Мешкова Н.Н., Федорович Е.Ю. (сост.) Хрестоматия по зоопсихологии и сравнительной психологии
Подождите немного. Документ загружается.

Психология дождевого червя
61
вей только слабым красным светом. Очень слабый белый свет притягивает дожде-
вых червей. Иногда в лунную ночь наблюдаются далекие путешествия дождевых
червей. Однако они неспособны отыскать брошенную ими норку и поэтому с
наступлением дня роют новую. Утром черви прячутся в своих норках.
Это животное обладает очень разнообразными органами чувств и на некото-
рые раздражители реагирует уже при минимальной интенсивности. Дождевой червь
имеет различные средства для познавания присущим ему способом окружающего
мира; он может анализировать влияния внешней среды и накапливать приобре-
тенный опыт. Жизнь червя кажется нам монотонной и бедной только потому, что
мы еще не изучили ее как следует. Очень сложное строение нервной системы,
обилие и разнородность связей между ее элементами, изученных лишь в неболь-
шой степени, показывают, что мы имеем дело с животным, которому присущи
многообразные восприятия.
Ж. И. Резникова
ПОВЕДЕНИЕ МУРАВЬЕВ НА КОРМОВОМ УЧАСТКЕ
1
Функционирование такой сложной системы, как многовидовое сообщество
муравьев, во многом определяется характером поведения и взаимодействия осо-
бей на кормовом участке. Появляется все больше данных в пользу того, что дей-
ствия муравьев, главным образом, социально обусловлены (Захаров, 1972, 1974,
1975). К настоящему времени многое известно о различных формах координации
деятельности внегнездовых рабочих, а также о способах добывания пищи и об
особенностях ориентации. Однако за пределами исследований остались некото-
рые важные вопросы поведения муравьев, познание которых необходимо для раз-
работки концепции сообщества социальных насекомых.
Одна из наименее изученных сторон жизни муравьев — индивидуальное поведе-
ние особей и роль индивидуума в жизни семьи. До сих пор основное внимание иссле-
дователей привлекало групповое поведение. Оценка внутривидовой и межвидовой
изменчивости индивидуального поведения муравьев пока практически отсутствует.
Нет экспериментальных работ, посвященных исследовательскому поведению
муравьев в естественных условиях; в то же время это — важная форма деятельно-
сти, присущая всем животным. Многие этологи склонны рассматривать его как
фундамент всего поведения (Шовен, 1972). Изучение исследовательского поведе-
ния в сравнительно-видовом аспекте может пролить свет на возможности исполь-
зования биологического потенциала на уровне многовидовых сообществ муравь-
ев. То же касается способности муравьев к научению, поскольку наши
представления о сложности задач, с которыми в природных ситуациях сталкива-
ются муравьи, и о способах их решения еще явно недостаточны.
Наконец, едва ли не самой интригующей стороной социальной жизни муравь-
ев является их «язык». До сих пор неясно, могут ли муравьи использовать дистан-
ционное наведение на источник пищи, подобно пчелам, какое количество ин-
формации они могут передавать и какие факторы определяют выбор того или
иного способа обмена сведениями.
В данной главе приводятся лишь первые подходы к решению этих вопросов,
отнюдь не претендующие на полноту исследования.
ИНДИВИДУАЛЬНОЕ ПОВЕДЕНИЕ МУРАВЬЕВ
Среди немногих работ, посвященных изучению индивидуального поведения му-
равьев, большинство проведено в лабораторных условиях и посвящено главным об-
разом функциональному разделению особей в семье и различиям в уровнях их актив-
ности. Я. Добжанская (Dobrzanska, 1958), используя индивидуальное мечение, впервые
поставила ряд экспериментов по выявлению связи поведения муравьев F. polyctena и
F. sanguinea с выполняемыми ими функциями. Оказалось, что наименее раздражимые
особи несут функции, не требующие подвижности; другие выполняют работы, свя-
занные с частой сменой задач и с постоянным активным движением.
1
Резникова Ж.И. Межвидовые отношения муравьев. Новосибирск: Наука, 1983. С. 51—91
(с сокр.).
