Caballero B. (ed.) Encyclopaedia of Food Science, Food Technology and Nutrition. Ten-Volume Set
Подождите немного. Документ загружается.


animals, leading to spastic pareses and paralysis of
legs. Later on, the role of thiamin in the central ner-
vous system, especially in pyruvate metabolism of the
brain, could be demonstrated, too.
0030 Neurological signs are also characteristic of the
deficiency state in human beings. The classical mani-
festation of deficiency can be seen in beriberi. This
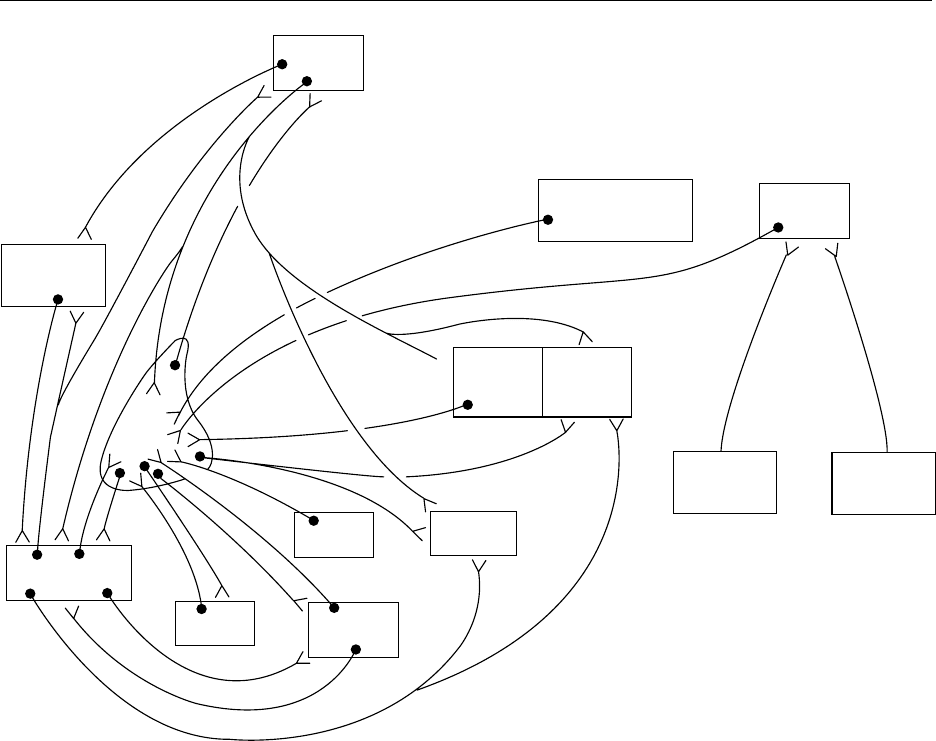
disease is endemic in several regions of East Asia.
Depending on how far the disease has progressed
and additional nutrients are involved (e.g., protein
deficiency), it is characterized by either polyneuro-
pathias and degenerative muscle wasting (dry or
atrophic form) or cardiac arrhythmias and edema
(wet or exudative form). The development of exuda-
tive beriberi predominates in cases of a high carbohy-
drate intake and increased physical activity. Protein
deficiency is probably involved, too. The Wernicke–
Korsakoff syndrome, the cerebral form of deficiency,
is characterized by mental confusion, memory dis-
turbances, ataxia, ophthalmoplegia, and nystagmus.
This syndrome can be interpreted as neuropathia of
the central nervous system and is most commonly
observed in malnourished alcoholics. The thiamin-
dependent PDH and KDH complexes are key
enzymes in the metabolic pathways of glucose and
other energy-delivery substrates. The activities of
both enzymes were found to play a rate-limiting
role in mammalian brain mitochondrial prepar-
ations. Thus, lactic acid accumulation, as a result
of the impaired glucose oxidation and disruption
of oxidative phosphorylation utilizing pyruvate, a-
ketoglutarate, and succinate as substrates has been
demonstrated in brain mitochondria from deficient
rats. There is some evidence that localized lactic
acid accumulation may contribute to the brain lesions
in the Wernicke–Korsakoff disease. Futhermore, it is
suggested that neurological symptoms may involve
an impairment of the cholinergic neurotransmitter
function. Acetylcholine and g-aminobutyrate are pri-
marily produced through the oxidative metabolism
of glucose or via the citric acid cycle with acetyl-
coenzyme A (acetyl-CoA) and a-ketoglutarate as
precursors. Although no change in regional brain
acetylcholine levels was found in thiamin-deficient
rats, acetylcholine utilization and turnover were re-
duced, and the central cholinergic mechanisms
appear to be depressed in a deficient state. These
alterations may be due to a decreased PDH and
thereby activity limited acetyl-CoA production.
