Boggs S. Principles of Sedimentology and Stratigraphy
Подождите немного. Документ загружается.


Classication
Planktonic
Phytoplankton
Zooplankton
Meroplankton
Pseudoplankton
Benthonic
Sessile benthos
Vagrant benthos
Nektonic
14.5 Distribution of Organisms In Space: Paleobiogeography
497
Description
Example
Organisms at live suspended in
e upper water colu and
which have only a very weak
or limited ability to direct their
own movements
Have the abity to carry on
Diatoms, dinoflagellates
photosynthesis; primary food
coccolithophoridae
producers, or autotrophs
Do not carry on photosynthesis
Foraminifers, radio-laans,
and thus cannot produce their
graptolites
own food (heterotrophs); feed
on phytoplankton
Spend only their juvenile stage as Larvae of most benthonic
plankton; later become organisms such as
free-swimming or bottom-
molluscs
dwelling organisms
Organisms distributed by waves
Mussels, barnacles
and currents as a result of
attachment to floating seaweed,
driftwood, etc.
Bottom-dwelling organisms that
live either on or below the
ocean oor
Benos that attach themselves to Crinoids, oysters,
the substrate (epifauna) brachiopods
Benos that either creep or swim
Starfish, echinoids, crabs,
over e bottom (epifauna) or
clams, worms
burrow into the bottom (infauna)
Organisms able to swim eely and
Mobile cephalopods, fish,
thus move about largely
sharks
independently of waves and
currents
their remains are commonly preserved in the same environment in which they
lived. Because most benthos live in shallow water and have limited ability to
move long distances along the bottom, they tend to be more provincial, and of
somewhat less biostratigraphic significance, an plankton. Nonetheless, ben
thos can be dispersed outside their local environment because many benthon
ic species have a planktonic, juvenile larval stage during which they can be
dispersed by currents. Some workers originally questioned the importance of
larval transport as a mechanism for dispersal of shallow-water benthos over
long distances, such as across ocean basins. It was initially believed that the
duration of the larval stage was so short that the larva would change to the
adult phase while the organisms were still over deep water, causing them to
perish when they settled to e bottom. Subsequent work (e.g., Scheltema,
1977) has demonstrated, however, that there are many shoal-water or conti
nental-shelf benthic invertebrates whose larva have a pelagic stage lasting
from six months to more than one year. Scheltema refers to such larva as
teleplanic, or "far wdering." Su long-lived larval species could cross the

498
Chapter 14 I Biostratigraphy
modern Atlantic Ocean, for example, by way of the main surface currents.
Many workers interpret this ability to become dispersed by pelagic mecha
nisms as being favorable for interpopulation migration and, therefore, gene
flow. Valentine (1977b, p. 145) summarizes the importance of these long-lived
larval forms as follows:
Species with the more long-lived and hardy pelagic larva have the greater
chance to be widely dispersed after reproduction .... Therefore, species
with such atibutes would commonly be able to colonize habitats that lie at
some distance om their parental ranges, and would usually be able to
maintain gene flow to such outly ing populations. Species with shorter
planktonic development periods, smaller broods, or more resicted larval
requirements would tend to colonize only localities that are fairly close to
their parental regions. If a population became established at any consider
able distance om others, gene exchange might be sporadic or lacking alto
gether, leading to divergence between the colonists and the parental popu
lation, and a reduction in their usefulness in correlation.
For a species in a given locality, en, a geographic range exists for
which
colonization is essentially obligatory, as the region lies within the
normal migratory range of the population; us, by some standard time,
occupation is virtually assured. This can be called the local range.