Поведение муравьев на кормовом участке
63
Г. Веррон (Verron, 1974, 1976) исследовал локомоторную активность муравьев
Lasius niger, Lasius flavus и Formica fusca, помещая их на разграфленный манеж и
учитывая число делений, пробегаемых каждой особью, а также время, затраченное
на передвижение и остановки. Оказалось, что часть особей отличается большой ва-
риабельностью этих этологических признаков, остальных можно условно разделить
на сильноактивных (в семье их меньшинство), средне- и низкоактивных. Сходные
данные были получены на Tapinoina erraticum в результате наблюдений за транспор-
тировкой куколок из чрезмерно нагретой в прохладную зону (Meudes, 1973).
У Myrmica laevinodis исследовались индивидуальные различия в уровне актив-
ности и агрессивности (Le RouxA. М., Le Roux С., 1979). Оказалось, что в среднем
50% рабочих совершенно неактивны и не выходят из гнезда, 30% активны, но не
агрессивны, а 20% и активны и агрессивны, причем последняя группа может
быть еще разделена на малоагрессивных и агрессивных.
В естественных условиях индивидуальное поведение муравьев изучалось в пла-
не функциональной дифференциации особей (Oekland, 1931; Kill, 1934; Stager,
1937; Otto, 1962; Boulig, Janzen, 1969). Так, К. Хорстманн (Horstmann, 1973) выде-
ляет среди внегнездовых рабочих F. polyctena три профессиональные группы: вос-
ходители на деревья, специализированные охотники на почве и сборщики
строительного материала. Собственно индивидуальной изменчивости поведения
внегнездовых рабочих на кормовом участке посвящена только работа Ф. Бернарда
(Bernard, 1971), описавшего различия в технике сбора семян у Messor barbara:
большинство муравьев таскают семена одно за другим, пробираясь в густых за-
рослях, 8—9% срезают и тащат в гнездо целый стебелек с 20—60 семенами, и
лишь 1—2% сборщиков умеют сбрасывать зерна вниз с вершины холма.
Соотношение различных элементов поведения у муравьев разных функциональ-
ных групп до сих пор остается неисследованным у подавляющего большинства видов.
В частности, лишь в немногих работах применялся метод построения этограмм
(Harkness M.L., Harkness R.D., 1976; Higashi, 1978; Brandao, 1978).
<...> Мы оцениваем индивидуальное поведение муравьев, согласно принци-
пам фиксирования и классификации элементов поведения Е.Н. Панова (1978) по
соотношению элементов поведения и, кроме того, по характеру траекторий пе-
редвижения на кормовом участке (Резникова, 1974а, 1977а; Резникова, Шиллерова,
1978, 1979). Большой вклад в эти исследования внесла О.А. Шиллерова (Богатыре-
ва) (1980). Вопросам индивидуального поведения муравьев разных видов посвя-
щена ее дипломная работа (1980).
Наблюдения проводились в 1971, 1976—1979 гг. в ассоциациях I и II
1
. Предвари-
тельно большую часть внегнездовых рабочих в выбранных семьях метили индивиду-
альными метками с помощью цветных нитролаков, смешанных с нитроклеем. Метка
держалась на муравьях до полутора месяцев. Код составлялся так, чтобы стирание
краски можно было контролировать: например, желтая точка на груди муравья озна-
чала, что белой точке на левой стороне брюшка должна соответствовать красная
точка на правой. Все кодовые сочетания заносились в журнал, и одновременно с
мечением новых муравьев производилось подновление старых меток. Для индиви-
дуального отлова муравьев проворных видов применялись конические колбочки с
прорезанным в дне отверстием.
Наблюдения проводили в периоды высокой активности муравьев с момента вы-
хода из гнезда до возвращения в него. Для статистической обработки отбирали «пол-
ные» рейсы, исключая муравьев, потерянных наблюдателями до возвращения в гнездо.
1
Ассоциация — здесь: группа видов муравьев, обитающих на общей территории (прим. ред.-сост.).
