Горбунова В.Н., Савельева-Васильева Е.А., Красильников В.В. Молекулярная неврология. Заболевания нервно-мышечной системы
Подождите немного. Документ загружается.

одним из геномных ДНК-зондов, изолированных из области
локализации гена DM (Harley et al., 1992). При проведении с
использованием этого зонда блот гибридизации по Саузерну
в норме обнаруживаются два рестрикционных фрагментам
то время как у большинства пациентов один из этих фраг-
ментов замещен на более крупный. Размеры этого необыч-
ного фрагмента могут варьировать у разных пациентов и даже
в тех случаях, когда больные принадлежат одной семье. На-
блюдается достоверная положительная корреляция между
тяжестью течения, а также более ранним началом заболева-
ния и длиной этого фрагмента. Одновременно методами по-
зиционного клонирования был идентифицирован микросател-
литный CTG-повтор, локализованный в З'-нетранслируемой
области гена DM. Длина этого повтора оказалась значитель-
но больше у больных с миотонической дистрофией (Brook et
al., 1992). Этот повтор при рестрикции геномной ДНК эндо-
нуклеазой EcoRI попадал именно в тот из фрагментов, гиб-
ридизующихся с использованным ДНК-зондом, длина кото-
рого оказывалась большей у больных. Таким образом, была
расшифрована природа необычного рестрикционного EcoRI-
фрагмента.
Изолированный CTG-повтор в популяции также отли-
чается крайней нестабильностью — количество копий в кла-
стере варьирует от 5 до 30—37 При этом для многих популя-
ций характерен двухвершинный характер распределения ал-
лелей. Так, в отечественных популяциях доминируют аллели
с 5 (42.5%) и с 11—13 (37%) повторами (Малышева и др.,
1998). Однако у больных с миотонической дистрофией мини-
мальное количество триплетов значительно больше и состав-
ляет не менее 50 при наиболее мягких формах болезни, от
100 до 1000 у пациентов с классическим течением и с дебю-
том во взрослом возрасте, в то время как при врожденных
формах заболевания количество CTG-триплетов может до-
стигать трех тысяч.
Определение размера CTG-повтора методами ПЦР или
блот-гибридизации по Саузерну позволяет проводить прямую
171
молекулярную диагностику миотонической дистрофии. Око-
ло 90% хромосом с экспансированными CTG-повторами в
отечественных популяциях имеют одинаковый гаплотип по
двум внутригенным полиморфным сайтам рестрикции. Этот
же гаплотип характерен для 91% хромосом с CTG(5) и для
подавляющего большинста хромосом с CTG(15) (Малышева
и др., 1998). Интересно отметить, что транскрибируемый не-
стабильный CTG-(5-30) мотив в З'-нетранслируемой области
гена DM обнаружен только в геноме человека, несмотря на
то что фланкирующие нуклеотиды присутствуют в гомологич-
ном гене мыши.
Недавно в геноме человека был идентифицирован еще
один нестабильный патогенетический транскрибируемый, но
не транслируемый CTG-повтор (Koob et al., 1999). Этот по-
втор расположен в области 13q21, и его экспансия является
причиной развития одной из генетических форм спиноцере-
беллярной атаксии (SCA8). Это неожиданная находка, так как
до сих пор все доминантные спиноцеребеллярные атаксии
были связаны с экспансиями CAG-повторов, кодирующих по-
лиглютаминовые треки, входящие в состав различных бел-
ков. Удлиненные полиглютаминовые треки способны изби-
рательно индуцировать образование устойчивых к протеоли-
зу ядерных агрегатов в определенных типах нейронанальных
клеток, чаще всего в клетках Пуркинье мозжечка. Накопле-
ние подобных ядерных включений оказывает цитотоксичес-
кий эффект и является причиной развития специфических
нейродегенеративных процессов, приводящих к различным
формам спиноцеребеллярных атаксий. Очевидно, что пато-
генетический механизм действия экспансированного нетран-
слируемого CTG-повтора при форме 8 спиноцеребеллярной
атаксии совершенно иной и он, по-видимому, сходен с меха-
низмом действия таких же динамических мутаций при мио-
тонической дистрофии. Для этих двух заболеваний характе-
рен сопоставимый уровень экспансий CTG-повтора и очень
высокая митотическая и мейотическая нестабильность экс-
пансированных повторов, особенно заметная при прохожде-
172

нии мутантного аллеля через женский гаметогенез. Эти осо-
бенности отличают форму 8 от других типов спиноцеребел-
лярных атаксий. Различия в фенотипических проявлениях
экспансий CTG-повторов, приводящих к таким клинически
разным заболеваниям, как миотоническая дистрофия и спи-
ноцеребеллярная атаксия, обусловлены особенностями функ-
ционирования тех генов, работа которых нарушается вслед-
ствие возникновения этих мутаций.