BARRIERS TO DISPERSAL
Each species thus has a potential geographic range that is determined by its
habitat requirements. Few species actually occur throughout their potential
range. Their distribution is restricted owing either to barriers of some type that
prevent their expansion into all areas of suitable habitat or because the species
may not have had time to spread to all suitable areas, especially if barriers are
present. At any given time, there are many regions in the world that could be
colonized by species if they could reach them in appropriate numbers, but
they are barred from reaching them by intervening inhospitable areas. Many
species eventually find ways to broach narrow barriers and perhaps in time
even to cross wider barriers. Once barriers are crossed, or barriers disappear,
the migrant species may find itself in competition for environmental niches
with similar species or similarly adapted species in the new province. In the
face of this competition, either the indigenous species or the migrant species
may become extinct. Alternatively, the less well adapted species could evolve
and become adapted to a different environmental niche. Once a barrier is sur
mounted, the colonizers typically expand their range at the new location until
it is circumscribed by other barriers, filling out their new local range. The in
truding species may subsequently broach still other barriers, hopping from
one habitable region to another across barriers of varying difficulty of penetra
tion and episodically expanding their total species range (Valentine, 1977a).
The broaching of barriers thus leads to expansion of the total range of a
species, although in some cases it may lead to extinction of the species in the
new region or to its evolution to a more adaptable species. On the other hand,
if the opposite situation prevails and a barrier "suddenly" appears and divides
a once-continuous area of suitable habitat, the result is the segregation of the
species into different populations separated by the barrier. The separated pop
ulations would gradually evolve to different species, each with a more re
stricted geographic range than the parent species (Dodd and Stanton, 1981).
Numerous ecological factors can act as barriers to dispersal of organisms,
all of which can be grouped under two major categories: habitat failure, as
when shelf habitats give way to deep-sea condions or to land, and temperature.

14.5 Distribution of Organisms in Space: Paleobiogeography
499
Hallam (1981) suggests that the major controls on faunal provinciality are cli
mate and plate movements. Dodd and Stanton (1981) indicate that the principal
factors controlling geograpc distribution of species are depth and elevation
(that is, water dep and land elevation) and temperatures. These possible con
trols on provinciality are discussed in furer detail in the followg paragraphs.
Temperature
Te mperature is a major barrier to migration of species, and it commonly affects
larvae more an adult organisms. Because the distribution of worldwide tem
peratures is latitudinally controlled, temperature barriers are most important
latitudinally, although seasonal and even diurnal temperature changes are
also important. e boundaries of all modern biotic provinces are in part tem
perature controlled, and ancient biotic provinces were undoubtedly similarly
controlled. Warm-water taxa are restricted primarily to the equatorial zone of
the ocean because no other large parts of the ocean, either at the surface or at
depth, are warm enough to sustain these tropical species. Cold-water taxa, on
the other hand, can extend eir range closer to the equatorial region by migrat
ing down the bathymetric gradient into deeper and colder water, e
phenome
non
of submeence, if they are capable of adapg to greater depths. Also, if a
polar species c manage to find a way of breakg rough the temperature
barrier and crossing e equatorial region, it can find suitable cold-water habi
at or near the surface in the higher latitudes of the other hemisphere.
Some species of organisms are adapted to a wide range of temperatures
and may thus be distributed through a much wider range of temperature
zones than less tolerant species. Nonetheless, even tolerant species are sensi
tive to temperature variations and do not occur toughout all temperature
zones. It must be recognized also that marine temperature zones have changed
throughout geologic time as world climatic zones have shifted response to
plate movements and episodes of glaciation. A given geographic region of the
world may thus record a succession of colder water or warmer water faunas
through time in response to these shifting climatic conditions.
Geographic Barriers
The terms habitat failure, plate movements, and depth-elevation used above
are all different ways of expressing the concept of geographic barriers. ese
geographic barriers arise out of the distribution pattern of landmasses and
oceans and variations in water depths of the oceans. All organisms have limit
ed water depths at which they can survive. Thus, water that is either too deep
or too shallow can constitute a barrier to a particular species of organism.
Landmasses constitute barriers to the dispersal of marine organisms, and the
open ocean is a barrier to migration of land animals and plants from one con
tinent to another. The most important factors influencing geographic barriers
appear to be changes in sea level and changes in the nature and geographic
distribution of landmasses and the ocean oor brought about by plate move
ments (discussed in a following section).
