Ермишин А.П. Биотехнология. Биобезопасность. Биоэтика
Подождите немного. Документ загружается.


ГЛАВА ВТОРАЯ
36
Кардинально решить эту проблему можно путем встраивания трансгена в хромо-
сому бактериальной клетки с помощью методов генетической инженерии. Для этого в
хромосоме бактерии выбирают последовательность, которая может быть повреждена
без ущерба для жизненно важных функций клетки. В нее и встраивают нужный нам
ген со всеми необходимыми для его работы регуляторными элементами. Чтобы осуще-
ствить встраивание трансгена именно в это место, будущий сайт интеграции выделяют
из хромосомной ДНК и клонируют (для интеграции в нужный сайт вводимый ген
должен содержать не менее 50 нуклеотидов хромосомной ДНК). Затем фрагменты ука-
занной ДНК этого сайта переносят в векторную плазмиду, которую конструируют для
переноса нужного нам трансгена таким образом, чтобы она располагалась впереди и
сзади трансгена, как говорят генетики, фланкировала его. Векторная плазмида в дан-
ном случае несколько отличается от тех, которые используются доя клонирования ге-
нов. Она не должна обладать способностью самостоятельно реплицироваться в клетке-
хозяине.
После трансформации бактерии такой рекомбинантной плазмидой, несущей
нужный нам трансген, фланкированный последовательностями ДНК хромосомного
сайта интеграции, происходит спаривание этих последовательностей с гомологичны-
ми последовательностями бактериальной хромосомы. Далее благодаря естественному
процессу генетической рекомбинации (процесс кроссинговера) под действием соответ-
ствующих ферментов клетки-хозяина происходит включение фрагмента ДНК, содер-
жащего трансген, в бактериальную хромосому (рис. 2.7).
Число копий встроенного в хромосому бактерии трансгена можно увеличить с
помощью методов традиционной селекции микроорганизмов. Как показали экспери-
менты, десяток копий трансгена, встроенного в бактериальную хромосому, мог обес-
печивать более высокую выработку протеина — продукта трансгена, чем 20— 40 копий
этого гена в составе многокопийной плазмиды.
2.5.2. Перенос генов в клетки животных. Ученые уже достаточно длительное вре-
мя используют в своих исследованиях культивируемые in vitro дифференцированные
клетки (полученные из разных тканей) человека и животных. Введение в них чужерод-
ной ДНК ненамного сложнее, чем в клетки бактерий. Но поскольку в настоящей книге
речь идет прежде всего о методах создания генно-инженерных организмов, то методы
введения генов в культивируемые клетки животных рассматриваться не будут, так как
получить из них живой трансгенный организм невозможно в принципе.
Первый успех в трансформации животных относится к 1980 году [Gordon et al.,
1980], когда была продемонстрирована возможность введения и интеграции чужерод-
ной ДНК в геном мышей. Важно, что интеграция была стабильной и трансгены сохра-
нялись у потомства, правда, не экспрессировались. Уже через два года были получены
трансгенные мыши с активным чужеродным геном гормона роста [Palmitier et al.,
1982]. В этих первых экспериментах и во многих последующих использовался метод
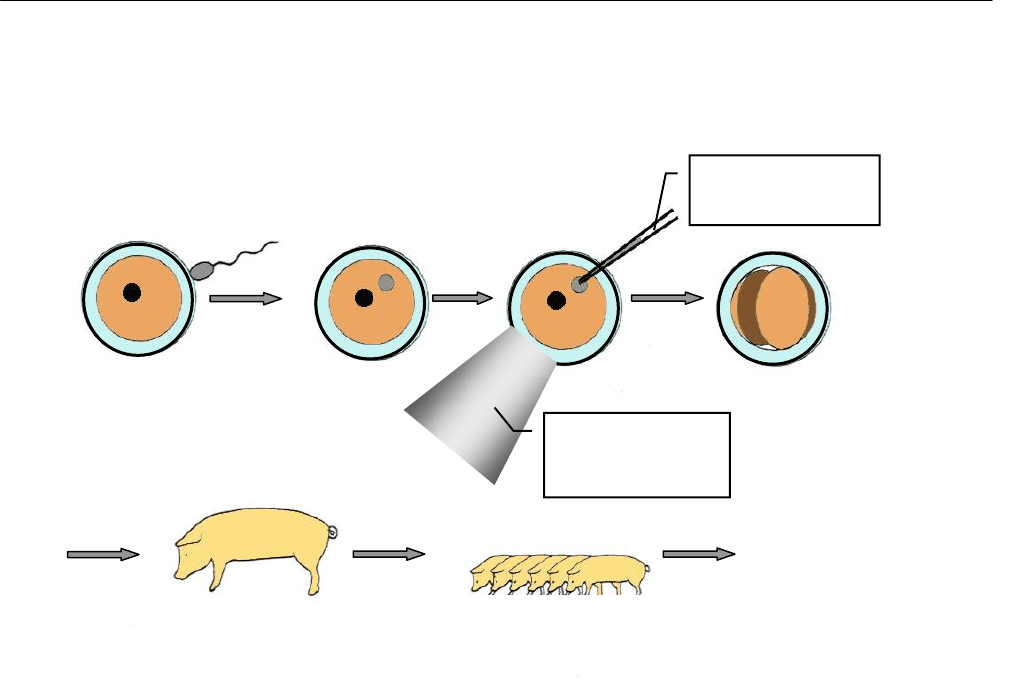
микроинъекции ДНК в пронуклеус (ядро) эмбриона на стадии одной клетки (зиготы).
