора и данным, опубликованным другими исследо-
вателями
. Видно, что в большинстве случаев результаты работы модели неплохо
воспроизводят основные параметры анализируемых поселений двустворчатых мол-
люсков. Исключение составляет Modiolus modiolus, для корректного моделирования
которого недостаточно фактического материала. Тем не менее, если мы сравним по-
лученные для него результаты не со средними параметрами его поселений, а с наи-
более плотными (см.
раздел об этом виде в систематической части), то в этом слу-
чае получим лучшее соответствие результатам моделирования. В плотных поселени-
ях средняя биомасса этого вида составляет 2873 г/м
2
, средняя плотность поселения –
1967 экз./м
2
, а средняя масса одного экземпляра – 1.46 г.
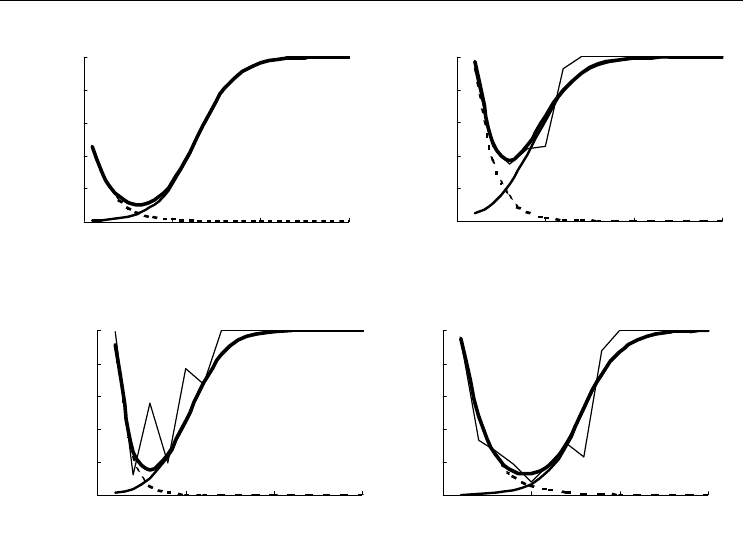
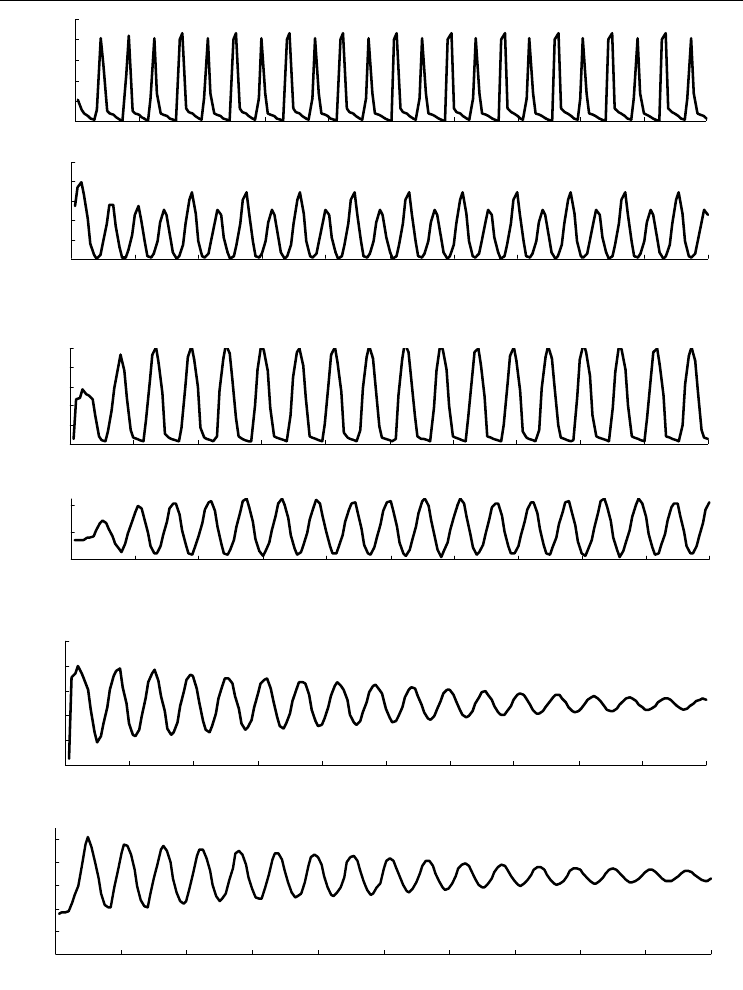
На рис. 46 и 47 приведены графики дифференциальной смертности всех четырех
видов и многолетняя динамика плотности и биомассы, сгенерированная моделью.