64
Ж.И. Резникова
Два наблюдателя хронометрировали продолжительность поведенческих реакций му-
равьев с помощью двухстрелочного секундомера и одновременно вычерчивали тра-
екторию передвижения особи на плане кормового участка. Для этого кормовой уча-
сток предварительно был разбит на квадраты со стороной 0,5 м. Всего проанализировано
640 этограмм и траекторий муравьев, что соответствует примерно 100 ч наблюдений.
Мы старались проследить как можно большее число рейсов одной и той же
особи за любой срок — от одного дня до месяца. Такие наблюдения в природе,
насколько нам известно, не проводились. В то же время без них очень трудно
представить себе место муравья в семье и изменчивость его поведения. Это оказа-
лось сложной задачей: многие меченые муравьи теряли метку или терялись сами,
погибали на кормовом участке, становясь добычей пауков или других муравьев, а
чаще всего на неопределенный срок исчезали в гнезде. Поэтому мы очень доро-
жили теми особями, которые исправно выходили на кормовой участок и давали
нам возможность судить о степени постоянства функций муравьев на кормовом
участке и об их территориапьном консерватизме. Таким муравьям мы давали име-
на и старались использовать каждую встречу с ними в своих целях. Иногда удава-
лось наблюдать до 20 рейсов одного фуражира в течение месяца.
Пользуясь классификацией Е.Н. Панова (1978), мы выделяли в качестве реги-
страционных элементов единицы поведения II уровня — несложные блоки локо-
моций (бег, транспортировка добычи, обследование травостоя) и «позы» (рез-
кий поворот на месте, ощупывание предмета). Правда, в некоторых случаях трудно
решить, к какому уровню принадлежит рассматриваемая единица поведения: ко
II или к III. Так, обследование небольших площадок, травостоя и т.п. представля-
ет скорее «комплекты поз», однако ввиду их несложности нам кажется нецелесо-
образным рассматривать их отдельно. Более или менее упорядоченные последова-
тельности (ориентировочные окологнездовые, охотничьи рейды и т.п.) мы
рассматривали в качестве поведенческих ансамблей.
Для количественной оценки траекторий (Резникова, 1980) были выбраны сле-
дующие параметры:
1) отдаление — расстояние от гнезда до наиболее удаленной от него точки
траектории;
2) степень извилистости — отношение отдаления к общей длине пробега;
3) отношение числа самопересечений траектории к общей длине пробега;
4) максимальная ширина охвата участка траекторией;
5) площадь охвата участка траекторией;
6) амплитуда отклонения траектории от прямой, проведенной от гнезда до
наиболее удаленной от него точки (среднее из 10 измерений, произведенных на
равном расстоянии).
ВНУТРИВИДОВАЯ ИЗМЕНЧИВОСТЬ ИНДИВИДУАЛЬНОГО
ПОВЕДЕНИЯ
Наблюдения за тремя семьями Formica cunicularia (Резникова, Шиллерова, 1978)
позволили выделить две относительно автономных группы: трофобионтов и актив-
ных фуражиров
1
(соотношение их численности примерно 1 : 2). И те и другие могут
1
Трофобионты — группа рабочих муравьев с узкой специализацией — собиратели выделе-
ний тлей.
Активные фуражиры — рабочие муравьи, активно разыскивающие корм и производящие
мобилизацию пассивных фуражиров к источникам корма (прим. ред.-сост.).
Поведение муравьев на кормовом участке
65
заниматься транспортировкой строительного материала. Серии дальних рейсов (их
бывает 3—5 за день) обычно начинаются и заканчиваются кратковременными выхо-
дами в пригнездовую зону.
Трофобионты, посещающие наземные колонии тлей на переферии кормового
участка, используют в большинстве рейсов один и тот же путь. Около 90% време-
ни они затрачивают на бег по поверхности почвы или по стеблям трав, остальное
время приходится на чистку тела или ориентировочные реакции. Муравьи, посе-
щающие кормовые пещерки (с подземными колониями тлей и цикадок) в сред-
ней зоне кормового участка, обладают более изменчивым и разнообразным пове-
дением. Они тратят значительно больше времени (до 25%) на обследование
территории (ощупывание мелких предметов, прикорневых частей растений, об-
следование почвы), что, возможно, связано с поиском новых кормовых пещерок.