Sea-level Changes
Causes of major cycles of sea-level change are discussed in Chapter 12. Fluctu
ations in sea level cause significant interruptions in biogeographic provinces
because of variations in water depths on the continental shelves. During a
major drop sea level, water is withdrawn om the continental shelves, ex
posing much of the inner shelf. The habitable area of shallow water is greatly

500
Chapter 14 I Biostratigraphy
reduced, leading to crowding and increased competition among shallow
water species that cannot move seaward into deeper water, and to probable
extinction of less adaptable groups. During major rises in sea level, water
depths on the outer continental shelf are increased, and the total area of shal
low water along continental margins is also vastly increased owing to spread
of the seas over the edges of the continents. The available environmental nich
es for shallow-water organisms are correspondgly increased, resulng in
less competition among species for available space and food. These conditions
lead to expansion of the local ranges of species as they move into favorable
habitats, and also probably to rapid emergence of new species (speciation) as a
result of adaptive radiaon of groups at survived the preceding episode of
lowered sea level. On the other hand, as discussed in the preceding section,
transgressions can be accompanied by the onset of low-oxygen conditions
(anoxia) in some environments. Anoxia may have lead to extinction of some
invertebrate animal groups at various times in the past (Fig. 14.1.3).
An inverse relationship appears to exist between the area of continents
covered by sea and phenomenon of endemism. Endemism is the tendency of
species or other taxa to have a very limited geographic range, as contrasted
with pandemism, which is the tendency of species to have worldwide distrib
ution. At times of low sea level and restriction of seas, faunal migrations be
tween continental shelf areas is rendered more difficult, resulting in less gene
flow. Thus, mo local speciation may occur among the dispersible organisms
that
occupy shallower water habitats (Hallam, 1981).
Plate Movements
Te
ctonism is the major factor controlling the distribution of landmasses and
ocean basins. Major changes in the environmental framework of the marine
realm occur as the geographic positions, configurations, and sizes of conti
nents
and ocean basins are changed by global plate tectonic processes. Plate
movements can greatly affect topographic barriers by producing changes in
oceanic widths and depths. As previously discussed, changes in rates of seafloor
spreading may have a major effect on sea level. Plate movements can also alter
latitudinal temperature gradients by shifting the geographic position of conti
nents, and they can even affect the distribution pattes of major ocean cur
rents. The creation or destruction of migration barriers may thus be tied
closely to plate tectonics events.
Other Barriers
Other barriers, less important than temperature and geographic barriers, may
also help to define the boundaries of biogeographic provinces. Salinity dier
ences constitute an important boundary between freshwater and marine
provinces;
however, salinity is a relatively unimportant barrier within the ma
re realm itself. Salinity can markedly increase in some small, restricted arms
of the ocean where evaporation rates are high. Conversely, lower than normal
salinities may ensue in some coastal areas where freshwater runoff is high.
These salinity variations can conol local communities of organisms but not
the distribution of organisms on a provincial level. In the open ocean, salinity
tends to be highest in the equatorial gion, where evaporation rates are at a
maximum, and lowest in the middle latitudes, where some dilution occurs as
a result of freshwater runoff from the continents. Even so, e salinity in these
regions varies only a few parts per ousand from the average ocean salinity
(35 %o ), a variation not adequate to seriously affect the dispersal of organisms
in the open ocean.

14.6 combined Effects of the Distribution of Organisms in Time and Space
501
Currents aid in the dispersal of planktonic species and the larva of ben
thonic species, but they help also, in some parts of the oce, to maintain the
temperature gradients that create barriers to dispersal. Thus, currents may act
as
either a barrier or an aid to dispersal. The long-term pattern of currents is it
self affected by plate movements.
14.6 COMBINED EFFECTS OF THE DISTRIBUTION
OF ORGANISMS IN TIME AND SPACE
Both the environmental and the temporal (variations with time) records are impor
tant for interetation of geologic history. If organisms throughout geologic time
had been spread over the world and not confined to specific biogeographic
provinces and environments, worldwide correlation of strata on the basis of fossils
would be greatly facilitated, assuming that evolutionary changes have been simul
taneous and worldwide. Under these conditions, however, fossils would provide
little or no help working out ancient depositional environments because more or
less the same organisms would have lived all environments. Conversely, if or
ganisms were disibuted in biogeographic provinces as they are today, but organic
evolution never occurred, we would be able to terpret local ancit environments
with great confidence because ancient sedimentary rocks would contain the same
species as mode environments. By the same token, these species would be of no
value in corlation and the unraveling of local chronologies because the same
species would have existed throughout geologic time.