Обращает на себя внимание, что значение смертности Modiolus modiolus в первый
год жизни существенно ниже, чем у остальных видов. Скорее всего, это объясняется
недостаточным объемом материала, по
которому был рассчитан этот показатель.
Возможно также, что именно это обстоятельство явилось причиной того, что расчет-
ные характеристики поселений этого вида заметно отличаются от реальных. Для ос-
тальных трех видов удалось подобрать вполне приемлемые значения параметров
функции дифференциальной смертности. Исключение составляет Macoma balthica,
МОДЕЛЬ ДИНАМИКИ ОБИЛИЯ
95
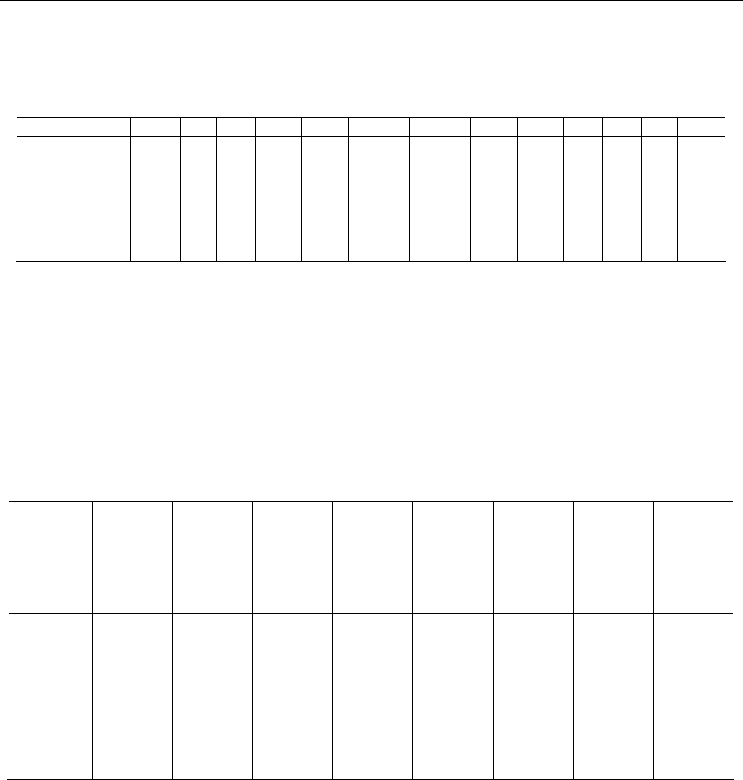
Таблица 7
Входные параметры модели развития плотного поселения
двустворчатых моллюсков
Input parameters for the model of bivalve dense settlement development
Вид N A
m
L
0
L
∞
k W
0
α β M
r,0+
λ μ
0
μ c
Modiolus
modiolus
1000 5 0.30 115.0 0.050 0.00002 0.00015 2.964 0.005 0.45 0.45 1.5 2
Mytilus edulis 10000 3 0.35 73.6 0.138 0.00002 0.00010 2.964 0.050 0.85 0.9
2
2.5 50
Macoma
balthica
2000 2 0.30 28.1 0.129 0.00002 0.00016 2.960 0.015 0.99 0.9
0
4.0 1000
Mya arenaria 1000 4 0.30 55.3 0.158 0.00002 0.00018 2.750 0.002 0.90 0.95 2.0 16
Примечание. A
m
– возраст наступления половозрелости, μ
0
– доля молоди, погибающей в
первый год жизни. Обозначения остальных столбцов – см. формулы (13) – (21). Размерность
величины N – экз./м
2
, c – экз./г, A
m
– годы, L
0
и L
∞
– мм, W
0
– г, α – г/мм, D
l,0+
, μ
0
и μ – доли
единицы. Остальные величины безразмерные. Величина A
m
означает номер текущего года
жизни, а не возраст в полных годах, поэтому ее значение (например, для Modiolus modiolus) в
традиционной записи было бы 4+.