Метод весьма сложный и деликатный. Он требует наличия сложнейшей микроскопи-
ческой техники и микроманипуляторов. Эмбриональную клетку закрепляют в микро-
капилляре, чей внешний диаметр немного меньше диаметра клетки. Затем с помощью
другого микрокапилляра протыкают оболочку клетки и ядра и впрыскивают в него
трансгенную ДНК (рис. 2.8). Эмбриональную клетку имплантируют в матку суррогат-
ной матери, где происходит развитие эмбриона.
С помощью описанного метода получены генно-инженерные крысы, кролики,
овцы, свиньи, козы, телята и другие млекопитающие. Однако этот метод не только
очень сложный, но и дорогостоящий. Его эффективность для всех перечисленных жи-
КАК ПОЛУЧАЮТ ГЕННО-ИНЖЕНЕРНЫЕ ОРГАНИЗМЫ
37
вотных, особенно жвачных, намного ниже, чем для мышей. Использовать его для мас-
сового производства трансгенных животных экономически невыгодно.
Рис.2.8 Схема трансгеноза животных с помощью микроинъекций ДНК
Микроинъекции чужеродной ДНК в пронуклеусы птиц сильно затруднены из-за
их непрозрачности. То же касается и большинства низших позвоночных, например
рыб. Поэтому для этих видов проводят инъекцию ДНК в цитоплазму эмбриональных
клеток, что резко снижает эффективность метода из-за разрушающего действия эндо-
нуклеаз на введенные гены. Тем не менее, с помощью данной разновидности метода
микроинъекций ДНК удалось получить трансгенные генотипы у кур, нескольких ви-
дов рыб, некоторых морских беспозвоночных.
Несколько лет назад получены первые обнадеживающие результаты в примене-
нии альтернативного подхода в трансформации животных, основанного на использо-
вании не одноклеточных эмбрионов, а половых клеток (гамет). Инкубация изолиро-
ванных сперматозоидов в
растворе чужеродной ДНК с последующим использованием
их для оплодотворения in vivo или in vitro позволила получить генно-инженерные ор-
ганизмы у нескольких видов животных [см. Houdebine, 2001]. Однако результативность
этого метода оказалась очень низкой. Более того, отмечены случаи модификации или
дезактивации вводимых трансгенов под действием эндонуклеаз.
Для повышения эффективности этого метода предлагается использовать обра-
ботку
сперматозоидов детергентами, которые повреждают их мембраны. В результате
ДНК легко проникает в сперматозоиды, что позволяет повысить степень ее включения
в геном. Правда, сперматозоиды после такой обработки теряют способность оплодо-
творять яйцеклетки, поэтому приходится применять их инъекцию в цитоплазму ооци-
тов. В целом подход недостаточно эффективен для многих видов.
2.5.3. Перенос генов
в клетки растений. Характерной особенностью растительных
клеток является наличие вокруг них жесткой целлюлозной оболочки, которую не про-
ткнешь никакими микрокапиллярами. Поэтому для их трансформации используют
специфические методы. И все же с целью обеспечения возможности применения
обычных для микробов и животных клеток методов трансформации разработаны эф-
Микрокапилляр с
раствором ДНК
Микрокапилляр
д
ля удерживания
яйцеклетки
Деление клетки
Перенос трансформированной
яйцеклетки приемной матери
Развитие трансформированной яйце-
клетки и рождение детенышей
Оценка детенышей на
наличие т
р
ансгена

ГЛАВА ВТОРАЯ
38
фективные способы ферментативного растворения целлюлозных оболочек. В резуль-
тате получают так называемые протопласты, способные при определенных воздейст-
виях (химических и электрических) сливаться между собой, а также интегрировать чу-
жеродную ДНК.
Для трансформации растительных протопластов применяют хорошо известные
по бактериям методы стимуляции этого процесса с помощью СаСl
2
, а также полиэти-
ленгликоля. Очень эффективным методом оказалась и электропорация. Весьма пер-
спективной рассматривается методика введения чужеродной ДНК в протопласты с
помощью микроинъекций. Созданы даже компьютерные системы, соединенные с
микроскопом, обеспечивающие достаточно легкое обнаружение, прикрепление и пе-
ремещение отдельных протопластов и микрокапилляров, через которые вводят ДНК в
клетку.
Однако основная
проблема в трансформации протопластов растений — возмож-
ность использовать ее лишь для ограниченного числа видов растений, для которых
разработаны эффективные методы выделения, культивирования протопластов и по-
лучения из них растений-регенератов (подробнее об этом ниже).