Эти муравьи изредка могут совершать охотничьи рейсы.
Активные фуражиры («охотники») обрабатывают широкие секторы кормового
участка и, видимо, хорошо знакомы со всей территорией в целом. На пути фуражи-
ра, несущего добычу, мы последовательно ставили преграды, перегоняя его таким
образом в самый дальний сектор по отношению к тому, где этот муравей был впервые
замечен. Из 14 особей лишь 2 затратили на возвращение в гнездо более 10 мин,
остальные находили его без всяких затруднений. Ранее такие опыты, позволяющие
предположить наличие «внутреннего плана» территории у активных фуражи-
ров, проводились нами у Cataglyphis aenescens и А. Буркхалтером (Burkhalter, 1972),
М. и Р. Харкнессами (Harkness М., Harkness R., 1976) у Calaglyphis bicolor.
Характер поведения и конфигурация траекторий наиболее лабильны у активных
фуражиров. Траектории их отличаются большой извилистостью (0,11±0,07), тогда
как при сборе пади
1
этот показатель близок к единице. Во время охоты наблюдаются
и наибольшая площадь охвата территории — 9,62 ± 2,3 м
2
(при сборе пади — 1,19 ±
0,6 м
2
, при окологнездовом патрулировании — 1,6 ± 0,1) и максимальная амплитуда
отклонения (соответственно 2,13 + 0,2 м; 0,7 ± 0,3 м и 0,4 ± 0,08 м).
При поиске добычи муравьи либо передвигаются широким челноком, либо пос-
ледовательно обследуют небольшие участки на разном удалении от гнезда. Поведен-
ческий репертуар охотника включает обыскивание травостоя, ощупывание предме-
тов, охоту и транспортировку добычи. Чаще всего муравей находит мелких насекомых
(цикадок, клопов) в куртинах злаков или в укрытиях на поверхности почвы, сжима-
ет их в челюстях и сразу транспортирует в гнездо. В поисках добычи активные фура-
жиры могут в течение нескольких дней или нескольких периодов активности обраба-
тывать один и тот же участок и, наоборот, в один день обследовать разные секторы
территории.
Спектры поведенческих реакций муравьев во время охотничьих рейсов весьма
изменчивы. Однако можно выделить и присущие каждому муравью особенности.
Так, на рис. 1, а, б видно, что муравей М
;
очень подвижен и почти не тратит
время на чистку тела и обследование травостоя и мелких участков на поверхности
почвы; напротив, у муравья М
2
не менее 30% времени уходит на обследование
травостоя или ориентировочные реакции.
У F. pratensis поведение особей связано с глубокой специализацией. Для этого
вида характерно четкое вторичное деление территории и наличие узкопрофесси-
ональных групп фуражиров. Поэтому индивидуальная изменчивость поведения
отдельных особей выражена значительно слабее, чем у F. cunicularia. При сравне-
1
Падь — сладкие выделения тлей, которыми питаются муравьи (прим. рад.-сост.).