Таблица 8
Параметры развития плотного поселения двустворчатых моллюсков,
полученные в результате моделирования
Parameters of bivalve dense settlement development computed by the model
Вид
Средняя
биомасса,
г/м
2
Средняя
плотность,
экз./м
2
Плотность
половозре-
лых экзем-
пляров,
экз./м
2
Средняя
масса
э
кземпляра
,
г
Средняя
масса
половозре-
лого
экземпля-
ра, г
Средняя
продолжи-
тельность
жизни в
поселении,
годы
Макси-
мальная
продолжи-
тельность
жизни,
годы
Продолжи-
тельность
цикла,
годы
Modiolus
modiolus
1814.275 828.5 299.9 2.190 6.050 15 25 17
Mytilus
edulis
2181.804 39767.8 655.7 0.055 3.328 8 12 8
Macoma
balthica
13.230 512.5 78.1 0.026 0.169 8 11 9
Mya
arenaria
55.082 404.3 25.0 0.136 2.204 9 15 11
для которой было бы лучше воспользоваться значением λ = 1.1 вместо использован-
ного в модели значения 0.99. Это, однако, было сделано сознательно. Указанное из-
менение данного параметра очень слабо сказывается на общих характеристиках по-
селения, генерируемых моделью, однако упрощает общий ход изменений биомассы,
поэтому для расчетов динамики показателей обилия этого вида использовано то
зна-
чение этого показателя, которое приведено в табл. 7 для того, чтобы продемонстри-
ровать, что модель может описывать и более сложные циклические процессы (см.
график динамики биомассы Macoma balthica на рис. 47).
Продолжительность циклов, рассчитанных при помощи модели, устанавливалась
методом компонентной фильтрации, так как деление числа лет на число пиков дает
небольшую ошибку
из-за неточности снятия данных с графика. Как и ожидалось, во
всех случаях только один тренд объяснялся значимой дисперсией: циклический, от-
вечающий сгенерированной моделью динамике. Продолжительность полученных
циклов приведена в табл. 8. Для Macoma balthica компонентная фильтрация показы-
вает два цикла длиной в 9 и 17 лет. Оба цикла хорошо видны на рис. 47. Доля дис
-
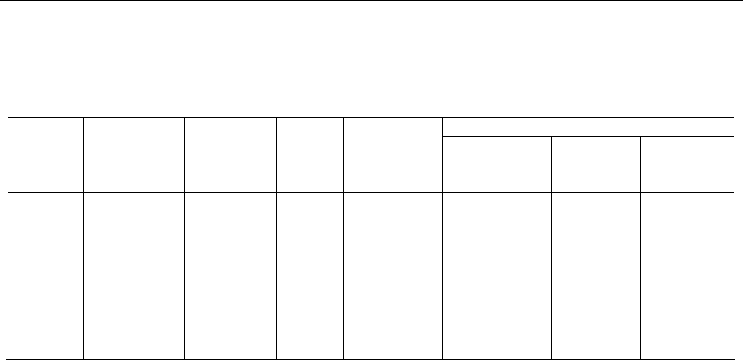
Глава 5. ДИНАМИКА ОБИЛИЯ
96
Таблица 9
Параметры плотных поселений двустворчатых моллюсков,
по материалам автора и литературным данным
Parameters of bivalve dense settlement after original and published data
Продолжительность цикла, годы
Вид
Средняя
биомасса, г/м
2
Средняя
плотность,
экз./м
2
Средняя
масса
экзем-
пляра, г
М
аксимальна
я
продолжи-
тельность
жизни, годы
данные автора,
Луканин и др.,
1990
Максимо-
вич, 1991
Герасимова,
2001
Modiolus
modiolu
s
772.7±247.5 53.3±15.6 13.4 Нет данных Нет данных
Нет дан-
ных
Нет данных
Mytilus
edulis
4884.4±499.2 16079±7293 0.3 10÷15 6÷12
Нет дан-
ных
5÷6
Macoma
balthica
20.1±1.67 433.6±60.0 0.046 8 5÷6 7÷9 4÷5
Mya
arenaria
62.8±9.7 275.7±100.7 0.228 15 >20
Нет дан-
ных
16÷17
Примечание. Данные о среднем обилии Modiolus modiolus и Mytilus edulis в Белом мо-
ре – см. систематическую часть. Данные о продолжительности жизни в Белом море – там же.