0031Infantile beriberi occurring in breast-fed infants
between 2 and 6 months has particularly devastating
consequences. Symptoms include cyanosis, dyspnea,
tbl0004 Table 4 Symptoms of clinical thiamin deficiency
a
Deficiency form Neurological signs Cardiovascular signs Other signs Nutrient interactions
Atrophic beriberi (dry or
polyneuritic form)
Degenerative
polyneuropathia of
lower extremities,
paresthesia, reflex
disorders, dropfoot,
muscle waste,
convulsions
(Caloric restriction),
anorexia
Exudative beriberi (edematous
or cardiovascular form)
Sinus tachycardia,
cardiac arrhythmias,
heart dilatation
(beriberi heart),
pericardial effusion,
cardiac insufficiency
Lung and peripheral
edemas, ascites,
dyspnea, rarely lactic
acidosis without
edemas
(High carbohydrate
intake, protein
deficiency), intense
physical activity
Wernicke–Korsakoff
Syndrome (cerebral form)
Nystagmus,
ophthalmoplegia,
cerebellar ataxia,
paralysis of the sixth
brain nerve,
polyneuropathia
(burning feet
syndrome), reflex
disorders
Psychoses,
hallucinations,
memory loss,
irritability
Alcohol abuse
Infantile beriberi
(2–6 month)
Convulsions (increased
intracranial
pressure)
Tachycardia, cardiac
insufficiency
Vomiting, diarrhea,
abdominal
distension, anorexia,
cyanosis, dyspnea
a
Modified from Bitsch R (1997) Vitamin B
1
(thiamin). In: Biesalski HK, Schrezenmeier J, Weber P and Weiß H (eds) Vitamine, Physiologie, Pathophysiologie,
Therapie, pp. 67–74. Stuttgart: G. Thieme.
5778 THIAMIN/Physiology

tachycardia, abdominal distension, and convulsions.
Sudden death often occurs as a result of cardiac fail-
ure. Nursing mothers, however, often do not show
any signs of clinical deficiency. The thiamin defi-
ciency symptoms are summarized in Table 4.
Therapeutic Administration and Toxicity
0032 The importance of thiamin for the metabolism of the
central and peripheral nerve system, as is becoming
clear from the deficiency symptoms, implicates its
therapeutic application in all neurological system
disorders. The recommended doses are 100–300 mg
per day, often combined with pyridoxine and cobala-
min. Because of the limited gastrointestinal absorp-
tion, the dose should be split up over the day. In
progressive polyneuropathia and clinical beriberi,
the same dosage may be initially administered i.m.
or i.v., for a rapid tissue restoration, followed by oral
therapy.
0033 Lipid-soluble allithiamin derivatives can be given
in smaller doses, because of the improved absorption.
The passage of those derivatives across the blood–
brain barrier is alleviated, too, as has been shown in
animals. A significant improvement in the neuro-
pathia score by oral application of the lipophilic
benfotiamin has been observed in diabetic polyneur-
opathia patients as well.
0034 As with other water-soluble vitamins, thiamin tox-
icity is very small when given orally, because of the
limited absorption. In rare cases, with parenteral
dosages above 100 mg, impaired sensations or even
anaphylactic reactions have been observed, but it is
not clear as to what extent additional components of
the injection solution may interfere.
Thiamin-dependent Enzymopathies
0035 Thiamin-dependent enzymes are involved in four
rare hereditary enzymopathies: leucinosis, Leigh
syndrome, congenital lactacidosis, and thiamin-
dependent megaloblastic anemia.
0036In leucinosis or maple syrup disease, the degrad-
ation of branched chain amino acids is inhibited with
following accumulation of ketoanalogs. In the allevi-
ated or intermittent form, this disorder can be im-
proved by an intake of 10–150 mg of B
1
per day
under simultaneous protein restriction. If untreated,
severe neurological complications, physical, and
mental dysfunctions develop, followed by death.
0037Leigh syndrome is characterized by a necrotizing
encephalomyelopathia and is also observed in rumin-
ants. The exact biochemical defect is poorly under-
stood but is thought to be a hereditary malfunction of
thiamin triphosphate transferase, leading to impaired
TTP formation, occasionally combined with a de-
crease in the amount of ketoacid dehydrogenase.
Neurological symptoms develop, progressing from
nystagmus and paralysis of the external eye muscles
to convulsions, ataxias, and disorientation resem-
bling the Wernicke disease. Thiamin therapy may be
successful, particularly when lipophilic derivatives
(fursultiamin) are ingested up to gram dosages. Lact-
acidosis (which occurs at the same time) can be alle-
viated by oral intake of hydrogencarbonate combined
with a low-carbohydrate diet.
0038The clinical picture of congenital lactacidosis re-
sembles Leigh syndrome, and it is often impossible to
distinguish between the two. The primary cause is a
defect of the PDH or of partial enzymes of this com-
plex. Generally, the prognosis is deleterious. The
lactacidosis and the concomitant neurological symp-
toms could only in rare cases be affected by thera-
peutic thiamin doses.
0039Thiamin-responsive megaloblastic anemia has
until now been very rare. The characteristic symp-
toms are a very strange combination of anemia, laby-
rinthine deafness, and insulin-dependent diabetes.
The anemia can be improved only by oral doses of
tbl0005 Table 5 Congenital thiamin-responsive enzymopathies
Metabolic disorder Biochemicaldefect Clinicalsymptoms Therapy
Leucinosis (maple syrup disease) Ketoacid dehydrogenase for
metabolism of branched-
chain amino acids
Acidosis, ketosis, cerebral
degeneration
Leucin-, isoleucin-, and valin-
poor diet, thiamin (orally)
10–150 mg daily
Leigh syndrome (necrotizing
encephalomyelopathia)
TTP formation, keto acid
dehydrogenase
Lactacidosis, nystagmus,
ataxia, convulsions, mental
and psychic abnormal
development
Thiamin (orally) 2.0 g daily
(lipid-soluble derivatives)
Congenital lactacidoses PDH Lactacidosis, neurological
symptoms, mental
retardation
Thiamin lipid-soluble (?)