Среди методов прямого переноса генов в растительные клетки наибольшее рас-
пространение получил метод биологической баллистики (биолистики), или
, как его
еще называют, бомбардировки частицами (particle bombardment), генного дробовика
(gene gun technique). Суть метода заключается в том, что на мельчайшие частицы (диа-
метром 0,4—1,7 мкм) из вольфрама или золота напыляют рекомбинантные ДНК, со-
держащие предназначенные для переноса гены с необходимыми регуляторными эле-
ментами. Эти частицы разгоняют до огромной скорости (300—600 м/с) в специальном
пневматическом аппарате («генной пушке») и помещают на их пути растительные
ткани (меристемы, незрелые зародыши, колеоптили, протокормы, пыльцу) либо куль-
туры клеток (каллюс или суспензию клеток). В результате частицы проникают в клет-
ки, а чужеродная ДНК встраивается в ДНК хромосом или органелл (рис. 2.9). Несмотря
на ряд недостатков (нежелательная для трансформации растений многокопийность
вставок, разрушение части вносимых генных конструкций и др.), метод биолистики
является основным для получения трансгенных однодольных растений, среди которых
все злаковые культуры, а также многих двудольных растений, являющихся некомпе-
тентными или слабо компетентными для основного метода введения генов в расти-
тельные клетки — агробактериальной трансформации.
Агробактериалъная трансформация. Раскрытие природы агробактериальной
трансформации по праву считается одним из величайших открытий XX века. Оно дало
возможность использовать в генетической инженерии уникальный природный меха-
низм горизонтального переноса генов (т.е. переноса генов между отдаленными в сис-
тематическом отношении видами организмов). Учитывая большое значение агробак-
териальной трансформации при получении генно-инженерных растений, авторы на-
стоящей книги сочли необходимым подробно изложить современные представления о
механизме этого процесса (по Mlynarova, Nap, 1997; Zupan et al., 2000). Это также долж-
но способствовать лучшему пониманию генетических особенностей ГИО, полученных
с помощью данного метода, их отличий от исходных форм.
Онкологическое заболевание растений — корончатый галл — издревле известно
людям (его упоминание имеется еще в трудах Аристотеля). Этой болезни, имеющей
серьезные агрономические последствия из-за нарушения нормального роста растений,
подвержены многие двудольные растения: виноград, косточковые фруктовые деревья,
розы и др.

КАК ПОЛУЧАЮТ ГЕННО-ИНЖЕНЕРНЫЕ ОРГАНИЗМЫ
39
A
B
Рис. 2.9. Метод введения ДНК в клетки растений с помощью биолистики (по Helenius
et al., 2000). А - внешний вид «генной пушки» Helios™ Gene Gun фирмы BioRad. Б -
«обстрел» листьев из «генной пушки». В - результаты «обстрела»: пятна на листьях -
экспрессия репортерного гена gus в трансформированных клетках. Г - регенерация
растений из каллюса, полученного из обработанных листьев.
В начале прошлого века установлен возбудитель заболевания — почвенные бак-
терии из рода Agrobacterium: A.tumefaciens (вызывает у растений образование опухоли —
корончатого галла), A.rhizogenes (вызывает образование на стебле растений многочис-
ленных корней — hairy root), A.rubi (возбудитель галлов у тростника). Однако только в
1970-х годах удалось раскрыть механизм патогенеза. В 1974 году Van Lareke с сотр. и
Zaenen с сотр. установили, что у вирулентных штаммов почвенных бактерий в отличие
от невирулентных имеется большая плазмида, получившая название Ti (tumor inducing
— индуцирующая опухолеобразование) у A. tumefaciens и Ri (root inducing — индуци-
рующая образование корней) — у A.rhizogenes.
В ходе дальнейшего изучения этих плазмид выяснилось, что опухоль возникает в
результате необратимого переноса и включения определенного фрагмента (Т
-ДНК,
т.е. переносимой ДНК) плазмиды в хромосомную ДНК растительной клетки [Chilton et
al., 1977]. Причем гены, расположенные на Т-фрагменте ДНК бактериальной плазми-
ды, активно экспрессируются в растительной клетке, что выражается в синтезе боль-
шого количества специфических соединений опинов — октопинов и нопалинов при
заражении A. tumefaciens или агропинов при заражении A. rhizogenes. Помимо этих со-
единений в клетках образуются в повышенных концентрациях различные фитогормо-
ны, резко стимулирующие ростовые процессы, что приводит к образованию опухоли.
Опины, экскретируемые инфицированными клетками, служат источником азота, уг-
лерода и энергии для возбудителей заболевания — агробактерий, поскольку у них на
Б
Г

ГЛАВА ВТОРАЯ
40
Ti-плазмиде имеются гены, кодирующие ферменты, необходимые для катаболизма
этих соединений. Как видим, агробактерии создали для себя весьма уютную экологи-
ческую нишу, в которой у них практически нет конкурентов.