The real fossil record reflects the fact that both segregation into biogeograph
ic provinces d organic evolution took place. Owing to organic evolution, we are
able to correlate strata of a given age from one area to another and to work out the
relative chronology of strata in a given area. Because many organisms were con
fined to biogeographic provinces in the past, however, we cannot always correlate
time-equivalent strata from different environments because the organisms that ex
isted in different biogeographic provinces during the same period of time were
different. Thus, coelation between biogeographic provinces is difficult, and is
commonly not possible to make worldwide correlations. On the other hand, be
cause dierent groups of organisms were confined to different provinces and dif
ferent environments, e provinciality of ancient organisms provides an invaluable
tool for interpretg ancient sedimentary environments.
The provinciality of organisms creates special problems from the stand
pot of determining the total vertical stratigraphic range of a species. A species
may exist in one province for long periods of time before broaching a particular
barrier and spreading into a nearby province. After migration into the new
province, the species may die out in the old province while continuing to thrive
for some time in the new region. Therefore, the local vertical range of a species
in a given province, sometimes called the teil zone, may be much shorter than
the total range of the species on a global scale. Paleontologists must be extreme
ly careful about recognizing this possibility when using fossils for time correla
tions. This problem is demonstrated in Figure 14.7, which illustrates some of the
factors that can affect the range of a species. This diagram shows that the range
of a species is aected both by evolutionary changes and by the presence of bar
riers that can regulate the times of migration into and first appearance in nearby
provinces.
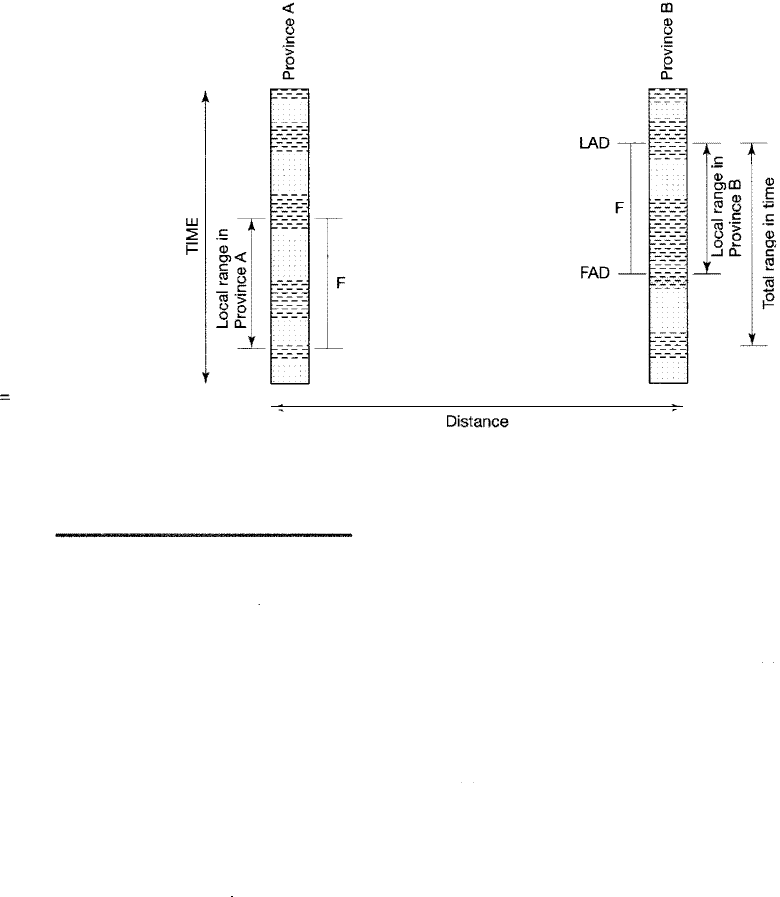
502
Chapter 14 I Biostratigraphy
Figure 14.7
LAD
Diagram illustrating the difference in local
range and total range of a hypothetical
species (F). Species F first appears in
Province A and is restricted to Province A
by a barrier. Later removal of the barrier al
lows migration to Province B, where the
species persists for a time after it has died
out in Province A. FAD first appearance
datum; LAD
=
last appearance datum.