4. Механизмы экспансии CTG-повтора в гене DM
Итак, мутационным механизмом миотонической дистро-
фии является амплификация CTG-повтора в З'-нетранслиру-
емой области гена DM. По-видимому, начальным предраспо-
лагающим к мутационному событию фактором служит пере-
ход от аллеля с пятью CTG-повторами к аллелям, состоящим
из 19—30 триплетов. Популяционная частота этого гетеро-
генного класса аллелей — (CTG)-19—30 — достигает 10% и,
по-видимому, эти аллели составляют резерв для повторных
мутаций. У больных отмечается также черезвычайно высо-
кая соматическая нестабильность CTG-повтора (Ashizawa et
al., 1993), что согласуется сданными о различном характере
экспансии у некоторых однояйцовых близнецов (Fu et al.,
1992). В эмбриональном периоде подобная нестабильность
особенно заметна между 13 и 16 неделями беременности.
Наибольшая экспансия характерна для сердечной мышцы,
при этом степень гетерогенности не коррелирует с началь-
ной величиной экспансированного CTG-повтора (Martorell et
al., 1995). В отличие от мышц, в клетках перифирической кро-
ви больных наблюдается большая нестабильность (CTG)-no-
втора в сторону его экспансии (Martorell et al., 1995). Даль-
нейший анализ нестабильности CTG-повтора, проведенный
на протяжении 1—7 лет на клетках крови 111 пациентов с
варьирующим течением миотонической дистрофии, показал
прямую зависимость уровня гетерогенности от времени и на-
чальной длины повтора (Martorell et al., 1995). Не исключено,
что определенный вклад в формирование соматического мо-
173

заицизма вносят индивидуальные генетические факторы, а
также факторы окружающей среды.
Хорошо прослеживаемой клинической особенностью
болезней экспансии, и в частности миотонической дистро-
фии, является антиципация. Расшифрована молекулярная
природа этого явления. Оказалось, что аллели, связанные с
относительно небольшими экспансиями CTG-повтора, при
которых отсутствуют или наблюдаются лишь слабые клини-
ческие проявления заболевания, обладают высокой мейоти-
ческой нестабильностью. Их прохождение через гаметогенез
сопровождается дальнейшим значительным нарастанием
числа триплетов в нестабильных повторах и в следствие это-
го более резким снижением функции соответствующего гена
(Buxton et al., 1992). Еще одной особенностью наследования
бопезней экспансии является геномный импринтинг — раз-
ное течение заболевания в зависимости от того, получил боль-
ной мутантный аллель от матери или от отца. Так, при миото-
нической дистрофии амплификация CTG-области встречается
в материнском гаметогенезе значительно чаще, чем в отцов-
ском. Это особенно справедливо для тяжелых врожденных
форм заболевания.
В опытах in vitro с использованием геномных клонов,
содержащих локус DM с экспансированными CTG-повтора-
ми, исследовали роль одного из ключевых ферментов репа-
рации — мутазы S2 (продукта гена MSH2), в нестабильности
тринуклеотидных повторов, ответственных за целый ряд тя-
желых нейродегенеративных болезней (Pearson et al., 1997).
Оказалось, что мутаза S2 способна специфическим образом
связываться со структурами, образованными смещением
комплементарных нитей ДНК в области локализации таких
повторов и, таким образом, может принимать участие в их
экспансии.