Процесс агробактериальной трансформации инициируется прежде всего повре-
ждением растения, например, механическим путем при вспашке, прополке и т.п. По-
раненные клетки выделяют в окружающую среду различные химические соединения,
которые привлекают агробактерий (положительный хемотаксис). Бактерии прикреп-
ляются к растениям в местах поранения с помощью целлюлозных фибрилл, что пре-
дохраняет их от смывания, например, при дожде. В генетическом контроле описанных
начальных этапов агробактериальной трансформации в основном задействованы ге-
ны, расположенные на хромосоме бактерии. Их активность определяет все многообра-
зие взаимодействий между бактерией и растением, включая распознавание растения-
хозяина (определенные штаммы агробактерий могут трансформировать определен-
ные виды растений) (рис. 2.10).
Однако в генетическом контроле собственно переноса Т-ДНК главными являются
гены, расположенные в vir-области Ti-плазмиды (vir-гены плазмиды). Правда, некото-
рые хромосомные гены могут влиять на активность плазмидных vir-генов. Она инду-
цируется сигнальными молекулами химических соединений, выделяемых растениями
при поранении: фенольными соединениями типа ацетосирингона и альфа-
гидрооксиацетосирингона (у табака), предшественниками лигнина, синапиновой ки-
слотой, кониферилалкоголем или некоторыми флавоноидами. Для индукции vir-генов
существенным является также наличие таких факторов, как кислая среда, легко ката-
болизируемые источники углерода, например сахароза или моносахариды, темпера-
тура ниже 30°С.
Плазмидные vir-гены расположены на одном регулоне размером около 40 т.п.н.,
который занимает место отдельно от Т-области. У октопиновых штаммов агробактерий
vir-регулон содержит не менее 10 идентифицированных vir-локусов, которые состоят
из нескольких генов, обозначаемых virA, virB и т.д. до virJ. У нопалиновых штаммов аг-
робактерий в аналогичном регулоне обнаружено семь локусов.
Индуцирующий сигнал соединений, выделяемых растениями при поранении,
воспринимается протеинами — продуктами генов virA и virG. Протеин VirA является
трансмембранным и имеет короткий периплазматический связывающий домен (место
прикрепления определенных молекул) — N-терминал, который воспринимает сигна-
лы от растительных фенольных соединений, и длинный цитоплазматический район
— так называемый С-терминал. С-терминал обладает автофосфорилирующей актив-
ностью и способен активировать путем фосфорилирования протеин VirG. Активиро-
ванный VirG является протеином, способным связываться с ДНК. Он распознает соот-
ветствующие регуляторные последовательности других плазмидных vir-генов и спосо-
бен активировать их транскрипцию.
Активация экспрессии vir-генов под действием VirG приводит к запуску ряда по-
следовательных взаимосвязанных событий, которые обеспечивают подготовку Т-ДНК к
переносу в растительную клетку. Первый этап включает сайт-специфическое выреза-
ние кодирующей нити Т-ДНК протеинами — продуктами генов virD. Полипептид
VirD2 в присутствии протеина VirD1 распознает специфические последовательности
Т-ДНК, состоящие из 25 п.н., расположенные в начале и конце Т-области, и производит
вырезание кодирующей нити ДНК. Эти последовательности получили название соот-
ветственно левый (LB) и правый (RB) края Т-области. Они играют исключительно важ-
ную роль в процессе переноса Т-ДНК из бактериальной клетки в растительную.