Продолжительность цикла Mytilus edulis
– см. Луканин и др., 1990. Данные о среднем обилии
Macoma balthica и Mya arenaria – по материалам многолетних наблюдений в губах Сельдяная
и Медвежья. Циклические изменения показателей обилия Macoma balthica в этих губах про-
слеживаются нечетко, так как они сильно замаскированы ярко выраженной сезонной динами-
кой (см. предыдущий раздел). Однако, если взять в анализ только
летние данные, то компо-
нентная фильтрация обнаруживает довольно четкий 5 – 6-летний цикл. Продолжительности
наблюдений (17 лет) недостаточно для того, чтобы надежно определить длительность цикла у
Mya arenaria, хотя он и заметен достаточно неплохо по основному тренду (см. предыдущий
раздел).
персии, объясняемая вторым циклом, статистически незначима, и поэтому он не
включен в таблицу. Однако следует иметь в виду, что при других начальных пара-
метрах модель может сгенерировать и вполне достоверную вторичную цикличность.
Так, в процессе исследовании модели при введении в нее исходных параметров,
сходных с теми, которые использовались для моделирования M. balthica
, были полу-
чены близкие результаты, но за одним высоким пиком биомассы следовали два низ-
ких, а не один, как показано на рисунке. В этом случае доля дисперсии, объясняемая
вторичным циклом, была статистически значимой на уровне вероятности P
2
.
Таким образом, модель дает основание считать, что зависящие от плотности по-
селения факторы, препятствующие оседанию молоди, могут вызывать автоцикличе-
ские колебания показателей обилия двустворчатых моллюсков, причем, по-
видимому, цикличность, вызываемая такой причиной, может быть и не столь про-
стой. Естественно, что в природе на подобный процесс накладывается действие раз-
личных
биотических и абиотических факторов, так что строгость циклов неминуемо
должна нарушаться. Моделирование такого воздействия, а также последствий моди-
фикации среды плотным поселением двустворчатых моллюсков представляет собой
отдельную задачу, хотя и вполне возможную. Основные сложности на этом пути нам
видятся в недостаточной изученности обсуждаемых явлений.
Модель, несомненно, может быть усовершенствована. Так, не
исключено, что для
описания случайной дифференциальной смертности стоит подобрать другую функ-
цию. Логистический закон подкупает своей простотой, однако имеет тот недостаток,
что эта функция симметрична относительно точки перегиба. Хотя в рассмотренных
случаях удалось подобрать вполне удовлетворительные значения ее параметров, не
МОДЕЛЬ ДИНАМИКИ ОБИЛИЯ
97
Modiolus modiolus Mytilus edulis
Macoma balthica Mya arenaria
Рис. 46. Графики дифференциальной смертности, построенные для всех четырех видов.
Тонкая линия – фактические данные (для Modiolus modiolus они отсутствуют); пунктирная линия – падение
ювенильной смертности; сплошная линия – рост случайной смертности; толстая линия – общий ход диф-
ференциальной смертности. По оси абсцисс – возраст моллюсков в годах, по оси ординат – вероятность
для моллюска данного возраста не дожить до следующего года.
