Latchman. Eukariotic Transciption Factors
Подождите немного. Документ загружается.


CHAPTER 7
REGULATION OF TRANSCRIPTION
FACTOR SYNTHESIS
7.1 TRANSCRIPTION FACTOR REGULATION
Transcription factors play a central role in a number of biological processes,
producing, for example the induction of specific genes in response to parti-
cular stimuli, as well as controlling the cell type-specific or developmentally
regulated expression of other genes. The ability to bind to DNA (Chapter 4)
and influence the rate of transc ription either positively (Chapter 5) or nega-
tively (Chapter 6) are clearly features of many transcription factors that reg-
ulate gene expression in response to specific stimuli or in specific cell types.
Most importantly, however, such factors must also have their activity regulated
such that they only become active in the appropriate cell type or in response
to the appropriate stimulus, thereby producing the desired pattern of gene
expression.
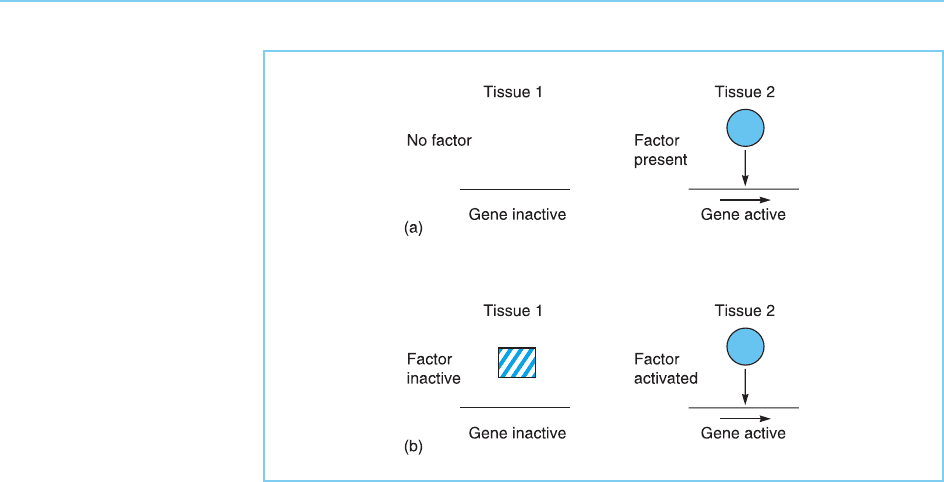
Two basic mechanisms by which the action of transcription factors can be
regulated have been described. These involve either controlling the synthesis
of the transcription factor so that it is made only when necessary (Fig. 7.1a) or
alternatively, regulating the activity of the factor so that pre-existing protein
becomes activated when required (Fig. 7.1b). This chapter considers the
regulation of transcription factor synthesis while Chapter 8 considers the
regulation of transcription factor activity.
7.2 REGULATED SYNTHESIS OF TRANSCRIPTION FACTORS
Regulating the synthesis of transcription factors such that they are only made
when the genes that they regulate are to be activated is an obvious mechanism
of ensuring that specific genes become activated only at the appropriate time
and place. This mechanism is widely used therefore, particularly for transcrip-
tion factors that regulate the expression of cell type-specific or developmen-
tally regulated genes. Specific examples of the regulated synthesis of
particular transcription factors which illustrate the role of this mechanism
in regulating cell type-specific or developmental gene expression are dis-
cussed in the following sections.
7.2.1 THE MyoD TRANSCRIPTION FACTOR
Probably the most novel approach to the cloning of the gene encoding a
transcription factor was taken by Davis et al. (1987), who isolated cDNA clones
encoding MyoD, a factor which plays a critical role in skeletal muscle-specific
gene regulation. They used an embryonic muscle fibroblast cell line known as
C3H 10T‰. Although these cells do not exhibit any differentiated character-
istics, they can be induced to differentiate into myoblast cells expressing a
number of muscle-lineage genes upon treatment with 5-azacytidine
(Constantinides et al., 1977). This agent is a cytidine analogue having a nitro-
gen instead of a carbon atom at position 5 on the pyrimidine ring and is
incorporated into DNA instead of cytidine. Unlike cytidine, however, it can-
not be methylated at this position and hence its incorporation results in
demethylation of DNA. As methy lation of DNA at C residues is thought to
play a critical role in transcriptional silencing of gene expression (for review
see Latchman, 2002), this artificial demethylation can result in the expression
of particular genes which were previously silent.