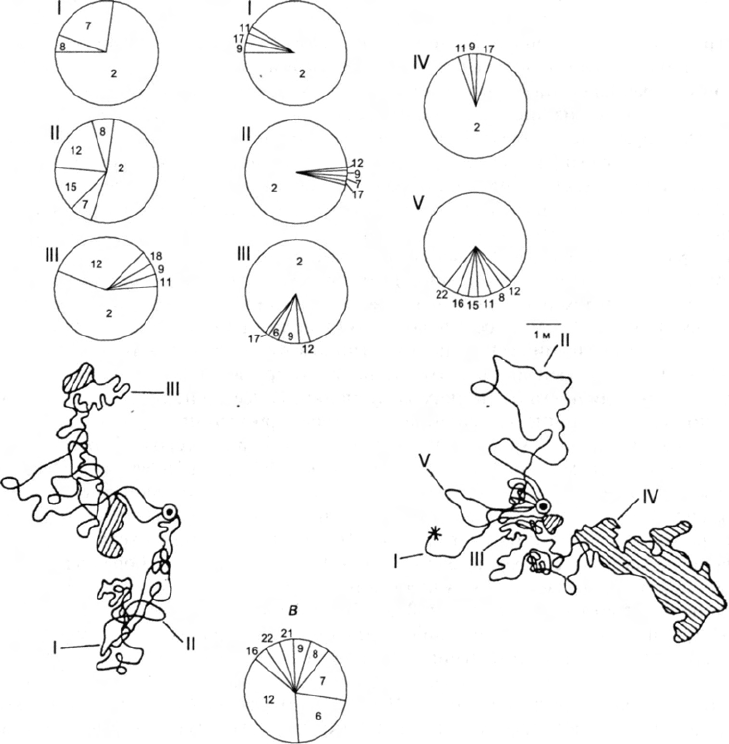
66
Ж.И. Резникова
а б
Рис. 1. Индивидуальные траектории и этограммы двух особей Formica cunicularia М,
(а) и М
2
(б) в разные дни и часы и усредненная этограмма для 50 активных фуражиров
лугового муравья (в):
Элементы поведения для рис. 20—22: 1 — бежит по прямой, 2 — перемещается плав-
ными зигзагами, 3 — бежит с кратковременными остановками, 4 — резко поворачивает,
5 — останавливается, приподнимаясь на передних ногах, 6 — подробно обследует не-
большие участки почвы, 7 — стоит неподвижно, 8 — ощупывает предметы антеннами, 9—
стоит и шевелит антеннами, 10 — оглядывается, 11 — взбирается на возвышения и огля-
дывается, 12 — обследует травостой, 13 — перебирается через травинки, 14 — находит
добычу, 15 — умерщвляет добычу, 16 —транспортирует добычу, 17 — соприкасается ан-
теннами с другими муравьями, 18 — избегает контакта с другими муравьями, 19 —
обменивается пищей (трофаллаксис), 20 — стычка с другими муравьями, 21 — чистит
антенны, 22 — чистит тело и ноги; 1—V — номера траекторий движения и соответствую-
щие им этограммы: для М, 16/VIII в 17.05 (I), 16.35 (II) и 10.03 (III); для М
2
24/VIII в 16.05
(I), 10.20 (II), 15/VIII в 17,02 (III), 26,VIII в 11.15 (IV) и 26/VIII в 8.15 (V); на траекториях
заштрихованы области, которые муравьи тщательно обследовали
Поведение муравьев на кормовом участке
67
нии активных фуражиров оказалось, что в то время как в поведении каждой
особи F. cunicularia содержится почти полный набор актов, выделенных нами у
обоих видов, у F. pratensis в среднем их 6—8. Отличительная особенность поведе-
ния лугового муравья — большая доля времени, затрачиваемого на контакты с
другими муравьями на кормовом участке (рис. 1, б).
Formica uralensis экологически во многом сходен с F. pratensis (Stebaev, Rcznikova,
1972; Резникова, Самошилова, 1981), но организация кормового участка и поведение
особей черноголового муравья совершенно иные (см. ранее). Как уже отмечалось,
вторичное деление территории выражено очень слабо. Большинство внегнездовых
рабочих обрабатывают разные зоны кормового участка и, по-видимому, могут осу-
ществлять весь набор функций. Из 20 особей, наблюдавшихся многократно, лишь
один муравей совершал только охотничьи рейсы, а остальные совмещали их с посе-
щением кормовых пещерок и колоний тлей.
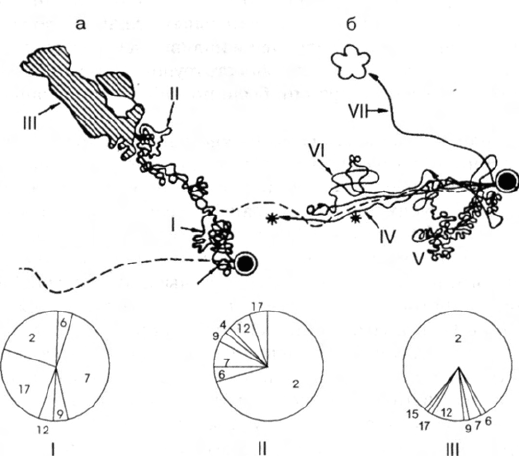
Типичный случай представлен на рис. 2. Муравей В 24/VII в 9.05 вышел для около-
гнездового ориентирования (траектория I). В 9.20 он выходил с охотничьим рейсом
(траектория II), причем часто контактировал с другими особями, обследовал расти-
тельность, останавливался для ориентировки. В 18.53 он охотился несколько дальше
от гнезда (траектория III). В другие дни этот муравей посещал кормовую пещерку на
дороге (траектория IV). 20/VII он дважды выходил на охоту в разные зоны кормового
участка (траектория V, VI), а также посещал кормовую пещерку (траектория VII).