КАК ПОЛУЧАЮТ ГЕННО-ИНЖЕНЕРНЫЕ ОРГАНИЗМЫ
41
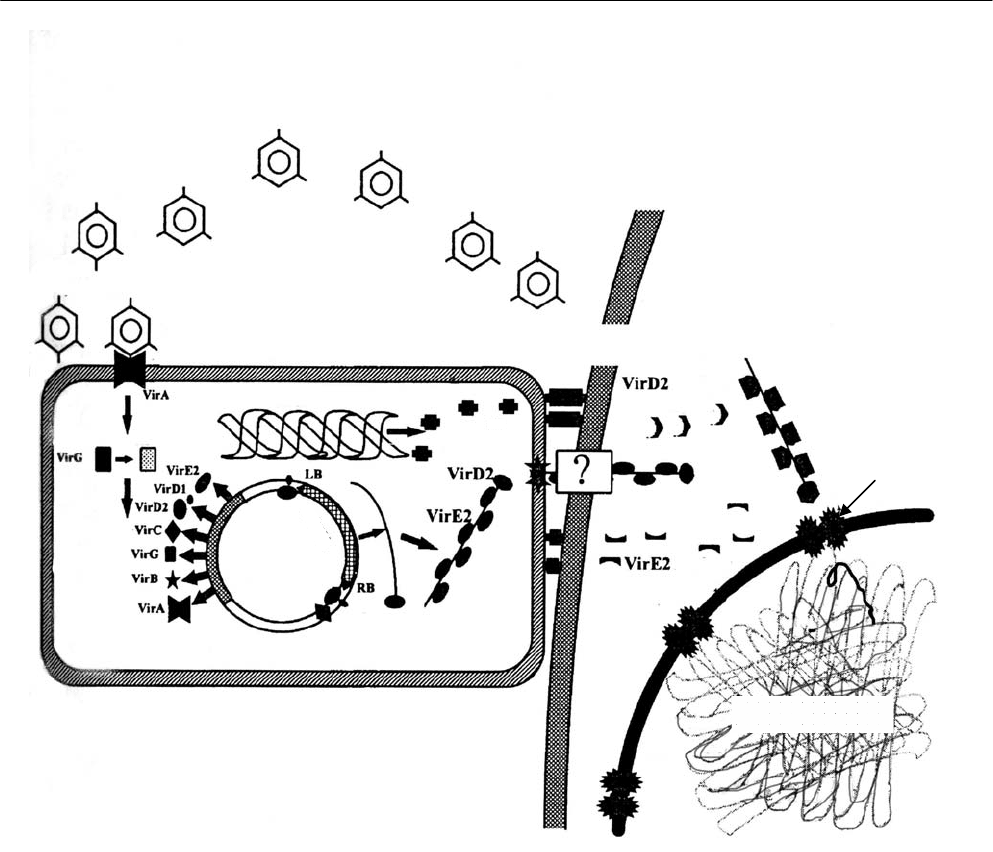
Рис. 2.10. Агробактериальная трансформация (по Mlynarova, Nap, 1997) (пояснения в тексте):
1 – vir-область Ti-плазмиды; 2 – Т-область Ti-плазмиды (Т-ДНК); 3 – Т-нить; 4 – Т-комплекс;
5 – продукты vir-генов бактериальной хромосомы
Вырезанная кодирующая нить Т-ДНК реплицируется, в результате чего образует-
ся однонитчатая ДНК (ssDNA), названная Т-нитью. Точно известно, что именно такая
однонитчатая Т-ДНК переносится в растительную клетку. После вырезания Т-ДНК
VirD2 остается ковалентно связанным с 5’-концом Т-нити и действует как направляю-
щий протеин при движении образовавшегося комплекса
из бактериальной клетки в
растительную, а также предохраняет 5-’конец от действия эндонуклеаз. Этот комплекс
включает также VirE2, который плотно покрывает Т-нить по всей длине, предохраняя
ее от деградации.
Перенос Т-комплекса через мембраны бактериальной и растительной клеток
происходит, как полагают, при участии протеинов — продуктов генов virB. Протеины
VirB формируют так называемый
трансмембранный поровый комплекс, который ус-
танавливает связь между наружной и внутренней мембранами бактериальной клетки
и через который происходит перемещение Т-комплекса. Протеины VirB4 и VirB11 яв-
ляются связанными с мембранами АТФазами, что говорит об активности процесса пе-
реноса Т-комплекса, который осуществляется с затратой энергии. Перемещение Т-
комплекса в цитоплазме растительной клетки к ядру также является активным процес-
сом. Предполагается, что протеины VirD2, а также, возможно, и VirE2 «узнаются» в ци-
топлазме растительной клетки специальными эндогенными протеинами, обеспечи-
вающими доставку Т-комплекса к ядру.
5
бактериальная хромосома
Ti плазми-
да
1
2
34
фенольные соединения
активация
индукция
ядерная пора
встроенная
Т-ДНК
яде
р
ная
Д
НК
ядро
клеточная стенка
цитоплазма
растительная
клетка
Agrobacteriun tumefaciens

ГЛАВА ВТОРАЯ
42
Интеграция Т-ДНК в геном растения завершает процесс ее переноса. Анализ по-
следовательностей ДНК вокруг места вставки показал, что этот процесс не является
сайт-специфичным и не требует высокой степени гомологии. То есть встраивание Т-
ДНК происходит не в строго определенные места (сайты) растительного генома, а в
значительной степени случайно. Тем не менее, отмечено, что «вставки» чаще оказыва-
ются в области транскрипционно активного хроматина, в частности вблизи теломер
хромосом растений. Обычно встраивается одна или несколько копий Т-ДНК в одном
или нескольких местах растительного генома. Нередко встраивание Т-ДНК не проис-
ходит вообще.
Интеграция Т-ДНК в геном растения сопровождается делецией небольшого уча-
стка растительной ДНК (10—100 п.н.). Это говорит о том, что 5’- и З’-концы Т-ДНК
располагаются в процессе встраивания недалеко друг от друга и данный процесс
предполагает участие ферментов эксцизионной репарации. Механизмы встраивания
разных концов Т-ДНК в растительный геном различаются. Вставка 5’-конца обеспечи-
вается протеином VirD2. По-видимому, этот протеин создает благоприятный субстрат
для лигаз клеток растений. Интеграция же 3’-конца в значительной степени аналогич-
на процессу незаконной рекомбинации. После встраивания концов Т-ДНК в одну из
нитей ДНК растения происходит дупликация комплементарной нити Т-ДНК.