5. Биохимическая характеристика продукта гена DM
Ген миотонической дистрофии кодирует белок, получив-
ший название миотонин-протеинкиназы — Mt-PK, и поэтому
174
ген DM обозначается иногда как DMPK (dystrophia myotonica
grotein kinase). Этот белок (Mt-PK), состоящий из 624 амино-
кислот, относится к семейству серин/треонин протеинкиназ,
родственных цАМФ-зависимым протеинкиназам (Fu et al.,
1992). Молекулярный вес полноразмерной изоформы Mt-PK,
транслируемой с первого AUG-кодона гена DMPK, составля-
ет 72 кД (Timchenko et al., 1995). Наиболее консервативен
каталитический домен этого белка, содержащий специфиче-
скую киназную и нуклеотид-связывающую конценсусные по-
следовательности. Значительно менее консервативен боль-
шой N-терминальный домен. Промежуточный домен с высо-
ким содержанием альфа-спиралей имеет небольшое сходст-
во с филаментными белками. По-видимому, миотонин-про-
теинкиназа, подобно другим протеинкиназам данного типа,
играет критическую роль в регуляции клеточной дифферен-
цировки и в репликации ДНК, а сам ген DMPK имеет функ-
ции рецессивного супрессора опухолей. Это согласуется с
данными о повышенной частоте у пациентов с миотоничес-
кой дистрофией пиломатриксом (опухолей тканей, производ-
ных нервного гребня), паратиреоидных аденом и небольших
карцином кишечника (Harris et al., 1996).
С использованием набора моноклональных антител на
каталитический и альфа-спиральный домены Mt-PK в скелет-
ных мышцах человека обнаружены три изоформы миотонин-
протеинкиназы — основная с м.в. 55 кД и минорные с м. в.72
и 80 кД (Pham et al., 1998). 5 типов антител на каталитичес-
кий и 5 на альфа-спиральный домены идентифицировали толь-
ко минорные изоформы, причем 72-кД белок присутствовал
во всех исследованных тканях, тогда как 80-кД изоформа
белка с варьирующей экспрессивностью обнаруживалась
главным образом в скелетных мышцах и в интеркалярных
дисках сердечной мышцы. Два типа моноклональных анти-
тел на каталитический домен обнаруживали только мажор-
ную 55-кД изоформу белка, присутствующую исключительно
в скелетных мышцах. Ни один из типов моноклональных ан-
тител не подтвердил ко-локализации Mt-PK с ацетилхолино-
175

выми рецепторами в нейромышечных соединениях. Метода-
ми субклеточного фракционирования и седиментационного
анализа показано, что основная часть Mt-PK в скелетных
мышцах и в мозге имеет цитозольную локализацию.
6. Молекулярные основы патогенеза миотонической
дистрофии
У больных, по крайней мере со взрослой формой забо-
левания, наблюдается уменьшение экспрессии гена DMPK
как на уровне мРНК, так и на белковом уровне (Fu et al.,
1993). В опытах, проведенных на культурах клеток, показа-
но, что в соматических гибридах, содержащих 19-ю хромосо-
му с экспансированным CTG-повтором, снижен синтез пер-
вичного РНК-транскрипта гена DMPK и нарушен его процес-
синг (Carango et al., 1993). Наиболее вероятным представля-
ется, что экспансия CTG-повтора влияет не на транскрип-
цию как таковую, а на посттранскрипционную модификацию
мРНК гена DMPK. Обнаружение повышенных уровней удли-
ненных транскриптов гена DMPK в ядрах клеток пациентов с
миотонической дистрофией позволяет предполагать возмож-
ность нарушения каких-то этапов процессинга мутантного
первичного РНК-транскрипта, таких как полиаденилирование
или транспорт мРНК из ядра в цитоплазму (Wang et al., 1995b;
Taneja et al., 1995). Доминантный характер наследования
миотонической дистрофии, по-видимому, связан с дефици-
том дозы зрелого продукта гена DMPK. Не исключено, что
повышение количества или активности миотонин-протеинки-
назы может оказывать терапевтический эффект на взрос-
лых пациентов. Однако в целом данные по влиянию экспан-
сированного CTG-повтора на экспрессию гена DMPK носят
противоречивый характер.
Более привлекательной кажется другая гипотеза, каса-
ющаяся молекулярных механизмов патогенного действия
динамических мутаций при миотонической дистрофии. По-
казано, что экспансированные CTG-повторы формируют на-
иболее сильные из известных естественных элементов ук-
176
ладки нуклеосом (Wang, Griffith, 1995). При этом эффектив-
ность образования нуклеосом возрастает с увеличением дли-
ны повтора. Можно предположить, что экспансированные
блоки повторов способны менять локальную структуру хро-
мосом, что, в свою очередь, может сопровождаться наруше-
нием экспрессии DMPK и/или других близко расположенных
генов. Это предположение нашло экспериментальное под-
тверждение. Так, недалеко от CTG-повтора была идентифи-
цирована последовательность, гиперчувствительная к дей-
ствию ДНКазы I. При обработке ДНКазой I нормальной ДНК
в этом сайте происходит разрыв. Однако мутантные после-
довательности ДНК с большими экспансированными CTG-
повторами оказываются устойчивы к действию фермента
(Often, Tapscott, 1995). Наиболее простым объяснением дан-
ного феномена является то, что увеличение длины CTG-по-
втора в гене DMPK нарушает локальную структуру хромати-
на, вследствие чего этот гиперчувствительный сайт стано-
вится недоступен для действия ДНКазы I. Предложены и не-
которые другие гипотезы для объяснения механизма повреж-
дающего действия экспансированного CTG-повтора.