Помимо этого, он участвовал в столкновении с семьей F. picea, находящейся в 9 м
от гнезда по ходу дороги. Характерно, что чем дальше муравьи отходят от гнезда, тем
больший процент времени в их поведенческом репертуаре занимают ориентировоч-
ные реакции и меньший — контакт с другими муравьями. При повторных выходах на
тот же маршрут доля ориентировочных актов уменьшается. <...>
Рис. 2. Траектории и этограммы одного из внегнездовых рабочих черноголового
муравья:
I — 24/VII в 9.05, II — в 9.20, III — в 18.53, IV—VII — 20/VII в разное время суток; штрихо-
вой линией показана фуражировочная дорога
68
Ж.И. Резникова
ГРУППОВОЕ ОБУЧЕНИЕ У МУРАВЬЕВ
Обучение, связанная с ним лабильность поведения и обмен информацией ле-
жат в основе механизмов межвидового взаимодействия. В данном разделе мы по-
пытаемся осветить вопросы, необходимые для дальнейшего изложения: каковы
изменчивость психических возможностей фуражиров одного и разных видов и
способность их к решению сложных задач?
Возможности муравьиного интеллекта давно занимали умы исследователей. По-
скольку долгое время господствовало мнение о том, что у насекомых вырабаты-
ваются лишь элементарные условные рефлексы, сложность задач, которые экс-
периментаторы предлагали муравьям, наращивали очень робко. Сама по себе
способность муравьев к запоминанию и научению была экспериментально проде-
монстрирована с помощью различных методик, начиная с классических экспе-
риментов Т. Шнейрлы, обучавшего их в лабиринтах (Schneirla, 1933, 1946), и
кончая работами последних лет (Длусский, 1967; Резникова, 1971, 1975; Плеханов,
Кауль, 1976; Мазохин-Поршняков, Мурзин, 1976, 1977; Мурзин, 1976, 1977). В част-
ности, при групповом обучении у F. polyctena удалось выработать сложные инстру-
ментальные условные рефлексы: муравьи должны были тянуть за одну из нитей,
чтобы получить доступ к кормушке, спрятанной за ширмой. Интересно, что на-
блюдались случаи, когда тянули за нитку одни муравьи, а проникали за ширму
другие, т.е. обучалась именно группа в целом (Rechtsteiner, 1968, 1971).
Приобретение опыта, в том числе основанного на подражании, имеет особенно
большое значение для муравьев, так как срок жизни одной особи достигает у некоторых
видов 7 лет (Otto, 1962), а в среднем срок жизни рабочих особей — 1,5—2,5 года
(Haskins С, Haskins Е., 1980), т.е. больше, чем у многих видов грызунов. В настоящее
время считается доказанным существование у муравьев сигнальной преемственнос-
ти
1
(Захаров, 1974). При решении задач, требующих объединенных усилий группы
особей, или задач, основанных на подражательных реакциях, должна проявляться
неоднородность психических возможностей и индивидуального опыта муравьев. Про-
явление индивидуальных вариаций и способность группы муравьев к решению сложных
задач, требующих логических операций, были почти не исследованы до настоящего
времени.