In the case of 10T‰ cells therefore this demethylation was thought to result
in the expression of previously silent regulatory loci that are necessary for
differentiation into muscle myoblasts. Several experiments also suggested that
212 EUKARYOTIC TRANSCRIPTION FACTORS
Figure 7.1
Gene activation mediated
by the synthesis of a
transcription factor only in
a specific tissue (a) or its
activation in a specific
tissue (b).
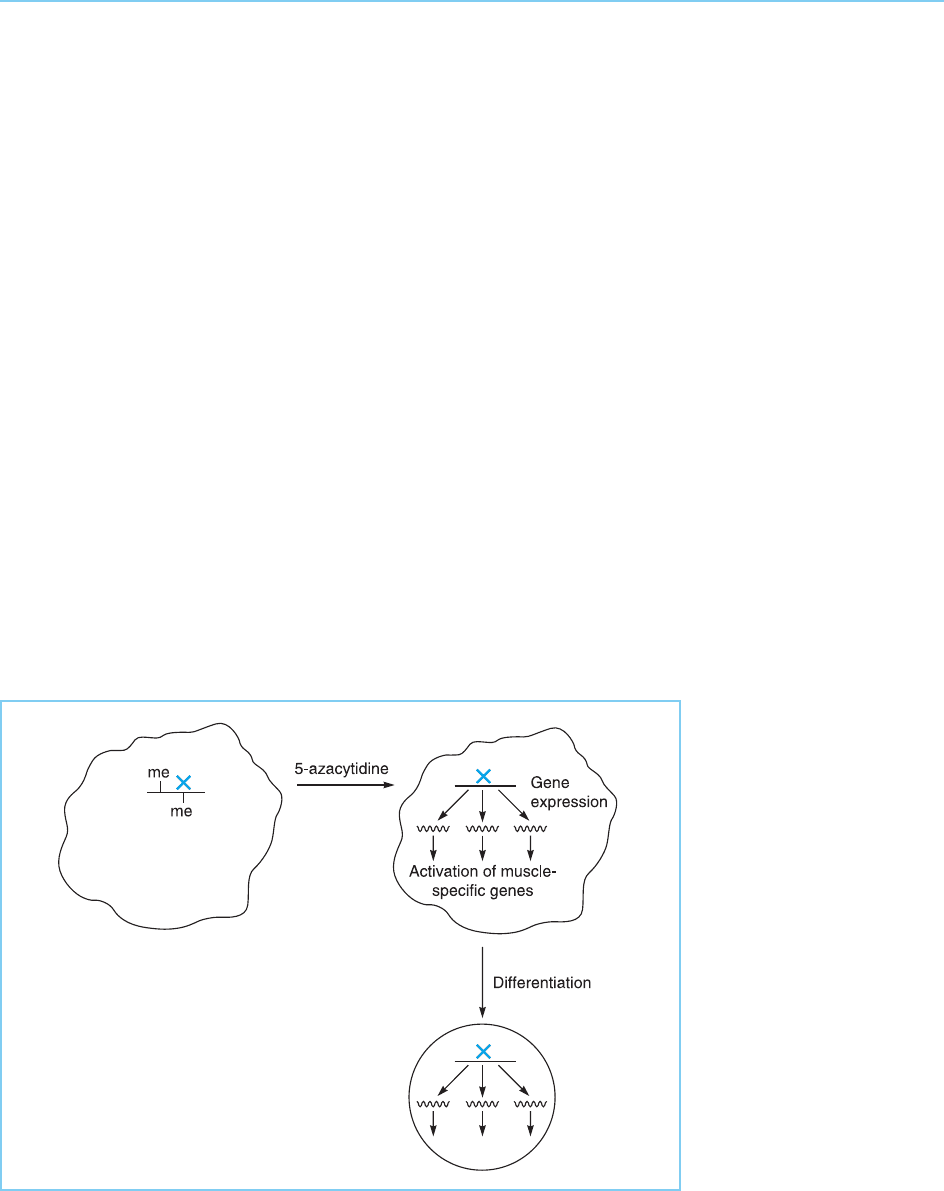
the activation of only one key regulatory locus might be involved. Thus 5-
azacytidine indu ces myoblasts at very high frequency consistent with only the
demethylation of one gene being required while DNA prepared from differ-
entiated cells can also induce differentiation in untreated cells at a frequency
consistent with the transfer of only one activated locus.
Hence differentiation is thought to occur via the activation of one regula-
tory locus (gene X in Fig. 7.2) whose expression in turn switches on the
expression of genes encoding muscle lineage markers which is observed in
the differentiated 10T‰ cells and thereby induces their differentiation. This
suggested that the regulatory locus might encode a transcription factor which
switched on muscle-specific gene expression.