В опытах in vitro идентифицирована группа белков, спо-
собных специфически связываться с CTG-повторами, нахо-
дящимися как в однонитевой, так и в двунитевой форме (Yano
Yanagisawa et al., 1995; Richards et al., 1993; Timchenko et al.,
1996). При этом белки, связывающиеся с двунитевыми зон-
дами, имеют, как правило, ядерную локализацию. Кроме того,
обнаружен цитоплазматический белок, специфически взаи-
модействующий с РНКовыми CUG-триплетами — CUG-BP
(Timchenko et al., 1996). Возможно, именно этот белок вовле-
чен в патогенез миотонической дистрофии.
Несмотря на значительные успехи в расшифровке при-
роды мутаций при миотонической дистрофии, остается неяс-
ным, происходит ли дисфункция или приобретение новой
функции гена DMPK на уровне взаимодействия ДНК со струк-
турой хроматина и/или на уровне регуляции процессинга пер-
вичного РНК-транскрипта. По-видимому, в разных тканях
12 Заказ № 170
177
определяющими могут быть различные нарушения, которые в
комплексе и формируют плейотропный фенотип заболевания.
7. Характеристика других генов, участвующих в патогенезе
миотонической дистрофии
Сложность клинического фенотипа миотонической
дистрофии в сочетании с противоречивыми данными, ка-
сающимися действия GTG-экспансии на экспрессию DMPK
локуса, привели к необходимости исследования соседних
генов, которые могут быть вовлечены в этиологию заболе-
вания. Область локализации гена DMPK насыщена актив-
но транскрибирующимися последовательностями. У чело-
века, так же как и у мыши, в непосредственной близости
от гена DMPK в проксимальной области расположен ак-
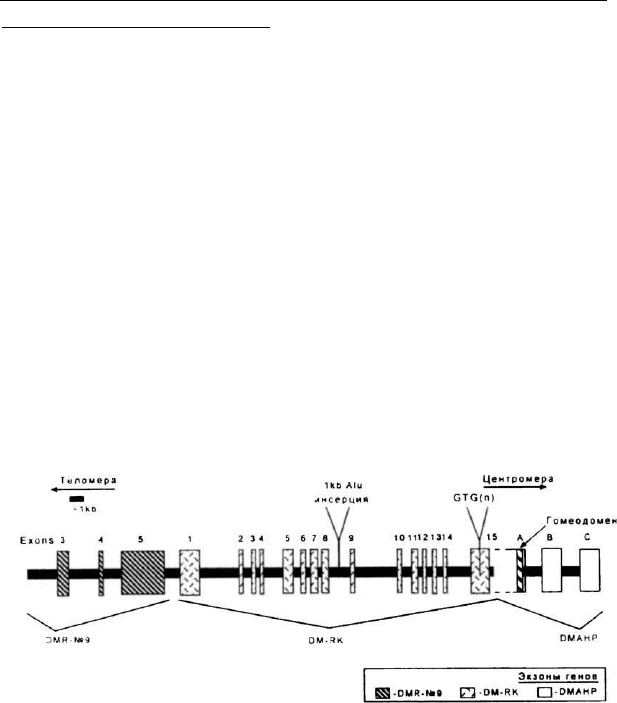
тивный ген — DMR-N9, обозначаемый иногда геном 59 —
рис. 20 (Jansen et al., 1992; Jansen et al., 1995). Этот ген
экспрессируется главным образом в мозге и в testis. Функ-
ция белкового продукта DMR-N9 неизвестна. Высказано
предположение, что экспансия CTG-повтора влияет на аль-
тернативную экспрессию генов DMPK и DMR-N9.