Thiamin-responsive
megaloblastic anemia
Thiamin transport in tissue
compartments
Megaloblastic anemia,
diabetes, labyrinthine
deafness
Thiamin (orally) 20–100 mg
daily (lipid-soluble
derivatives)
THIAMIN/Physiology 5779
20–100 mg of thiamin. Folate and vitamin B
12
admin-
istration is ineffective. In one case, the diabetes was
improved, and insulin therapy was no longer needed.
0040 The primary metabolic defect is unclear. Thiamin-
dependent enzymes remain unchanged, and there
may be an impairment in thiamin transport in certain
tissues. The enzymopathies are summarized in Table 5.
See also: Plant Antinutritional Factors: Characteristics;
Beriberi; Bioavailability of Nutrients; Nutritional
Assessment: Biochemical Tests for Vitamins and
Minerals; Functional Tests; Vitamins: Overview
Further Reading
Bitsch R (1997) Vitamin B
1
(Thiamin). In: Biesalski HK,
Schrezenmeier J, Weber P and Weiß H (eds) Vitamine,
Physiologie, Pathophysiologie, Therapie, pp. 67–74.
Stuttgart: G. Thieme.
Duran M and Wadman SK (1985) Thiamine-responsive
inborn errors of metabolism. Journal of Inherited Meta-
bolic Diseases 8(supplement 1): 70–75.
Finglas PM (1994) Thiamin. International Journal for Vita-
min and Nutrition Research 63: 270–274.
Food and Nutrition Board, Institute of Medicine (1998)
Dietary Reference Intakes for Thiamin, Riboflavin,
Niacin, Vitamin B
6
, Folate, Vitamin B
12
, Pantothenic
Acid, Biotin and Choline. Washington DC: National
Academy Press.
Frank T, Bitsch R, Maiwald J and Stein G (2000) High
thiamine diphosphate concentrations in erythrocytes
can be achieved in dialysis patients by oral administra-
tion of benfotiame. European Journal of Clinical
Pharmacology 56: 251–257.
Gregory JF (1997) Bioavailability of thiamin. European
Journal of Clinical Nutrition 51(supplement): 34–37.
Gubler CJ (1991) Thiamin. In: Machlin LJ (ed.) Handbook
of Vitamins, pp. 233–281. New York: Marcel Dekker.
Rindi G (1996) Thiamin. In: Ziegler EE and Filer Jr. LJ (eds)
Present Knowledge in Nutrition, pp. 160–166. Washing-
ton, DC: ILSI Press.
Rindi G, Patrini C, Laforenza U et al. (1994) Further studies
on erythrocyte thiamin transport and phosphorylation
in seven patients with thiamin responsive megaloblastic
anaemia. Journal of Inherited Metabolic Diseases 17:
667–677.
Souci SW, Fachmann W and Kraut H (2000) Food Compos-
ition and Nutrition Tables, 6th revised edn. Boca Raton,
FL: CRC Press/Medpharm Scientific.
Thin-layer Chromatography See Chromatography: Principles; Thin-layer Chromatography; High-
performance Liquid Chromatography; Gas Chromatography; Supercritical Fluid Chromatography; Combined
Chromatography and Mass Spectrometry
THIRST
D J Ramsay, University of Maryland at Baltimore,
Baltimore, MD, USA
This article is reproduced from Encyclopaedia of Food Science,
Food Technology and Nutrition, Copyright 1993, Academic Press.
Introduction
0001 Thirst is a term which evokes many meanings. In most
nonhuman studies, it is equated with measurements of
water intake. In human studies, analog rating scales
are often used to evaluate answers to questions such
as: ’How thirsty are you?’’How dry is your mouth?’
or ’How unpleasant does your throat feel?’ Its use is
not restricted to an appetite for water. Depending on
culture, experience, and availability, a variety of fluids
such as soft drinks and nonalcoholic beverages, in
addition to water itself, may be consumed. It has
been suggested that the definition is best left in
general terms such as ’the disposition to drink’ or
’the tendency to drink,’ and those who wish to use
the word in a more specific sense should define it.
Role of Thirst
0002Water loss from terrestrial animals is continuous.
Ingestion of food results in the addition of osmotic-
ally active solutes to the body fluids either because
they are contained in the foods (e.g., sodium chloride)
or because the process of metabolism produces them
(e.g., oxidation of sulfur-containing amino acids to
sulfate). These solutes are usually excreted in the
5780 THIRST

urine accompanied by water in amounts determined
by the concentrating capacity of the kidney. Thus, if
the kidney can produce urine four times more concen-
trated than plasma – the situation in humans – 25%
of the volume of water will be lost compared to the
situation in which the kidneys could not concentrate
urine over plasma. Nevertheless, even kidneys produ-
cing low volumes of urine at maximal concentrations
result in loss of body fluid. Water is also lost from the
skin by insensible perspiration and sweating. These
amounts are highly variable, depending on the level of
activity, body temperature, and ambient temperature
and humidity. Breathing also results in fluid lost as
expired air is fully saturated with water at body tem-
perature, whereas inspired air is usually drier and at
a temperature lower than that of the body. Small
amounts of water are lost in the feces, although in
diseases accompanied by severe diarrhea, such as
cholera, large amounts of fluid can be lost by this
route. All these mechanisms cause a continuous inev-
itable loss of fluid. Thirst resulting in the ingestion of
an appropriate volume of an appropriate fluid is the
only way to repair the fluid deficit. Without thirst,
animals would die of dehydration. Human subjects
who lose the sensation of thirst as a result of disease
are very difficult to treat. Even the most stringently
prescribed timetable of water intake regime can fail.