В случае если встраивание Т-ДНК прошло без осложнений, гены, расположенные
на ней, стабильно экспрессируются. Так называемые onc-гены (онкогенные) определя-
ют сверхпродукцию фитогормонов индолилуксусной кислоты (ауксина) и цитокини-
нов, что вызывает неконтролируемый рост и деление трансформированных клеток. В
результате образуется корончатый галл. Интересно, что клетки корончатого галла спо-
собны расти в культуре in vitro на питательной среде без фитогормонов, поскольку они
способны обеспечить ими себя самостоятельно. Активность другой группы генов, рас-
положенных на Т-ДНК, связана с образованием опухолеспецифичных метаболитов —
опинов (октопинов, нопалинов, агропинов).
Как видно из представленного описания агробактериальной трансформации, ни
онкогены, ни гены, кодирующие образование опинов, в самом процессе переноса Т-
ДНК из агробактерии в клетку растений активной роли не выполняют. В этом процес
-
се существенную роль играет лишь крайне незначительная часть Т-ДНК, представлен-
ная левым и правым краями из 25 п.н. Если удалить ту часть Т-ДНК, которая располо-
жена между ее краями, и встроить туда нужные нам гены с соответствующими регуля-
торными элементами, то они могут быть перенесены и встроены в
геном растения с
помощью агробактерии точно так же, как она делает это со своими собственными ге-
нами. Именно на этом принципе построено использование агробактериальной транс-
формации в генетической инженерии растений.
Существует два основных типа векторных систем на основе агробактерии. Первая
система основана на применении так называемых cis- или коинтегративных векторов.
Их создают путем вставки нужных нам генов в Т-область Ti-плазмиды Agrobacterium с
помощью гомологичной рекомбинации. Вторая система основа на использовании так
называемых trans- или бинарных векторов. Эти векторы не имеют гомологии с Т-ДНК.
Они обязательно содержат сайт начала репликации (ori) от плазмиды с широким кру-
гом хозяев либо ori-сайты как Agrobacterium, так и E.coli, благодаря чему способны авто-
номно реплицироваться в обоих этих микроорганизмах (рис. 2.11).
При создании современных трансгенных сортов растений в основном используют
бинарные векторные системы. Сам вектор представляет собой плазмиду, которая со-
держит сайт начала репликации (ori), а также по 25 п.н. левого и правого краев Т-ДНК,
между которыми расположены нужные нам гены с соответствующими регуляторными
элементами. Конструируют их исключительно методами технологии рекомбинантных
КАК ПОЛУЧАЮТ ГЕННО-ИНЖЕНЕРНЫЕ ОРГАНИЗМЫ
43
ДНК, описанными выше (встраивание нужных фрагментов ДНК между левым и пра-
вым краями осуществляют с помощью полилинкеров). Методы традиционной селек-
ции микроорганизмов, основанные на гомологичной рекомбинации, практически не
используются. Такие плазмиды можно клонировать в E.coli, где они способны автоном-
но реплицироваться.
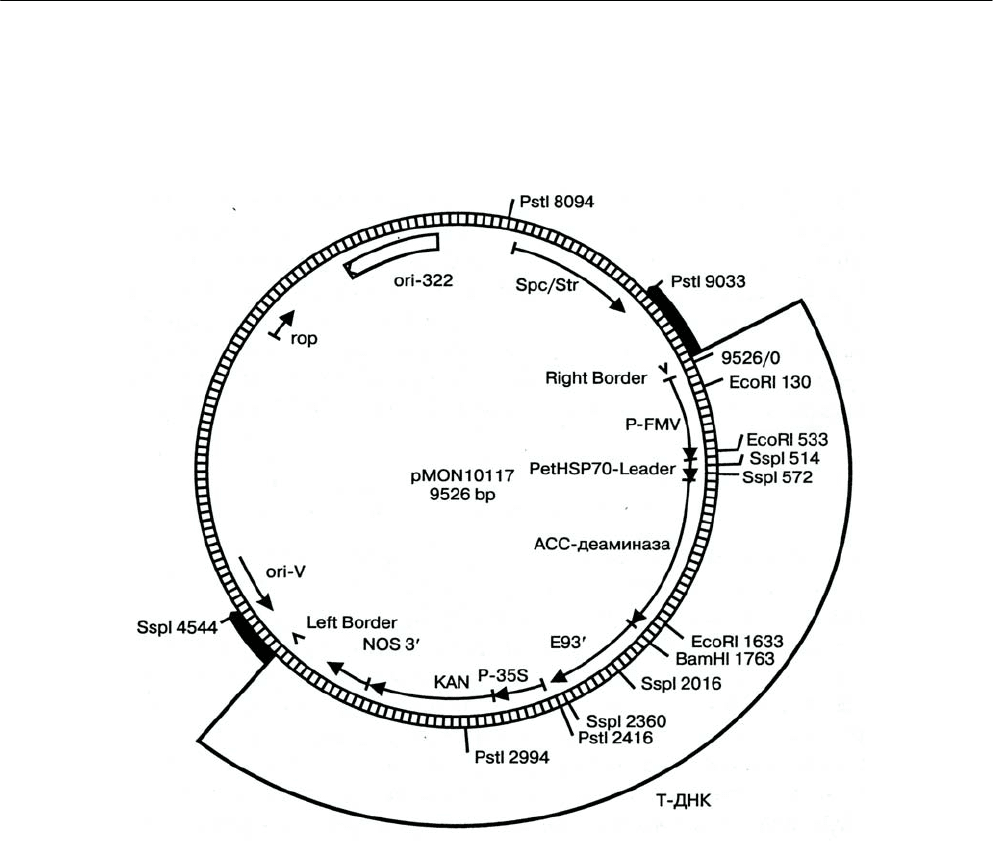
Рис. 2.11. Пример плазмидного бинарного вектора (по APHIS-USDA, 1994). Вектор pMON10117 использо-
ван при создании томатов с удлиненным сроком созревания благодаря пониженному уровню этилена –
фитогормона, регулирующего процесс созревания плодов (встраивание гена, кодирующего фермент
АСС-деаминазу; этот фермент играет важную роль в биосинтезе этилена). По внешнему периметру
плазмиды отмечены сайты рестрикции соответствующих рестриктаз, а также
номера последовательно-
стей нуклеотидов, начиная с правого края (Right border). По внутреннему периметру – последователь-
ность генетических элементов, содержащихся на плазмиде. В ДНК трансгенных растений включена
часть плазмиды, ограниченная левым и правым краями (Left and Right borders), – Т-ДНК. Ori-322 и ori-V-
сайты начала репликации плазмиды соответственно в E.coli и A. tumefaciens