FAD
Ba
rr
ie
r
remov
:}
I
I
I
.
� Bae
r
14.7 BIOCORRELATION
Biostratigraphic units are observable, objective stratigraphic units identified on
the basis of their fossil content. As such, they can be traced and matched from one
locality to another just as lithostratigraphic units are traced. Biostratigraphic units
may or may not have time significance. For example, assemblage biozones and
abundance biozones may cross time lines (be diachronous) when traced laterally.
On the other hand, taxon-range biozones and interval biozones, particularly those
defined by first appearances of taxa, yield correlation lines that coincide in gener
al with time lines. Biostratigraphic units may be coelated, irrespective of their
time significance, using much the same prciples employed in corlation of
lithostratigraphic units-matching by identity and position in the stratigraphic
sequence, for example. this section, we will first examine correlation by assem
blage biozones and abundance biozones, wch can be correlated as biostrati
graphic units even though they may not have time-stratigraphic significance. We
will then discuss biocorlaon methods based on terval zones and other zones
that yield time-stratigraphic correlations.
The following discussion is aed primarily at biocorrelation on the basis of
marine invertebrate organisms. Readers should keep in md, however, that stra
ta can be zoned on the basis of terrestrial animal and plant remains and that corre
lation can in some cases be made on the basis of such data (e.g., Flynn and
Swisher, 1995).
Correlation by Assemblage Biozones
Assemblage biozones are based on distinctive groupings of three or more taxa
without regard to their range limits (Fig. 14.3). They are defined by dierent suc
cessions of faunas or floras, and they succeed each other in a satigraphic secon
without gaps or overlaps. Assemblage zones have particular significance as an in
dicator of environment, which may vary gatly regionally. erefore, they tend to
be of greatest value in local correlations. Nonetheless, some assemblage zones
based on marine planktonic assemblages may be used for correlation over much
wider areas. The principle of correlation by assemblage zones is illustrated graph
ically in the very simple example shown in Figure 14.8.
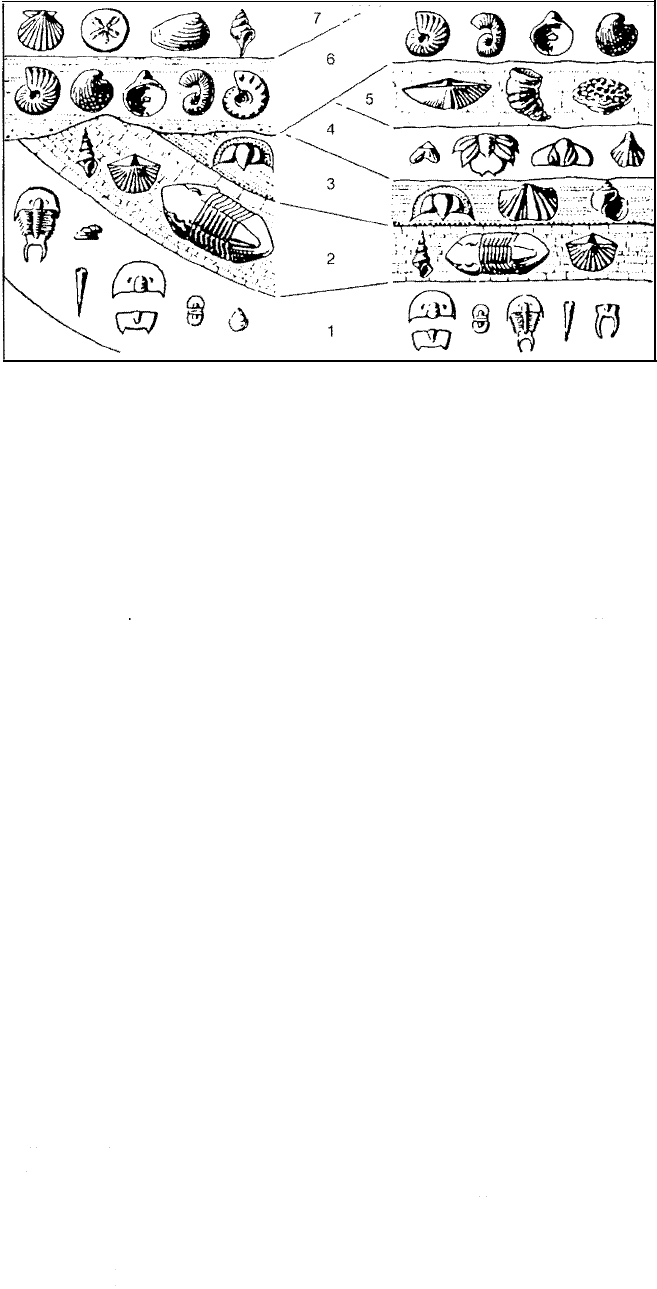
A
B
14.7 Biocorrelation
503
Figure 14.8
Diagram illustrating graphically
the principle of correlation by
fossil assemblages. [From Moore,
R. C., C. G. Lalicker, and A. G. Fis
cher, 1952, Invertebrate fossils:
McGraw-Hill Book Co., Fig. 1.3,
p. 8, reproduced by permission.]
Shaw (1964) points out, however, that the boundary between assemblage
zones is inherently fuzzy because above and below the limits of this zone will be
transition zones in which part of the characteristic fossil assemblage will be miss