Irreversible dehydration is the inevitable outcome of
the lack of thirst. (See Water: Physiology.)
Regulation of Thirst
0003 Water is the most abundant component of the body,
making up approximately 65% of body weight. The
transport of solutes throughout the body takes place
in physical solution, as do most of the physicochem-
ical processes which maintain life. The total osmotic
concentration of body fluids, or osmolality, is main-
tained within very narrow limits. In humans, as in
other animals, it rarely varies by more than 1% or
2%. This is important for two reasons. First, as water
can move freely through cell membranes, the osmol-
ality of intra- and extracellular compartments is
equalized by the movement of water down concen-
tration gradients or osmotic gradients. If plasma and
extracellular fluid osmolality increase, water will be
withdrawn from cells, resulting in cell shrinkage.
Thus, in situations of increased plasma osmolality,
extracellular fluid volume will expand and intracellu-
lar fluid volume will decrease. Acutely, cells behave as
perfect osmometers. It is therefore not surprising that
osmolality is maintained constant in order to main-
tain both blood volume and cell volume within
normal limits. Second, many bio-chemical reactions
and cell transport processes depend critically on the
concentrations of solutes. Thus accurate control of
water balance to insure constancy of the volume and
composition of body fluids is found in all mammals.
Cellular Dehydration Drinking
0004The correlation between total plasma solute concen-
tration or osmolality and drinking has been known
since the work of Mayer in 1900. More recently, the
relationship between plasma osmolality and drinking
has been described by a number of workers in a
number of species, including humans. The relation-
ship is generally investigated by determining the
effects of intravenous infusions of hypertonic sodium
chloride on water intake or, in the case of humans,
increased indications of thirst, as noted on analog
rating scales. In general, there is a linear increase in
thirst associated with increased plasma osmolality
similar to that seen in plasma vasopressin levels, the
major determinant of urinary concentration. Thus
increasing plasma osmolality leads to renal water
conservation and the stimulation of thirst, and
hence water-seeking behavior (Figure 1).
Extracellular Dehydration Drinking
0005Reduction of blood or extracellular fluid volume
without a change in its osmolality also stimulates
thirst. A number of maneuvres have been used to
investigate this phenomenon. Hemorrhage, restric-
tion of venous return by occluding the thoracic part
of the vena cava, or sequestration of fluid using sub-
cutaneous or intraperitoneal injections of colloid all
200
300
100
Water intake (ml)
Plasma vasopressin (pg ml
−1
)
Change in plasma osmolality
− 20
1.0
2.0
3.0
4.0
5.0
0+ 20
fig0001Figure 1 Increase in plasma osmolality stimulates vasopres-
sin secretion (——) and water intake (- - -) in dogs. Reproduced
from Thirst, Encyclopaedia of Food Science, Food Technology and
Nutrition, Macrae R, Robinson RK and Sadler MJ (eds), 1993,
Academic Press.
THIRST 5781

result in stimulation of drinking. In general, drinking
to extracellular fluid dehydration is less sensitive than
that to cellular dehydration. The reduction of ap-
proximately 10% in blood volume is usually required
to stimulate water intake, as is the case with stimula-
tion of vasopressin secretion. With larger reductions
in extracellular fluid volume, however, stimulation of
thirst and vasopressin secretion is marked. Presum-
ably, at the point where a reduction in extracellular
fluid volumes severely compromises circulatory
homeostasis, rapid restoration of blood volume be-
comes essential to insure survival.
Dehydration
0006 During periods of water deprivation, plasma osmol-
ality increases and extracellular fluid volume is re-
duced. It has already been emphasized that, under
normal circumstances, water is lost continually,
whereas drinking is a discontinuous process. Thus
animals are always in a situation of potentially having
to correct for dehydration by taking in fluids. In these
situations, raised plasma osmolality, or cellular dehy-
dration, is a more important input to the stimulation
of thirst than is extracellular fluid dehydration. In
primates, including humans, cellular dehydration
may make up 90% of the stimulus.
Satiety
0007 The stimuli responsible for the cessation of drinking
are different from those which cause it. For example,
dogs deprived of water for 24 h will make up fluid
deficits accurately within 5 min of being allowed
access to water. During this 5-min period, there is
no correction of the raised plasma osmolality or de-
creased extracellular fluid volume as water would not
have been absorbed from the gastrointestinal tract in
sufficient quantities. Indeed, dehydrated dogs with an
open gastric fistula will drink the same amount as
intact dogs. In studies on water-deprived humans,
subjects drank 65% of their total intake within
2.5 min. The remainder of the fluid deficit was con-
sumed over a longer period of time. This satiety effect
has been noted in a large variety of species; it is
associated with a rapid inhibition of vasopressin se-
cretion, and is based on oropharyngeal and gastric
factors. However, it is only a temporary phenomenon
and, unless drinking is followed by absorption of
fluid and correction of the cellular and extracellular
dehydration stimuli, drinking will be reinitiated. (See
Satiety and Appetite: Food, Nutrition, and Appetite.)