To isolate the gene encoding this factor , Davis et al. (1987) reasoned that it
would continue to be expressed in the myoblast cells but would evidently not
be expressed in the undifferentiated cells. They therefore prepared RNA from
the differentiated cells and remo ved from it by subtractive hybridization all the
RNAs which were also expressed in the undifferentiated cells. After various
further manipula tions to exclude RNAs characteristic of terminal muscle
differentiation such as myosin and others induced non-specifically in all
cells by 5-azacytidine, the enriched probe was used to screen a cDNA library
prepared from differentia ted 10T‰ cells.
This procedure (Fig. 7.3) resulted in the isolation of three clones, MyoA,
MyoD and MyoH whose expression was specifically activated when 10T‰ cells
REGULATION OF TRANSCRIPTION FACTOR SYNTHESIS 213
Figure 7.2
Model for differentiation
of 10T½ cells in
response to 5-
azacytidine. Activation of
a master locus (x) by
demethylation allows its
product to activate the
expression of muscle-
specific genes thereby
producing differentiation.
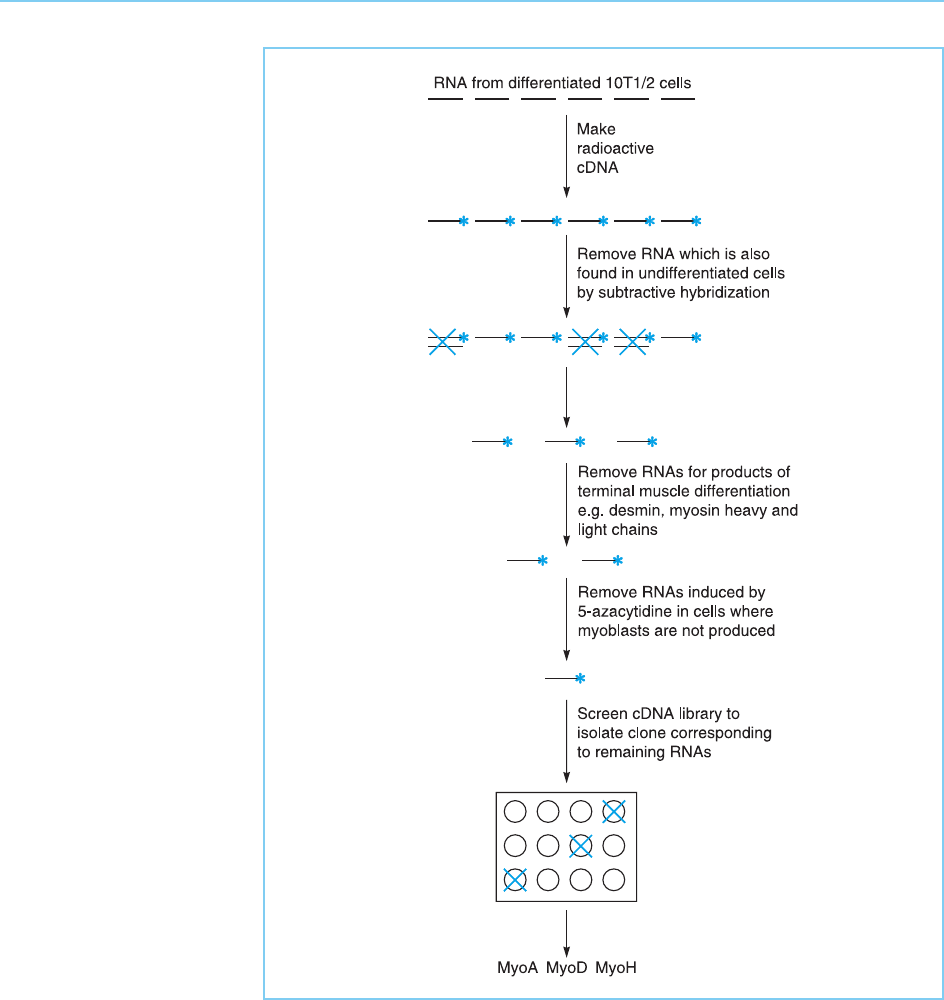
were induced to form myoblasts with 5-azacytidine. When each of these genes
was artificially expressed in 10T‰ cells, MyoA and MyoH had no effect.
However, artificial expression of MyoD was able to convert undifferentiated
10T‰ cells into myoblasts (Fig. 7.4). Hence expression of MyoD alone can
induce differentiation of 10T‰ cells into muscle cells and it is the induction of
214 EUKARYOTIC TRANSCRIPTION FACTORS
Figure 7.3
Strategy for isolating the
master regulatory locus
expressed in 10T½ cells
after but not before
treatment with 5-
azacytidine. Subtractive
hybridization was used to
isolate all RNA molecules
that are present in 10T½
cells only following
treatment with 5-
azacytidine. After removal
of RNAs for terminal
differentiation products of
muscle and RNAs
induced in non-muscle-
producing cells by 5-
azacytidine, the remaining
RNAs were used to
screen a cDNA library.