ing because it has not yet appeared or has already vanished. Therefore, there are
practical limits to the accuracy that can be achieved by assemblage zone correla
tions. Part of the problem in correlation by assemblage zones stems from the fact
that the number of fossil taxa that a biostratigrapher must work with is so large
that it is difficult to visually assimilate the data and draw meaningful zone bound-·
aries (e.g., Fig. 14.9). To overcome this problem, earlier workers tended to reduce
the number of taxa whose distributions would be studied, or they tried to make
composites of the samples. A more recent solution to this problem is to apply the
techniques of multivariate statistical analysis to recognition and delineation of as
semblage zones. These quantitative techniques provide a rational statistical basis
for delineating zones based on large numbers of taxa without taking the decision
making out of the hands of the biostratigrapher (e.g., Gradstein et a!., 1985).
Correlation by Abundance Biozones
As mentioned, abundance biozones are defined by the quantitatively distinctive
maxima of relative abundance of one or more species, genus, or other taxon rather
than by e range of the taxon. ey represent a time or times when a particular
taxon was at the peak of its development with respect to numbers of individuals.
Some biostratigraphers previously used abundance zones for time-stratigraphic
correlation under the assumption that there is a time in the history of every taxon
when it reaches its maximum abundance and that this abundance peak occurs
everywhere at the same time. The current prevailing opinion among biostratigra
phers
is that most abundance zones are unreliable and unsatisfactory for time
stratigraphic correlation. This opinion is based on the apparent fact that not all
species achieve a maximum abundance, or that if they do this peak is not neces
sarily recorded by layers of abundant specimens. Furthermore, peak abundances
that are recorded in the stratigraphic record may be related to favorable local eco
logical conditions that can occur at different times in different areas and that may
persist in one area much longer than in another. Maximum abundance may thus
represent local, sporadically favorable environments, suddenly unfavorable envi
ronments that caused mass mortality, or mechanical concentrations of the shells of
organisms after death. Some of the problems of correlating by abundance zones
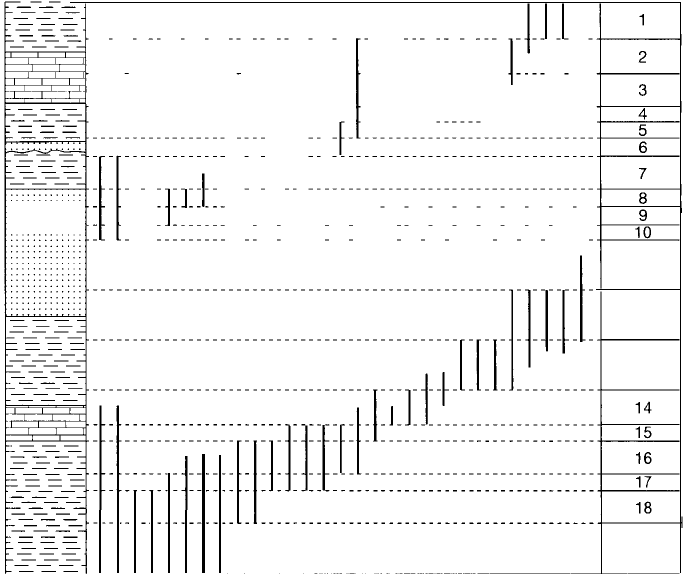
504
Chapter 14 I Biostratigraphy
Figure 14.9
Hypothetical stratigraphic sec
tion illustrating the large num
ber of fossil taxa that may be
involved in correlation by as
semblage biozones. Vertical
black lines represent the com
posite ranges of the species
found at various local sections.