Normal Drinking
0008 Not all water intake depends on this simple deficit–
satiety model. The water content of food, and that
provided by its metabolism, can supply significant
amounts of intake. The amount of water consumed
on each occasion can depend heavily on learning and
may be greatly influenced by other behaviors and by
social factors. In humans the situation is even more
complex and fluid may be taken in in association with
caffeine or alcohol, and depend on specific tastes,
such as sweetness, and on flavor. In spite of these
factors, however, it should be noted than on average
the kidney produces urine more concentrated than
plasma. Thus renal-concentrating mechanisms are re-
sponding to a potentially dehydrating situation. In
light of this information it might be asked why
animals are not continually drinking. The answer is
that drinking is only one of a large number of specific
behaviors which may be expressed by an animal; if it
were continually engaged in a search for water, to the
exclusion of other behaviors such as feeding or repro-
duction, the animal would be incapacitated. Presum-
ably, the inputs to thirst become more severe as
dehydration proceeds, and water-seeking behavior
moves to the top of the behavioral hierarchy and is
expressed, resulting in water intake. In the long term,
physiological control mechanisms must underpin
drinking behavior. If inadequate quantities of fluid
are taken in, particularly when urinary concentration
becomes maximal, negative fluid balance and dehy-
dration are the inevitable outcome.
Mechanisms of Thirst
0009Since the pioneering work of Verney on the location
of osmoreceptors controlling vasopressin secretion, it
has been realized that these are located in the hypo-
thalamus. It now seems clear that receptors respond-
ing to physiological changes in extracellular fluid
osmolality are located in circumventricular organs
on the anterior wall of the third cerebral ventricle.
In dogs, lesions of the organum vasculosum laminae
terminalis result in the permanent loss of drinking
behavior to increases in plasma osmolality of up to
15 mosmol kg
1
. In other species, another forebrain
circumventricular organ, the subfornical organ may
be involved. Circumventricular organs lack the
normal blood–brain barrier, and thus can be more
easily influenced by the composition of the blood
that perfuses them. The connectivity between the cir-
cumventricular organs and other parts of the brain
concerned with drinking behavior and secretion of
vasopressin are shown in Figure 2. The nucleus med-
ianus plays a central role in the integration of inputs
which stimulate thirst. Lesions of this region result in
the permanent cessation of drinking to any stimulus.
0010When extracellular fluid volume is reduced,
information may reach the hypothalamus in two
5782 THIRST
main ways. Afferents from atrial and arterial baro-
receptors enter the hindbrain via the ninth and
tenth cranial nerves and synapse in the nucleus
tractus solitarius. From this region, there are a
number of inputs to the hypothalamus, including
the nucleus medianus. Reduction in extracellular
fluid volume also results in the stimulation of the
secretion of renin from the kidneys. Through the
renin–angiotensin cascade mechanism this results in
raised plasma angiotensin II levels. Angiotensin II
generated in the blood interacts with receptors in
forebrain circumventricular organs, particularly
the subfornical organ. Angiotensin II is a potent
dipsogen, particularly when injected directly into
the third cerebral ventricle. Thus extracellular fluid
volume reduction can stimulate thirst directly
through vagal afferents or more indirectly through
the renin–angiotensin system.
0011The physiological inputs to thirst and the connect-
ivity of the hypothalamic nuclei associated with these
inputs are beginning to be understood. However,
there is little information which links these with the
complex neuro-physiological mechanisms associated
with water-seeking and drinking behavior. The mech-
anisms which allow animals to choose drinking from
the vast repertoire of behaviors available to them are
also not clear. There is much to be learned of the
higher-nervous-system functions involved in the link
between a fluid deficit and the stimulation of thirst
and fluid intake.
Requirements for Fluid
0012It is obvious from the foregoing that the requir-
ement for fluid in humans must vary widely. In a
temperate environment, the average loss of water
Parvo
PVN
Magno
PVN
Arterial
baroreceptors
NTS
SFO
SON
PVPO
MPO
LPO
OVLT
Medial
septum
NM
Atrial
baroreceptors
Parabrachial
nucleus
fig0002 Figure 2 Neuroanatomical interconnections between forebrain circumventricular organs and the nucleus medianus in the control of
water balance. SFO, subfornical organ; NM, nucleus medianus; OVLT, organum vasculosum laminae terminalis; MPO, medial preoptic
area; LPO, lateral preoptic area; PVPO, periventricular preoptic area; SON, supraoptic nucleus; PVN, paraventricular nucleus; NTS,
nucleus tractus solitarius. Reproduced from Thirst, Encyclopaedia of Food Science, Food Technology and Nutrition, Macrae R, Robinson
RK and Sadler MJ (eds), 1993, Academic Press.
THIRST 5783
from the skin by evaporation is 900 ml day
1
, in feces
100 ml day
1
and in urine 1500 ml day
1
. The aver-
age figure for the amount of water contained in foods
is 1000 ml day
1
; that produced by metabolism is
300 ml day
1
, leaving 1200 ml to be taken in in the
form of fluid. However, these amounts may vary
widely. For example, during moderate to severe
exercise, particularly in a warm environment, many
liters of fluid may be lost through sweating in order
to maintain normal body temperatures. Under such
circumstances, there is a marked stimulation of
thirst with the resulting intake of large amounts of
fluid. In the final analysis, in order to prevent
dehydration the amount of fluids taken in must
equal that which is lost.