Рис. 20. Структурная организация DM-района
Более вероятным представляется участие в контро-
ле миотонической дистрофии гена DMAHP (dystrophia
myotonica associated homeodomain protein) (или STX5), pac-
178

положенного в непосредственной близости с З'-конца от
DMPK и кодирующего так называемый DM-ассоциирован-
ный гомеодоменный белок. DMAHP экспрессируется во
многих типах тканей, включая скелетные мышцы, фибро-
бласты, лимфоциты, сердце и мозг. Известно, что родст-
венные гомеодоменные белки дрозофилы являются транс-
крипционными факторами, регулирующими экспрессию
генов как на транскрипционном уровне, так и на уровне
трансляции посредством их связывания с ДНК и/или с РНК
соответственно. CTG-повтор локализован внутри CpG-oc-
тровка в З'-нетранслируемой области гена DMPK, и не ис-
ключено, что разрушение этого элемента за счет экспан-
сии повтора может нарушать экспрессию ассоциирован-
ного с этим островком гена(ов). Ранее идентифицирован-
ный гиперчувствительный сайт для ДНКазы I почти цели-
ком расположен в промоторной области этого гена и изме-
нения в этом сайте, вызванные экспансией CTG-повтора,
могут изменять экспрессию DMAHP. Это предположение
было подтверждено при обнаружении в этом сайте энхан-
сера, регулирующего транскрипцию DMAHP (Klesert et al.,
1997). При анализе экспрессии DMAHP в клетках пациентов
с миотонической дистрофией, в которых отсутствует гипер-
чувствительный сайт ДНКазы I, обнаружено 2—4-кратное
снижение устойчивых DMAHP-транскриптов по сравнению
с контролем, причем уровень снижения коррелирует с дли-
ной экспансированного повтора. Не исключено, что нару-
шение работы двух соседних генов DMPK и DMAHP фор-
мирует сложный фенотип миотонической дистрофии.
8. Модельные линии животных
Предположение об участии нескольких соседних ге-
нов в контроле миотонической дистрофии согласуется с
данными по трансгенному разрушению гомологичного гена
Dmpk у мышей. У нулевых мутантов ( Dmpk(-/-)) 72-кД изо-
форма миотонин-протеинкиназы полностью отсутствует. У
таких животных в позднем возрасте развивается прогрес-
179
сирующая миодистрофия с патологическими чертами, сход-
ными с теми, которые характерны для пациентов с мягким
течением миотонической дистрофии (Jansen et al., 1996;
Reddy et al., 1996). У взрослых особей наблюдаются ульт-
раструктурные изменения в мышцах, варьирующая вели-
чина мышечных волокон, с повышенной частотой дегене-
рации и фиброзов. Частота дегенерирующих волокон у
взрослых мутантов повышена на 50% по сравнению с мо-
лодыми животными. У мышей с гиперэкспрессией DM-npo-
теинкиназы, полученных путем трансгеноза нормального
гена DMPK человека в ранние зародыши Dmpk(-/-) мутан-
тов, развивается изолированная гипертрофическая кардио-
миопатия, несмотря на то что 72-кД изоформа миотонин-
протеинкиназы присутствует в большом количестве. Одна-
ко во всех подобных моделях отсутствуют ведущие симп-
томы миотонической дистрофии, включая атрофию мышеч-
ных волокон, миотонию, катаракту и мужское бесплодие.
Эти результаты показывают, что продукт гена DMPK необ-
ходим для поддержания нормальной структуры и функцио-
нирования скелетных мышц. Однако инактивация или из-
менение характера экспрессии DMPK-гена не являются
единственными критическими условиями, необходимыми
для развития заболевания.
В опытах на трансгенных животных показано, что вве-
дение геномных областей, содержащих районы DM локуса
человека с экспансированным CTG-повтором, не приво-
дит к фенотипу, сходному с миотонической дистрофией.
Однако в соматических и зародышевых клетках трансген-
ных мышей наблюдается гипермутабильность введенного
повтора как в сторону экспансий, так и в сторону делеций
(Gourdon et al., 1997; Monckton et al., 1997). При этом не-
стабильность была умеренной при введении 45-кб конст-
рукции, включающей наряду с мутантным геном DMPK с
55 CTG-повторами два фланкирующих гена — DMR-N9 и
DMAHP. Значительно более выраженная мутабильность
наблюдалась в трансгенных линиях мышей, полученных в
180