Разнокачественность психических возможностей фуражиров. Индивидуальные
различия в способностях и навыках животных привлекли внимание исследовате-
лей в основном в последнее время, когда начались попытки межвидовых количе-
ственных сравнений. До сих пор в этом плане внимание уделялось лишь позвоноч-
ным животным. Так, Д.М. Рамбо (Rumbaugh, 1968) (цит. по: Дьюсбери Д., 1981)
установил, что в группе беличьих обезьян (саймири) часть животных решает за-
дачи на «формирование установки» на уровне высших обезьян (более 90% пра-
вильных выборов при втором предъявлении), а часть оказывается вообще не спо-
собной к решению. К.Н. Благосклонов (1974) на примере мухоловок-пеструшек,
которым предлагались задачи с перемещением гнезд и с преодолением пугающе-
го препятствия (трубка, вставленная в леток), показал, что формы решения задач
очень различны и адекватны накопленному индивидуальному опыту.
У муравьев разнообразие стереотипов поведения связано прежде всего с кас-
товым полиморфизмом и наличием группового и возрастного полиэтизма, т.е.
фиксированных различий в функциях, выполняемых разными особями. Этот воп-
1
Сигнальная преемственность — обучение животного в группе, основанное на подражании
другим ее членам (прим. ред.-сост.).
Поведение муравьев на кормовом участке
69
рос довольно хорошо изучен и подробно обсуждался в монографии А.А. Захарова
(1972). Однако дифференциация особей может быть еще более глубокой. Извест-
но, что в небольших функционально однородных группах муравьев выделяются
«одаренные» особи, которые обладают хорошей памятью и играют роль активато-
ров при выполнении различных функций и организации групп (Combes, 1941,
цит. по: Захаров, 1972).
Различия в способностях и уровне активности рабочих особей можно на-
блюдать даже в относительно простых ситуациях, когда группа муравьев сталки-
вается с препятствием на пути к пище или гнезду. Такой опыт был проделан
нами в 1968 г. с трофобионтами F. polyctena, которые спускались по стволу бере-
зы к муравейнику. Ствол окружили кольцом пластилина с нафталином. Преодо-
ление этого препятствия не было хаотическим: группы из 6—7 фуражиров оста-
навливались перед кольцом и ожидали своего «вожака» — самого активного
муравья, который первым преодолевал препятствие и затем пробегал через кольцо
туда и обратно, сопровождая остальных муравьев. Возможно, что здесь прояви-
лись отношения доминирования-соподчинения, связывающие знакомых между
собой особей, которые используют перекрывающиеся поисковые участки.
Предположение о наличии индивидуальной иерархии в группах рабочих особей выте-
кает и из работы А. М. и Г. Руа (Le RouxA. М., Le Roux G., 1979). В группах из 8—10 особей
М. laevinodis наблюдалось постоянное соотношение активно-агрессивных, активно-ма-
лоагрессивных, активных и совершенно неактивных муравьев. По этим критериям мура-
вьев перефуппировали в четыре однородные группы. Оказалось, что совместное содер-
жание агрессивных особей приводит к некоторому снижению их агрессивности и
активности, а неагрессивных — к появлению среди них агрессивных, особей. В результате
во всех группах соблюдается примерно то же соотношение особей по уровню агрессивно-
сти, что и первичное. При восстановлении первоначальной ситуации проявляется тен-
денция к сохранению муравьями ранга, приобретенного после первой перегруппировки.
Значительное количество муравьев оставались полностью неагрессивными в течение всех
перегруппировок.
Сходные результаты получены Г. Верроном (Verron, 1977): одновозрастных особей
Lasius niger объединили в четыре группы по уровню активности, которую они
проявляли, перетаскивая песчинки. Удаление самых энергичных особей заставля-
ло остальных проявлять большую активность, а их присутствие, напротив, по-
давляло деятельность остальных.
Эти опыты дают основание полагать, что ранг особей и их поведение в группах
зависят от психофизиологических свойств и к тому же поддерживаются активным
взаимодействием. Такое взаимодействие было исследовано А.А. Захаровым (1981)
в тонком лабораторном эксперименте с малыми группами муравьев Formica rufa.
Оказалось, что индивидуальная борьба за доминирование выражается в увели-
чении двигательной активности соперничающих особей, а также в проявлениях
агрессивности и прямого противоборства. В частности, муравьи устраивают свое-
образные турниры, когда претендующий на первенство фуражир пытается занес-
ти соперника в гнездо. Два фуражира некоторое время толкают друг друга, стара-
ясь сложить напарника «чемоданчиком». Если это долго не удается ни одному из
них, муравьи разбегаются.