Three candidates for the
master regulatory locus
MyoA, MyoD and MyoH
were isolated in this way.
MyoD expression by 5-azacytidine that is responsible for the ability of this
compound to induce muscle differentiation.
The differentiated 10T‰ cells produced by artificial expression of MyoD,
like those induced by 5-azacytidine express a variety of muscle lineage markers
and indeed also switch on both MyoA and MyoH as well as the endogenous
MyoD gene itself. This suggests that MyoD is a transcription factor that
switches on genes expressed in muscle cells. In agreement with this, MyoD
was shown to bind to a region of the creatine kinase gene upstream enhancer
which was known to be necessary for its muscle specific gene activity.
Moreover, it has been shown that MyoD can actually bind to its binding sites
within target genes when they are in the tightly packed chromatin structure
characteristic of genes that are inactive in a particular lineage (Gerber et al.,
1997). This binding results in the remodelling of the chromatin to a more open
form and is then followed by enhanced transcription stimulated by MyoD (Fig.
7.5). This alteration in chromatin structure is likely to be dependent on the
ability of MyoD to interact with the p300 co-activator protein (Puri et al., 1997)
(see Chapter 5, section 5.4.3). Like CBP, p300 has histone acetyltransferase
activity and is therefore able to alter chromatin to the more open structure
associated with acetylated histones (see Chapter 1, section 1.2.3).
Hence MyoD is capable of activating transcription by two distinct means,
namely the remodelling of chromatin and the direct stimulation of enhanced
REGULATION OF TRANSCRIPTION FACTOR SYNTHESIS 215
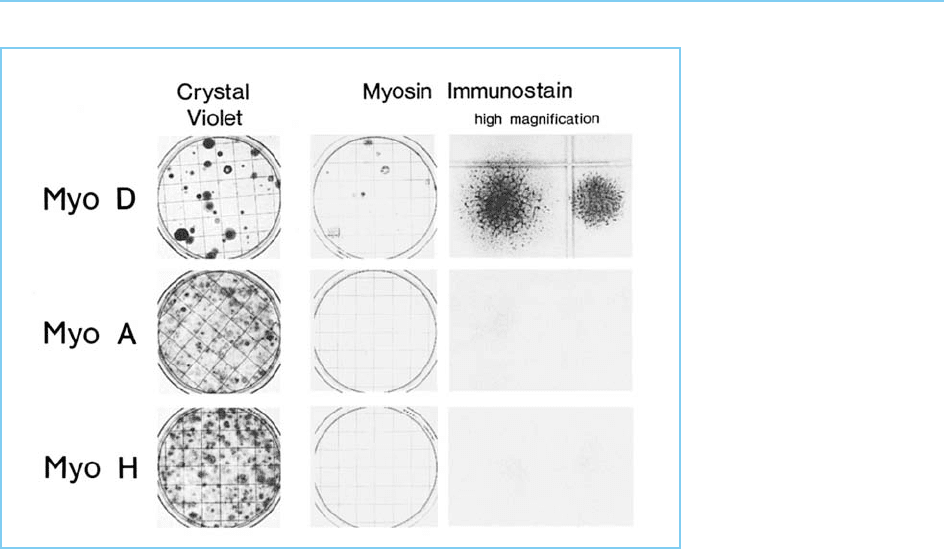
Figure 7.4
Test of each of the
putative master regulatory
loci MyoA, MyoD and
MyoH. Each of the genes
was introduced into
10T½ cells and tested for
the ability to induce the
cells to differentiate into
muscle cells. Note that
while MyoA and MyoH
have no effect,
introduction of MyoD
results in the production
of muscle cells which
contain the muscle
protein myosin. The
differentiated muscle cells
induced by MyoD cease
to divide on differentiation
resulting in less cells
being detectable by
staining with crystal violet
compared to the MyoA
and MyoH treated cells
which continue to
proliferate. Hence only
MyoD has the capacity to
cause 10T½ cells to
differentiate into non-
proliferating muscle cells
producing myosin,
identifying it as a master
regulatory locus for
muscle differentiation.
transcription (see Chapter 5, for a discussion of the mechanisms of tran-
scriptional activation). This is particularly important since it allows enhanced
synthesis of MyoD to induce the development of myogen ic cells from
non-differentiated precursors, in which the genes that must be switched on
are in an inactive closed chromatin structure which is inaccessible to many
transcriptional activators.