0013 The nature of the fluid ingested depends largely on
cultural factors. For example, consumption of soft
drinks appears to be the most characteristic feature
of the USA. Beer, accompanied by milk (in the
UK) and coffee (in Germany) are typical of middle
Europe, and coffee and milk are the most popular
beverages in northern Europe. Thus, as with food,
culture is the major determinant of drink preference.
It is important also to note the link between food
intake and fluid intake. Under normal circumstances
much of the daily intake of fluid is associated with
eating.
0014 A number of other factors will also influence fluid
intake. During the luteal phase of the menstrual cycle,
there is downward resetting of osmotic thresholds for
both thirst and vasopressin secretion. These changes
may be even more marked during pregnancy, when
plasma osmolality may fall by about 10 mosmol kg
1
during the first few weeks of gestation. During this
period the osmotic thresholds of both thirst and vaso-
pressin are again reduced.
0015There is a reduction of thirst in the elderly. This is
evidenced by the tendency for spontaneous dehydra-
tion to occur in the elderly, and their reduced sensitiv-
ity to infusions of hypertonic saline and water
deprivation. The situation in the elderly is also com-
plicated by their diminished ability to excrete water
loads. For example, if an elderly individual with di-
minished thirst is encouraged to drink more fluid in
warm weather, overcorrection can result in overhy-
dration. Thus in the elderly, there may be problems of
over- or underhydration. (See Elderly: Nutritionally
Related Problems.)
See also: Elderly: Nutritionally Related Problems; Satiety
and Appetite: Food, Nutrition, and Appetite; Water:
Physiology
Further Reading
Denton D (1982) The Hunger for Salt. Berlin: Springer-
Verlag.
Fitzsimons JT (1979) The physiology of thirst and sodium
appetite. Monographs of the Physiological Society 35.
Ramsay DJ and Booth DA (eds) (1991) Thirst: Physio-
logical and Psychological Aspects. Berlin: Springer-
Verlag.
Rolls BJ and Rolls ET (1982) Thirst. Cambridge:
Cambridge University Press.
Rundgren M and Hjelmqvist H (eds) (1989) Hypothalamus
and homeostasis Acta Physiologica Scandinavica
Supplementum 136 (583).
Thyroid Disease See Iodine: Properties and Determination; Physiology; Iodine-deficiency Disorders
Thyroid Hormones See Hormones: Adrenal Hormones; Thyroid Hormones; Gut Hormones; Pancreatic
Hormones; Pituitary Hormones; Steroid Hormones
5784 THIRST

TIN
T Y Toribara, University of Rochester, Rochester, NY,
USA
Copyright 2003, Elsevier Science Ltd. All Rights Reserved.
Background
0001 Tin does not occur freely in nature but occurs mostly
in the form of the dioxide and sometimes as the
sulfide. It is found combined in silicic rocks such as
granite and in small amounts in feldspar. The chemis-
try of tin has been examined in great detail, and much
of it has been published recently in a single book. It is
the intention to cover only those topics that have
some relevance to the occurrence and properties of
tin in food.
Occurrence in Food
0002 The presence of tin in food is primarily the result of
man’s use of the element. The greatest exposure is the
result of the use in cans in which food is preserved.
Rather strong acid solutions are necessary for dissol-
ution of the metal, so usually, very little of the element
found its way into food kept in metallic containers
which were either lined with tin or sealed with
solders, leaving tin as one of the ingredients. (See
Canning: Principles.)
0003 Its metallic form has been used from very early
times as a foil and in alloys such as Babbitt (Sn, Cu,
Sb), bronze (Sn, Cu), pewter (Sn, Pb), solder (Sn, Pb),
and type metal (Sn, Pb, Sb). Contact with these alloys
may allow tin to leach into foods.
0004 Although not in food, exposure to tin chemicals
may occur through their use in dentistry. Tin(ii) fluor-
ide is used as an aid in preventing tooth decay through
its presence in toothpastes, topical solutions, and
mouthwashes.
0005 More recently, a number of organotin compounds
have been used as a stabilizer for the plastic PVC, as a
fungicide in paints, and for wood treatment. In most
uses, there is no direct contact of the organotin com-
pounds with food products, and whatever appears in
the food chain is the result of extraction from the
material containing the organotin compound. These
compounds will be discussed in greater detail in
separate sections.
Properties of Tin Compounds
0006 The compounds of tin may be classified as inorganic
or organic. The properties of the various forms and
the reactions that would cause the element to appear
in the food chain are considered.
Inorganic
0007Tin usually occurs as the metallic element in pure or
alloyed state, or as compounds in the divalent stan-
nous form or the tetravalent stannic form. The use of
the metal to coat cans and other containers of food
has been common for many years. The occurrence of
compounds of inorganic tin most likely to be found in
food result from dissolution of the metal.
0008Initially, tin would be dissolved by an acid and
would be in the divalent form. To keep the metal in
this form, the solution must be quite acidic, and the
element forms complexes of the form [SnX
3
]
, where
X may be a halide or one of several other anions.