После клонирования, изучения и отбора нужных нам конструкций генов ото-
бранные бинарные векторы переносят в специальные штаммы, созданные на основе
высоковирулентных штаммов Agrobacterium (как правило, A.tumefaciens). Характерной
особенностью этих штаммов является то, что они имеют так называемую разоружен-
ную vir-плазмиду. Последняя представляет собой Ti-плазмиду, которая содержит ин-
тактную
vir-область, но у которой полностью отсутствует Т-область.
Привнесенный в Agrobacterium бинарный вектор способен в ней автономно реп-
лицироваться благодаря наличию ori-сайта от плазмиды с широким кругом хозяев.
Вследствие же наличия разоруженной vir-плазмиды он может успешно переноситься и
встраиваться в геном клеток растений.
Как видим, при разработке бинарных векторных систем использована такая важ-
нейшая особенность механизма агробактериальной трансформации, согласно которой
vir-область Ti-плазмиды лишь обеспечивает перенос Т-области из бактерий в растения,
но сама при этом в растения не попадает. С другой стороны, в процессе переноса Т-

ГЛАВА ВТОРАЯ
44
области существенным является присутствие очень небольшой части Т-ДНК: по 25 п.н.
ее левого и правого краев. Остальная часть Т-ДНК, в том числе все онкогены и гены,
кодирующие образование опинов, для процесса переноса Т-ДНК несущественны. Они
выполняют в нем пассивную роль и могут быть безболезненно заменены любыми дру-
гими генами.
В бинарных векторных системах в отличие от естественных штаммов Agrobacterium
основные «участники» процесса агробактериальной трансформации — vir- и chv-гены,
которые обеспечивают перенос Т-ДНК, и переносимая Т-ДНК (точнее говоря, ее левый
и правый края со вставленными между ними нужными генами) — разделены физиче-
ски. Chv-гены расположены на бактериальной хромосоме, а vir-гены и Т-ДНК — на
разных плазмидах. Это очень удобно для целей генетических манипуляций с бинар-
ным вектором, поскольку существенно увеличивает емкость вектора.
Безусловно, агробактериальная трансформация — наиболее эффективная техно-
логия введения трансгенов в клетки растений. Однако она имеет ограничения, связан-
ные с кругом хозяев агробактерий. Специально отобранные штаммы Agrobacterium с
широким кругом хозяев могут инфицировать приблизительно половину двудольных
видов растений, а также некоторые из голосеменных. Многие двудольные и голосе-
менные и практически все однодольные растения устойчивы к агробактериям и нико-
гда не образуют опухолей. Тем не менее, совершенствование этого метода продолжает-
ся. В результате разработаны протоколы для агробактериальной трансформации риса
[Hiei et al., 1994], кукурузы [Ishida et al., 1996], пшеницы [Cheng et al., 1997]. Выделены
симбиотичные с растениями микроорганизмы, отличные от агробактерий (Rhizobium,
Sinirhizobium, Mesorhizobium), которые после соответствующей генетической модифика-
ции способны к горизонтальному переносу генов [Broothers et al., 2005].
2.6. Экспрессия трансгенов
Как отмечалось выше, каждый ген имеет сложную систему регуляции своей ак-
тивности, в отсутствие которой он просто не будет функционировать. Только для
транскрипции гена (образования мРНК) обязательно наличие, помимо кодирующей
области, также промотора, последовательности, обеспечивающей присоединение к
мРНК поли-А хвоста, и последовательности, указывающей место окончания транс-
крипции. В придачу к этим обязательным элементам генетические конструкции могут
содержать также регуляторные элементы, определяющие, например, место и время ак-
тивности переносимого гена (трансгена), другие гены (с соответствующими генетиче-
скими элементами), например так называемые селективные гены, с помощью которых
выделяют трансформированные клетки реципиентного организма среди преобла-
дающей массы нетрансформированных клеток.