Interestingly, as well as stimulating muscle-specific genes, MyoD also pro-
motes differentiation by modulating gene expression so as to inhibit cellular
proliferation, thereby producing the non-dividing phenotype characteristic of
muscle cells. Thus MyoD has been shown to activate the gene encoding the
p21 inhibitor of cyclin-dependent kinases (Halevy et al., 1995). This results in
the inhibition of these kinases whose activity is necessary for cell division (see
Chapter 9, section 9.4.2). In addition, MyoD can also repress the promoter of
the c-fos gene whose protein product is important for cellular proliferation
(see Chapter 9, section 9.3.1) indicating that MyoD can also act by repressing
genes whose products are not required in non-dividing muscle cells (Trouche
et al., 1993).
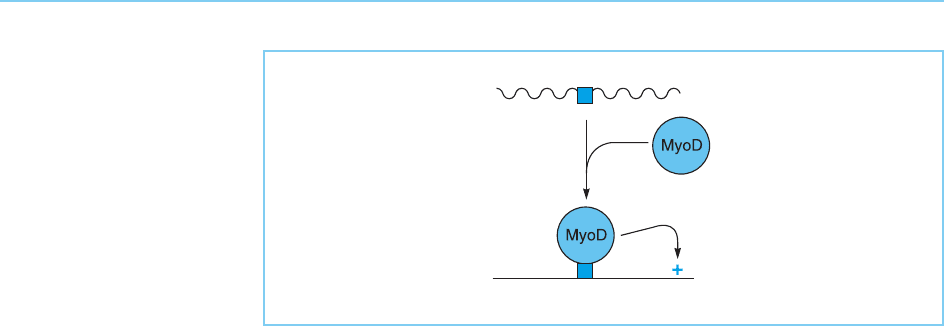
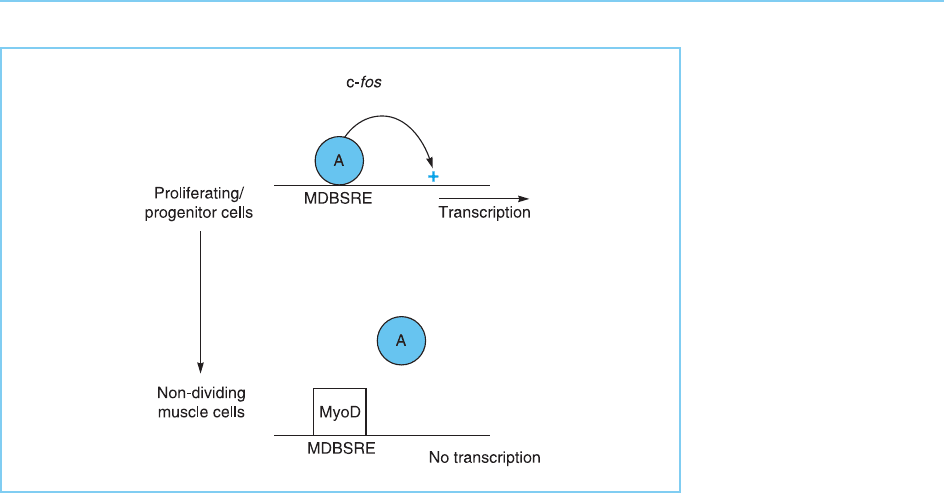
Like gene activation by MyoD, repression of the c-fos promoter is depen-
dent on DNA binding which, in this case, prevents the binding of a pos itively
acting factor to a site known as the serum response element which overlaps
the MyoD binding site in the c- fos promoter (Fig. 7.6). Obviously, in contrast
to its binding to the creatine kinase enhancer, MyoD must bind to its binding
site in the c-fos promoter in a form which cannot activate transcription. Hence,
like the glucocorticoid receptor, MyoD can have different effects on gene
expression depending on the nature of its binding site (see Chapter 6, section
6.2.1 for discussion of the mechanism of transcriptional repression by the
glucocorticoid receptor). In both cases, however, DNA binding by MyoD is
dependent upon a basic region of the protein which binds directly to the DNA
and an adjacent region which can form a helix-loop-helix structure and is
216 EUKARYOTIC TRANSCRIPTION FACTORS
Figure 7.5
MyoD binding to its
binding site (solid box)
both converts the
chromatin structure from
a closed (wavy line) to a
more open (solid line)
structure compatible with
transcription and also
directly enhances the
rate of transcription
(arrow).
essential for dimerization of MyoD (see Chapter 4, section 4.5 for further
discussion of these motifs).