Also, to keep the tin in the divalent form, it must be
kept under an inert atmosphere such as carbon diox-
ide. In dilute solution, the Sn(ii) compounds undergo
hydrolysis, giving hydrated tin(ii) oxide, which is not
very soluble. The extent of hydrolysis depends upon
the pH of the solution. In the usual pH range natur-
ally encountered in foods, rather extensive hydrolysis
occurs. In concentrated solution in the presence of
oxygen (air), the divalent form is oxidized to the
tetravalent Sn(iv) state, the ease of oxidation increas-
ing with pH. In very strong alkaline solutions, spon-
taneous disproportionation to the metal Sn and Sn(iv)
can occur. The divalent stannous form is a strong
reducing agent, and it functions as such in many
reactions.
0009Most of the inorganic forms of tin found in solu-
tion or combined with some solid material are in the
tetravalent form. Tin(iv) oxide, the form most abun-
dant in nature, dissolves in aqueous solutions of both
acids and alkalis. The predominant species present in
acid solutions containing complexing anions is
[SnX
6
]
2
, where X may be a halide or one of several
other anions. In strongly alkaline solutions, the main
species is [Sn(OH)
6
]
2
. In dilute solution in the ab-
sence of complexing agents, all aqucous solutions of
Sn(iv) tend to hydrolyze to give a precipitate of
hydrated tin(iv) oxide. The halide SnCl
4
, is a liquid
with a boiling point of 114
C.
Organic
0010Organic derivatives of both divalent and tetravalent
tin are produced by man in significant quantities for
various purposes. The preparations and structures
of many of these compounds have been studied
extensively. The divalent compounds are mostly of
TIN 5785

academic interest and would not be encountered eco-
logically. Tetra-, tri-, and disubstituted tin(iv) com-
pounds have been used extensively for a number of
purposes in recent years. Tetramethyltin is a very
good alkylating agent for the transfer of a single
methyl group. It has been used to convert inorganic
mercury to the monomethylmercury chloride for
measurement of small quantities by gas chromatog-
raphy. (See Mercury: Properties and Determination.)
0011 The property of most interest in the treatment of
samples for speciation is solubility. The dimethyltin
and trimethyltin chlorides and bromides are water-
soluble. Similar ethyl compounds are somewhat less
soluble in water. The tin compounds with larger alkyl
and phenyl groups are very slightly soluble or insol-
uble in water. The tetra-substituted compounds are
all insoluble in water.
0012 When the various compounds enter the environ-
ment, their fate is of importance. At near neutral pH
and normal environmental temperatures, alkyl and
aryl tin compounds are stable in the dark, but break-
down occurs in ultraviolet light. This has been ob-
served with compounds of trimethyltin, tributyltin,
and triphenyltin, in which sequential removal of the
alkyl groups occurs to yield finally inorganic Sn
4þ
.
Other mechanisms of breakdown by soil microorgan-
isms and fungi have been observed.
0013 The only significant formation of tin–carbon bonds
in the environment appears to be via methylation.
Monomethyltin was produced by an inoculum of
mixed microorganisms from Chesapeake Bay sedi-
ments with stannic chloride. An acrobic strain of
Pseudomonas (Ps 244) produced a variety of methyl-
tins from inorganic tin(iv) but very little from tin(ii).
Formation of methyltins by other organisms has also
been reported.
Analytical Methods
Sample Preparation
0014 In general, the analysis of the tin content of a sample
measures the total amount of the element. Most
methods used require a solution of the sample. To
extract the tin from an insoluble substance such as a
tin ore, an initial extraction with dilute nitric acid is
followed by a fusion of the insoluble residue with
sodium peroxide or an alkaline carbonate mixture.
Wet ashing procedures for samples such as tissues or
other biological material can be carried out using
concentrated nitric acid together with sulfuric acid.
Such a procedure will destroy the organic matter, so
that only the total tin content can be measured.
0015 For the successful analysis of an organotin
compound, some knowledge of the chemistry of the
compound is required. The solvent used for the first
extraction depends upon the compound of interest.
Aqueous combinations with tropolone with some or-
ganic solvent are the most common methods. The
method of analysis may be GC (gas–liquid chroma-
tography) or HPLC (high-performance liquid chro-
matography), again depending upon the compound
of interest. These methods will be discussed in a later
section. Sampling and the treatment of the sample are
of utmost importance. Tissue samples are usually
homogenized and then extracted. Complete extrac-
tion is difficult because of a relatively large emulsified
solids layer between the aqueous and organic layers.
One procedure to correct for this is to add a known
quantity of an organotin compound such as a tripro-
pyltin salt known not to be in the sample. The recov-
ery of this compound in the subsequent analysis will
give a correction factor for the compounds being
determined. For a number of compounds, the add-
ition of tropolone to the nonpolar solvent improves
the recovery in the extraction.
Instrumental Methods
0016In most laboratories, emission or absorption atomic
spectroscopy is usually used for measuring the tin
content of a sample. Tin can be analyzed with a
standard emission spectrometer. The newer high-
temperature DC (direct current) arc or ICP (induct-
ively coupled plasma) spectrometers are best.
0017Atomic absorption spectrometry is the most sensi-
tive method available. For the analysis of tin, a ni-
trous oxide–acetylene flame must be used. Modern
instruments with a graphite furnace are best. Here,
only a small sample (c.20ml) is required. Using the
programmed temperature facility, the sample is dried,
then volatilized and decomposed.