Изъятие особей-доминантов из групп со сложившейся поведенческой структу-
рой стимулирует быстрое линейное продвижение оставшихся особей по рангам.
При этом выход в лидеры не является формальным перемещением на освободив-
шееся место: именно в период своего утверждения в новой роли лидер резко
70
Ж.И. Резникова
увеличивает активность. Вполне возможно, что способность муравьев к обучению
коррелирует с их иерархическим положением в группе, обслуживающей опреде-
ленный участок территории, и с реактивностью по отношению к внутригруппо-
вым ранговым перемещениям.
Пока еще мало данных для того, чтобы сравнивать у разных видов соотноше-
ние числа особей, способных быстро справляться с новыми задачами, и особей,
не способных к этому. Однако можно полагать, что этот показатель связан со
способом использования кормового участка. При коллективной фуражировке до-
статочно лишь нескольким особям справиться с задачей, а остальные могут обу-
чаться в результате подражания. Если же муравьи действуют на кормовом участке
в одиночку, то существенные различия в их психических возможностях только
повлекут за собой неравномерное использование территории: сообразительные
особи будут полнее использовать ресурсы своего участка.
Такое предположение подтверждается опытами по коллективному обучению в
лабиринтах муравьев нескольких видов. Р. Шовен (Chauvin, 1963) устанавливал
лабиринты на фуражировочных дорогах F. polyctena. Это вид с наиболее явно вы-
раженным групповым поведением. В начале опыта только 10—20% муравьев пре-
одолевали лабиринт. Лишь спустя 10 ч, благодаря подражательным реакциям, число
их достигло 75—80%.
Аналогичный опыт был проведен нами с F. uralensis в степях Южной Хакасии
(1969). Лабиринт, предлагаемый муравьям, представлял собой несколько цилинд-
ров, вставленных один в другой так, чтобы входы в них располагались с противопо-
ложных сторон. В последнем внутреннем цилиндре помещалась приманка. До предъяв-
ления задачи муравьев прикармливали на площадке для наблюдений в лабиринте
первой сложности (один цилиндр). Площадки размещались в периферийной зоне
кормового участка. Для того, чтобы исключить возможность накопления муравьями
опыта, лабиринты разной сложности расставляли на разных площадках. Оказалось,
что в лабиринт второй сложности в течение первого часа наблюдений попадали 70%
из пытавшихся проникнуть туда муравьев; за то же время в лабиринт третьей слож-
ности попадали 10—25% особей, а четвертой — всего 1—2%.
Такой же эксперимент, проведенный с Cataglyphis setipes turcomanica (урочище
Акибай, ТССР, 1970 г.), выявил совсем иное соотношение: 95, 90 и 70% соответст-
венно. У муравьев этого вида кормовой участок организован по принципу одиночной
фуражировки, и большинство внегнездовых рабочих легко справлялись с задачей
самостоятельно.
Эксперименты, в которых одновременно предъявлялось большое число (50—100)
лабиринтов второй сложности, расставленных на всей площади участков наблюде-
ний (1200—1800 м
2
) в ассоциациях I—III, также выявили соответствие между ско-
ростью и характером обучения муравьев и способами организации фуражировки. Для
F. picea и F. cunicularia, действующих на территории в одиночку, такая задача вообще
не составила сложности, и они доставали приманку при первых же обнаружениях
лабиринта. Напротив, среди 20—40 особей F. pratensis, привлеченных к лабиринту,
лишь 2—3 муравья проникали к приманке в течение первых четырех часов наблюде-
ний. Впоследствии к ним присоединились еще 8—10 особей, и они вместе транспорти-
ровали приманку. Фуражиры М. scabrinodis и L. alienus действовали примерно одинако-
во: в лабиринте, находящемся вблизи гнезда, появлялись 2—3 разведчика, и в течение
0,5—1 ч они производили массовую мобилизацию рабочих (до 200 особей). <...>
Задачи, требующие логических операций. Высокий уровень психической организа-
ции муравьев вполне позволяет задуматься об их, способности усваивать логическую