Такие генетические конструкции «собирают» из фрагментов ДНК, которые могут
принадлежать совершенно разным организмам, относящимся к весьма отдаленным
систематическим группам, и даже с участием фрагментов ДНК, синтезированных ис-
кусственно. Например, в плазмиду почвенной бактерии Agrobacterium tumefaciens (A.
tumefaciens) встраивают ген, выделенный из ДНК рыбы (ген холодоустойчивости от
камбалы), промотор у этого гена от вируса мозаики цветной капусты, последователь-
ности присоединения поли-А хвоста и окончания транскрипции (терминальные по-
следовательности) от A. tumefaciens, селективный ген устойчивости к канамицину из
транспозона (подвижный генетический элемент) E.coli, промотор и терминальные по-
следовательности у этого гена те же, что и у гена холодоустойчивости. И вся эта гене-
тическая конструкция предназначена для переноса в растительный организм.
Выбор промотора при создании трансгенных конструкций имеет особое значе-
ние. Существует общая закономерность: прокариотические промоторы могут обеспе-

КАК ПОЛУЧАЮТ ГЕННО-ИНЖЕНЕРНЫЕ ОРГАНИЗМЫ
45
чить активность любого гена, в том числе и эукариотического, только в прокариотиче-
ском организме (у бактерий, синезеленых водорослей). В эукариотическом организме
может функционировать только ген, имеющий эукариотический промотор. Поэтому
при переносе генов от одного вида растений к другому можно использовать гены с их
собственными промоторами. Но если в растение переносится бактериальный ген, то
промотор у него должен быть заменен на растительный.
В последнем случае часто используют промоторы от растительных вирусов, в ча-
стности промотор вируса мозаики цветной капусты CaMV35S. Вирусы по своей приро-
де — существа исключительно активные. Ничто или практически ничто не может их
остановить после того, как они нашли свою жертву (хозяина). Попав в клетку хозяина,
вирусная ДНК интенсивно размножается и, используя «строительный материал и тех-
нику» (рибосомы и молекулы, необходимые для трансляции) клетки хозяина, активно
воспроизводит себе подобных. Это происходит в силу того, что в процессе эволюции
вирусы получили очень сильные промоторы, способные функционировать в любом
генетическом окружении. Для генетической инженерии такое свойство вирусных про-
моторов очень ценно, поскольку обеспечивает активную работу привнесенного в ор-
ганизм трансгена. Чтобы усилить активность трансгена, иногда перед ним помещают
сразу два промотора (генно-инженерные томаты с удлиненным сроком созревания).
Правда, с другой стороны, регулировать активность такого гена очень сложно: он ра-
ботает постоянно и с одинаковой интенсивностью. Во многих случаях это как раз то,
что и требуется.
Если исследователь ставит более сложные задачи в плане регулирования активно-
сти трансгенов, то в его распоряжении имеется в настоящее время достаточно широ-
кий выбор промоторов и других регуляторных элементов, комбинирование которых
позволяет достичь более тонкой регуляции активности трансгенов в пространстве (в
определенных тканях растения) и времени (в определенный период развития). В каче-
стве примеров тканеспецифических промоторов можно назвать промотор PSsuAra из
Arabidopsis thaliana, который обеспечивает экспрессию находящихся под ним трансге-
нов исключительно в зеленых тканях (листьях, стебле); промотор РТА29 из табака
Nicotiana tabacum, который обеспечивает экспрессию находящихся под ним трансгенов
в пыльниках. Промотор rbsSгена малой субъединицы рибулозобифосфаткарбоксила-
зы-оксигеназы A. thaliana — светочувствительный: экспрессия расположенных под ним
трансгенов на свету (например, в листьях) в сто и более раз выше, чем в темноте (на-
пример, в корнях, клубнях).
Для того чтобы сделать возможным эффективное включение протеинов — про-
дуктов трансгенов в метаболизм клетки, трансгенные конструкции могут содержать,
например, такие генетические элементы, которые кодируют образование транзитного
пептида, обеспечивающего доставку трансгенного фермента EPSPS к хлоропластам —
месту синтеза ароматических аминокислот (генно-инженерная соя, устойчивая к гер-
бициду глифосату).
Как указывалось выше, добиться эффективной экспрессии трансгенов у бактерий
намного проще, чем у высших организмов. Трансформация бактерий чаще всего огра-
ничивается включением в их клетки рекомбинантных плазмид, способных самостоя-
тельно в них реплицироваться. Чем больше копий таких плазмид образуется в клетке,
тем больше протеина — продукта привнесенного гена в ней вырабатывается. В случае
с растениями ситуация намного сложнее. Трансгены встраиваются в генетический ма-
териал клетки и их наследование в поколениях ГИО происходит в соответствии с зако-
нами классической генетики. Поэтому важно, чтобы характер наследования трансге-
нов был предельно простым и предсказуемым (желательно, чтобы они наследовались