Hence, synthesis of MyoD results in the production of the skeletal muscle
phenotype by activating and repressing the expression of specific target genes.
As expected in view of the critical role that MyoD plays in the development of
muscle cells, the MyoD mRNA is present in skeletal musc le tissue taken from a
variety of different sites in the body but is absent in all other tissues including
cardiac muscle (Davis et al., 1987; Fig. 7.7). The MyoD mRNA and protein
therefore accumulate only in a specific cell type where it is required and the
activation of the MyoD gene during myogenesis is likely to be of central
importance in switching on the expression of muscle-specific genes. In turn,
this suggests that other developmentally regulated transcription factors will be
involved in switching on MyoD expression during myogenesis. In agreement
with this, the paired-type homeobox factor Pax 3 (Chapter 4, section 4.2.7) has
been shown to activate MyoD expression and myogenic differentiation in a
variety of non-muscle cell types (for review see Rawls and Olson, 1997) while
the classical homeobox factor MSX1 can repress the transcription of the
MyoD gene (Woloshin et al., 1995).
Thus, MyoD is a transcription factor whose regulated synthesis results in
the activation of muscle-specific gene expression and the production of ske-
letal muscle cells. Interestingly, the observation that the introduction of MyoD
into cells switches on the endogenous MyoD gene (see above) suggests that a
positive feedback loop normally regulates MyoD expression so that once the
REGULATION OF TRANSCRIPTION FACTOR SYNTHESIS 217
Figure 7.6
MyoD binds to its binding
site (MDBS) in the c-fos
promoter in a
configuration which does
not activate transcription
and prevents binding of
an activating factor (A) to
the overlapping serum
response element (SRE).
This therefore results in
the repression of c-fos
transcription in MyoD-
expressing muscle cells.

gene is initially expressed, expression is maintained producing commitment
to the myogenic lineage (Fig. 7.8). This is of importance since MyoD appears
to be essential for the repair of damaged muscle in adult animals indicating
that its expression must be maintained throughout life (Megeney et al., 1996).
MyoD therefore offers a classic example of the role of transcription factor