The column at the right shows
one interpretation that could
be drawn from these fossil
data. [After Hazel, ]. E., 1977,
Use of certain multivariate and
other techniques in assem
blage zonal biostratigraphy:
Examples utilizing Cambrian,
Cretaceous, and Te rtiary ben
thic invertebrates, in Kauffman,
G. G., and ]. E. Hazel (eds.),
Concepts and methods of bios
tratigraphy: Va n Nostrand
Reinhold, Fig. 1, p. 289, repro
duced by permission.]
Stratigraphic
Section
Fossil Species
P
ossible
Assemblage
Zones
� �
• •
.
•
.
.
•
. . • •
.
• •
. • • .
•
. .
•
. .
•
• .
• •
. • • . • • . • .
•
. • • . • d. J
•.
l
••.
2
�-�-; = = == = = == = == = == = == = == = == = == j==
.
=. = =I= t IJ J = 1: = = = === == = == !
[·
·
·
·
·
·
·
·
·
·
···
f.[[ �·]
·
[ [·····························•
-
l
•••••••••••••
•t
� ·I� I
J.l
L
��·. ��. �� � ��. ��. �� � =�· .�
·
.�
·
.�� .�
·
.�
·
.�· .�� =�·
�1
9
0
11
1
2
13
------------------------------------------
1
9
----- --------------
are illustrated in Figure 14.10. In short, abundance zones may be used for bios
tratigraphic correlation but they do not provide a reliable means of time-strati
graphic correlation. Although they are sometimes used locally for correlation
within provinces, biostratigraphers usually prefer correlations based on assem
blage biozones or taxon-range or interval biozones.
Chronocorrelation by Fossils
Chronostratigraphic correlation is the matching up of stratigraphic units on the
basis of time equivalence. Establishing the time equivalence of strata is the back
bone of global stratigraphy and is considered by most stratigraphers to be the
most important type of correlation. Methods for establishing time-stratigraphic
correlation fall into two broad general categories: biological and physical/ chemi
cal. As mentioned, time-stratigraphic correlation by biological methods is based
mainly on use of concurrent range zones and other interval zones. Biological cor
relation methods also include statistical treatment of range-zone data and correla
tion by biogeographical abundance zones, which are biological events related to
climate fluctuations. A variety of physical and chemical methods available for
chronostratigraphic correlation are discussed in the following chapter. Logically,
this discussion of chronostratigraphic correlation by fossils also belongs in the
next chapter; however, I am including it here to keep all material relating to corre
lation by fossils in a single unit. The discussion of biocorrelation that follows rep
resents a very general introduction to this subject. For more rigorous treatment of
biocorrelation, see Gradstine et al. (1985) and Guex (1991).

14.7 Biocorrelation
505
Area 1
Relative
Abundance
Area 2
Relative
Abundance
Area 3
Relative
Abundance
Correlation by Ta xon-Range and Interval Biozones
Area 4
Relative
Abundance
Ta xon-range and interval zones are biozones that constitute the strata that fall be
tween the highest and/ or lowest occurrence of taxa. Figure 14.3 illustrates several
ways that the first and last appearances of taxa may be used to define biozones.