Fig. 46. Plots of differential mortality rate for the four species
Thin line – experimental data (they are absent for Modiolus modiolus); doted line – decreasing of juvenile differ-
ential mortality rate; solid line – increasing of random differential mortality rate; thick line – total differential
mortality rate. X-axis – age (years), Y-axis – probability to die during the current year.
исключено, что при изучении других видов это может затруднить использование мо-
дели. Второй момент, который, вне всякого сомнения, требует доработки,– это функ-
ция влияния взрослых моллюсков на оседание молоди. Нет никакой уверенности в
том, что на самом деле она линейна, хотя на первый взгляд такая гипотеза представ-
ляется логичной. Вопрос этот
требует дополнительного исследования, для чего необ-
ходимо проведение экспериментальных наблюдений.
Впрочем, и в том виде, в котором она существует в настоящее время, предлагае-
мая модель дает достаточно хорошие результаты и может оказаться полезной при
прогнозировании развития плотного поселения двустворчатых моллюсков в тех слу-
чаях, когда по тем или иным причинам
нет возможности для его длительного наблю-
дения, а основные необходимые параметры могут быть получены в процессе кратко-
временного исследования
.
Приведенные графики динамики показателей обилия наглядно объясняют тот
факт, что исследователи часто сталкиваются с аномальным демографическим векто-
ром, и то, что при не достаточно большом количестве проб можно вообще не встре-
тить в природе молодь некоторых видов. В тех случаях, когда обильное поселение
0,0
0,2
0,4
0,6
0,8
1,0
051015
0,0
0,2
0,4
0,6
0,8
1,0
051015
0,0
0,2
0,4
0,6
0,8
1,0
051015
0,0
0,2
0,4
0,6
0,8
1,0
0102030

Глава 5. ДИНАМИКА ОБИЛИЯ
98
Modiolus modiolus
Mytilus edulis
Рис. 47. Многолетняя динамика обилия четырех видов, сгенерированная моделью.
Для каждого вида приведено два графика: вверху – плотность поселения, внизу – биомасса. По оси абсцисс
время в годах, по осям ординат: на верхних графиках – плотность поселения, экз./м
2
, на нижних – биомас-
са, г/м
2
. Для Mya arenaria приведены графики, рассчитанные при различных значениях коэффициента c
(уравнение 19), с целью продемонстрировать широкие возможности модели. Продолжение рисунка – на
следующей странице.
Fig. 47. Long-term dynamics of abundance in the four species generated by the model.
Two plots for each species display: above – density, below – biomass. X-axis – time (years), Y-axes: upper plots –
density (specimen/m
2
), lower plots – biomass (g/m
2
). Dynamics using different values of coefficient c (equation
19) was computed for Mya arenaria to demonstrate the wide possibility of the model. The figure is to be contin-
ued on the next page.
0
500
1000
1500
2000
2500
0 20 40 60 80 100 120 140 160 180 200
0
1000
2000
3000
4000
5000
0 20 40 60 80 100 120 140 160 180 200
0
20000
40000
60000
80000
100000
0 20 40 60 80 100 120 140 160 180 200
0
1000
2000
3000
4000
5000
0 20 40 60 80 100 120 140 160 180 200
МОДЕЛЬ ДИНАМИКИ ОБИЛИЯ
99
Macoma balthica
Mya arenaria (c = 16)
Mya arenaria (c = 9)
Рис. 47. Продолжение.
Объяснения приведены на предыдущей странице.
Fig. 47. Continuation.
For explanations see previous page.
0
500
1000
1500
2000
2500
0 20 40 60 80 100 120 140 160 180 200
0
10
20
30
40
50
0 20 40 60 80 100 120 140 160 180 200
0
200
400
600
800
1000
0 20 40 60 80 100 120 140 160 180 200
0
50
100
0 20 40 60 80 100 120 140 160 180 200
0
200
400
600
800
1000
0 20 40 60 80 100 120 140 160 180 200
0
20
40
60
80
100
0 20 40 60 80 100 120 140 160 180 200