synthesis in regulating cell type-specific gene expression. It should be noted,
however, that, as discussed in Chapter 6 (section 6.2.2), the activity of MyoD is
218 EUKARYOTIC TRANSCRIPTION FACTORS
Figure 7.7
Northern blotting
experiment to detect the
mRNAs encoding MyoA,
MyoD and MyoH in
different muscle and non-
muscle tissues. Note that
the MyoD mRNA is
present only in skeletal
muscle as expected in
view of its ability to
produce muscle
differentiation, whereas
the MyoA and MyoH
mRNAs are more widely
distributed. nb, indicates
new born, ad, indicates
adult, rRNA indicates the
ribosomal RNA control
used to show that all
samples contain intact
RNA.
Figure 7.8
Ability of MyoD protein to
activate expression of its
own gene creating a
positive feedback loop
which ensures that,
following an initial stimulus,
the MyoD protein is
continuously produced and
hence maintains myoblast
differentiation.
also regulated by its interaction with the Id inhibitor protein. Hence, MyoD is
regulated both by regulating its synthesis and by regulating its activity.
7.2.2 HOMEOBOX TRANSCRIPTION FACTORS
In addition to its role in controlling cell type-specific gene expression, regula-
tion of transcription factor synthesis is also widely used in the control of
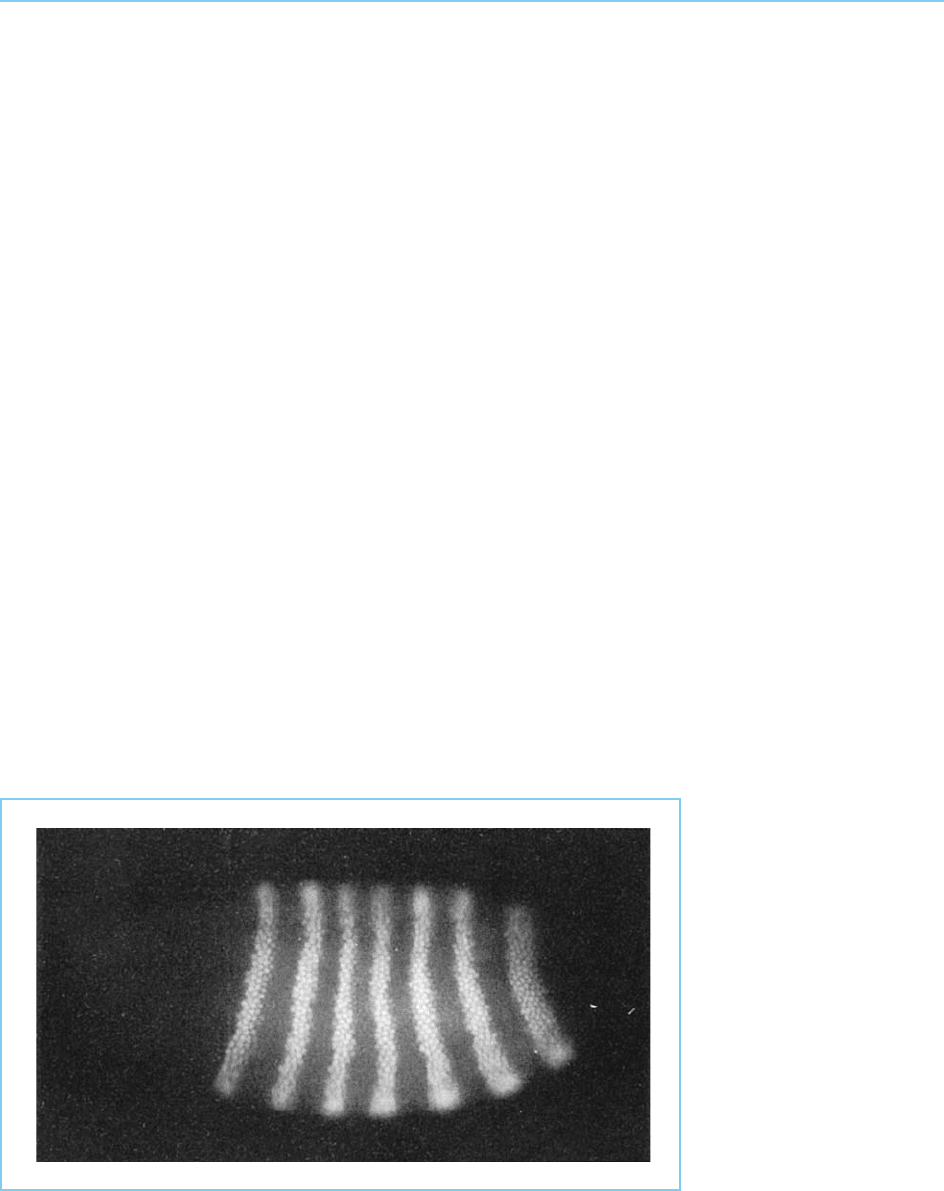
developmentally-regulated gene expression. Thus numerous studies of the
Drosophila homeobox transcription factors discussed in Chapter 4 (section
4.2), using both immunofluorescence wit h specific antibodies and in situ
hybridization, have revealed highly specific expression patterns for individual
factors and the mRNAs which encode them indicating that their role in reg-
ulating gene expression in development is dependent, at least in part, on the
regulation of their synthesis (Fig. 7.9).
Moreover, such regulated synthesis of specific transcription factors can
specifically determine the nature of the cell types that are produced during
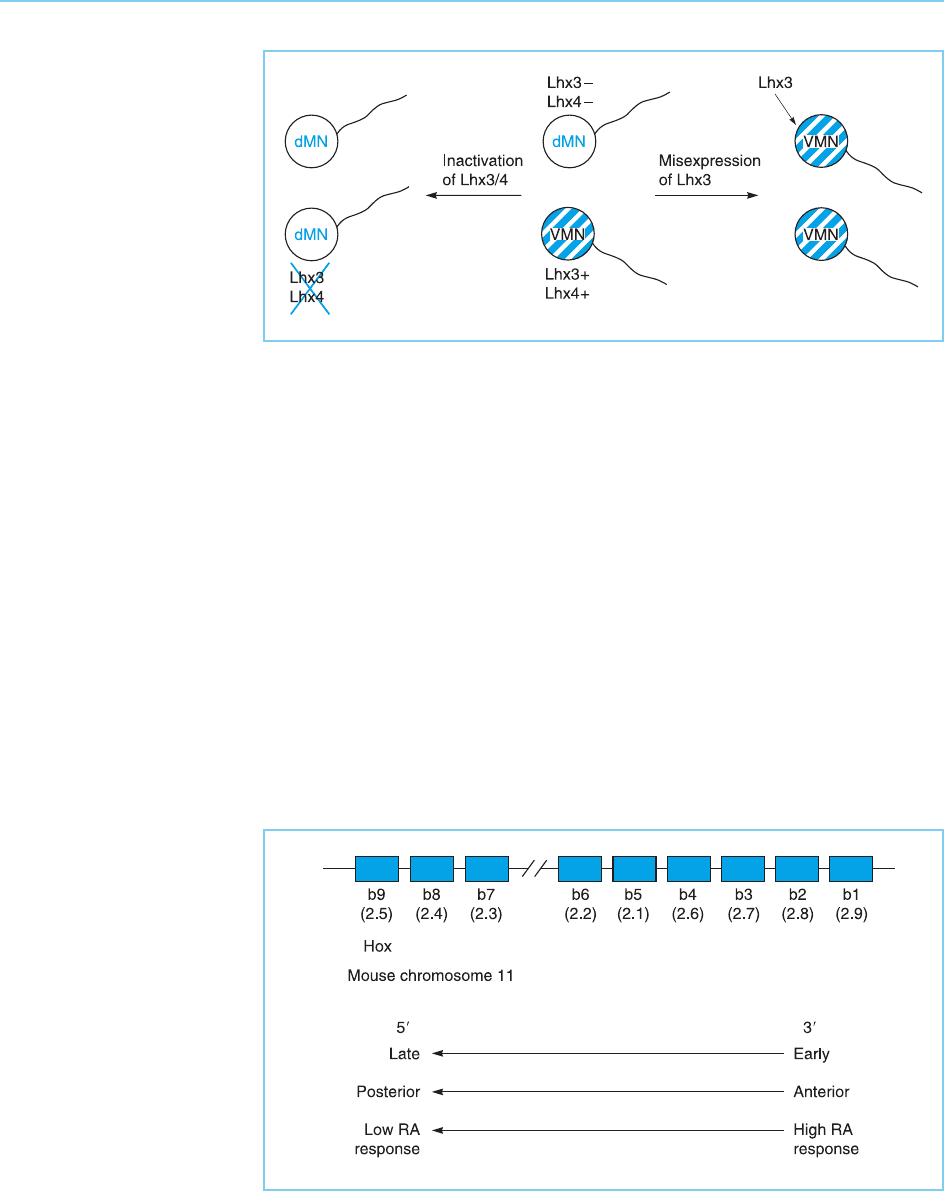
development. Thus, the LIM homeobox factors Lhx3 and Lhx4 are expressed
transiently in motor neurons whose axons extend ventrally (v-MN) but not in
those which extend dorsally (d-MN). In knock out mice lacking both Lhx3 and
Lhx4, cells which should become v-MN cells, instead become d-MN cells (Fig.
7.10). In contrast, misexpression of Lhx3 is sufficient to convert d-MN cells