0018The detection limits for the atomic spectroscopic
methods are:
0019Emission spectroscopy 30 mgl
1
0020Flame atomic absorption (AA) 100 mgl
1
0021Hydride generation AA 4 mgl
1
0022Graphite furnace AA 0.2 mgl
1
Chemical Methods
0023The early methods for the analysis of tin were gravi-
metric and titrimetric, depending on the quantity to
be measured. These techniques required the separ-
ation of the element prior to the final measurement.
In the gravimetric method, tetravalent tin is precipi-
tated as metastannic acid, as the sulfide, or with a
reagent such as cupferron after first separating it from
interfering ions. The insoluble tin compound is
filtered off, ignited to SnO
2
, and weighed.
5786 TIN

0024 Titrimetric methods involve reducing the tin to
tin(ii) and then oxidizing it to tin(iv) with an iodine
or iodate solution. The stannous form is readily oxi-
dized in the presence of air, and for accurate work, the
reaction is carried out under an atmosphere of CO
2
.
A method using tin(iv) makes use of the oxalatostan-
nate complex, which will incorporate one molecule of
sulfide to form the oxalatothiometastannate com-
plex. The amount of sulfide may be measured by
titration with iodine.
Chromatographic Methods
0025 The early analyses of organotin compounds were by
GC, both conventional and, more recently, with ca-
pillary columns being used. The most common orga-
notin compounds are those that are used as fungicides
in paints or sprays. These include tetra-, tri-, and
dimethyltin compounds, and the homologous ethyl
compounds as well as some phenyl derivatives. GC,
which would seem to be an obvious choice to distin-
guish among the various substituted organotin com-
pounds, must be used with caution. The problem
arises from the fact that elevated temperatures must
be used to volatilize some of the compounds, and at
elevated temperatures, some of the compounds will
undergo transformation. There is a further compli-
cation if a mixture contains both tetramethyltin and a
dimethyltin compound. Tetramethyltin is a good
methylating agent, and the dimethyl compound will
be methylated to a trimethyltin compound. (See
Chromatography: Thin-layer Chromatography.)
0026 In order to be able to operate at lower tempera-
tures, some of the organotin compounds, such as
certain halides, are converted to more volatile species.
Among the more common methods used are the re-
placement of the halide by a methyl or ethyl group, or
the formation of the organotin hydride.
0027 HPLC has become the method of choice for many
analyses, as decomposition of the compounds being
analyzed does not occur in the liquid phase at the
lower temperatures used. For the separation of
methyltin compounds in HPLC, a mobile phase of
60 parts acetone and 40 parts n-pentane is used. For
ethyltin compounds, a mixture of 70 parts acetone
and 30 parts n-pentane is used. As elevated tempera-
tures are not used, the redistribution of species that
occurs during GC is not encountered. (See Chroma-
tography: High-performance Liquid Chromatog-
raphy.)
Effects of Dietary Tin
0028 Most of the tin ingested from canned food has been
inorganic and has been considered low because of
its poor absorption and retention in tissues. Many
studies have been conducted at cellular levels using
Escherichia coli and yeast cells to determine the effect
of tin. It is not certain whether these studies may be
applicable to humans.
0029By using controlled diets fed to weanling rats over
prolonged periods of time, Rader and coworkers have
made an extensive study of the effects of 100–1100 mg
per gram of dietary tin on tissue and bone minerals of
these weanling rats. The level of tin used in these
studies was within the range of values reported for
tin present in unlacquered cans. The total food con-
sumption was restricted to the amount consumed by
the rats fed 1100 mg of tin per gram. The amount of
tin in the food of any rat was constant for a period of
4 weeks Food consumption by rats fed diets contain-
ing 0, 100, and 330 mg of tin per gram was measured
weekly. After 4 weeks, the rats were sacrificed, and
serum was prepared from the blood. Soft tissues were
taken from liver, kidney, and duodenum. Femurs were
also taken for examination. Serum ceruplasmin and
cholesterol were then measured.
0030The results of these studies indicate that anemia
and growth depression are common effects of chronic
ingestion of inorganic tin. Low levels of dietary tin
primarily affect copper and zinc status. Pronounced
copper depletion in all tissues examined was observed
at tin levels as low as 30 mgg
1
. Growth depression
generally occurs when dietary tin levels exceed
500 mgg
1
. Diets high in copper and iron (50 and
250 mgg
1
) reduced the signs of anemia in rats fed
tin at levels up to 1500 mg of tin per gram.
0031In normal animals, about 90% of copper in plasma
is contained in ceruloplasmin. The finding of dietary
decreased ceruloplasmin in tin-treated rats is consist-
ent with the observation of decreased copper in
plasma of rats fed diets containing more than 200 mg
of tin per gram. Copper plays a major role in bone
metabolism through its involvement in collagen
cross-linking. The levels of copper and iron have
been shown to affect anemia in rats.
0032Reduced growth is a common symptom of zinc
deficiency. Low dietary tin adversely affects the
growth of rats fed zinc-adequate diets. These studies
on weanling rats indicate that many effects of inor-
ganic tin have effects that are not directly attributed
to tin, as in the case of reduced growth, even in zinc
adequate diets.
0033The effects of continuous ingestion of small
amounts of tin that reduce the quantity of essential
elements to the levels that slow down growth and
cause anemia have emphasized the importance of
reducing the possible intake of tin. Since the principal
source of tin is in containers used for food including
cans, and items such as cans, trays, and other items
TIN 5787