into v-MN cells (Sharma et al., 1998). Hence, the regulated synthesis of these
two homeodomain proteins results in cells which express them becoming one
type of motor neuro n (v-MN), whereas the cells which do not express them,
form a different type of motor neuron (d-MN).
REGULATION OF TRANSCRIPTION FACTOR SYNTHESIS 219
Figure 7.9
Localization of the Ftz
protein in the Drosophila
blastoderm embryo using
a fluorescent antibody
which reacts specifically
with the protein. The
anterior end of the
embryo is to the left and
the dorsal surface to the
top of the photograph.
Note the precise pattern
of seven stripes of Ftz-
expressing cells around
the embryo.
In an even more dramatic example of this effect, the Pitx2 homeobox
factor is expressed only on the left side of the developing em bryo in the
mouse or chicken. Expressing Pitx2 on the right side of the embryo affects
the normal pattern of asymmetry between the left and right sides of the
embryo (Logan et al., 1998; Piedra et al., 1998), indicating that the appropriate
regulation of its synthesis is required for the embryo to develop distinct left
and right sides.
Hence, the regulated expression of homeobox factors is essential for their
role in regulating gene transcription and cell fate in development. As dis-
cussed in Chapter 4 (section 4.2), the mouse homeobox genes are found in
clusters containing a number of different genes (Fig. 7.11). Most interestingly,
in both Drosophila and mammals, the position of a gene within a cluster is
related to its expression pattern during embryogenesis. Thus, in the mouse
Hoxb cluster all the genes are expressed in the developing central nervous
220 EUKARYOTIC TRANSCRIPTION FACTORS
Figure 7.10
The homeobox
transcription factors Lhx3
and Lhx4 are expressed
in motor neurons whose
axons project ventrally
(vMN) but not in those
which project dorsally
(dMN). Inactivation of
Lhx3 and Lhx4 in knock
out mice converts vMN
cells into dMN cells
whereas artificial
expression of Lhx3 in
dMN cells converts them
into vMN cells.
Figure 7.11
Hoxb gene cluster on
mouse chromosome 11.
Note that in moving from
the left to the right of the
mouse complex, the
genes are expressed
progressively earlier in
development, have a
more anterior boundary of
expression and a greater
responsiveness to retinoic
acid.