These different biozones have varying degrees of usefulness in time-stratigraphic
correlation, as described below.
Ta xon-Range Biozones
Taxon-range biozones may be very useful for time correlation the taxa upon
which they are based have very short stratigraphic ranges. They are of little value if
the taxa range through an entire geologic period or several periods. Correlation by
taxon range zone is often referred to as correlation by index fossils. As mentioned,
index fossils are considered to be ose taxa that have very short stratigraphic
ranges, were geographically widespread, were abundant enough to show up in the
stratigraphic record, and are easily identifiable. Unfortunatel the term dex fos
sil has also been used in other ways and can have oer connotations. Therefore, it
is
less confusing when speaking of correlation based on the entire range of a taxon
to refer to it simply as correlation by taxon-range biozone. Correlation by taxon
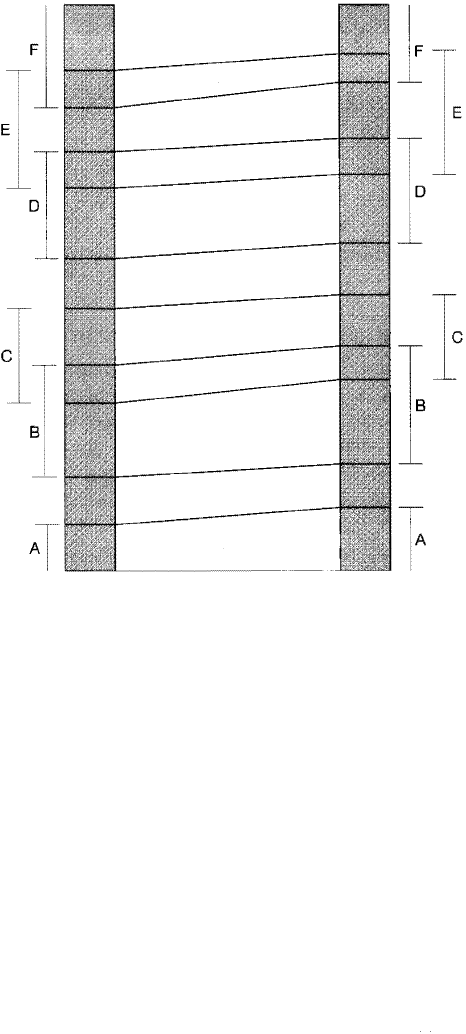
range biozone is illustrated diagrammatically in Zone 1 of Figure 14.11.
Inteal Biozones
When individual taxon-range biozones are very long, and correlation by taxon
range biozone is thus not suitable, much finer scale correlation is possible by using
other types of terval biozones. Interval biozones defined by the first (stratigraphi
cally lowest) appearance of two taxa, for example, are particularly useful in time
stratigraphic correlaon because they are based on evolutionary changes, along
phyletic lineages, that tend to occur very rapidly. Thus, the interval between the first
documented appearance of two taxa may represent a very short span of time, and
the age of the strata in this interval may be nearly synchronous throughout eir ex
tent. terval biozones defined on the last (stratigraphically hiest) appearances of
taxa are commonly considered to have less time significance than those based on
rst appearances because extinctions of taxa commonly do not occur wi the same
suddenness that new species appear through phyletic evolution.
Figure 14.11 illustrates some of the various methods that can be used for cor
relating between two stratigraphic sections on the basis of taxon-range or interval
biozones. Note from this illustration that interval biozones can be identified which
represent much shorter spans of time than that represented by the taxon-range
Figure 14.10
Schematic diagram illustrat
ing why correlation by abun
dance biozones may not
yield a true time correlation.
The same species may
achieve its maximum abun
dance at dierent times in
dierent localities. Relative
abundance increases to the
right.
506
Chapter 14 I Biostratigraphy
Section X
Section Y
Zone 10
Zone 9
Zone 8
Zone 7
Zone S
Zone S
Figure 14.1 1
Zone 4
Correlation between two hypothetical sections on the basis
of taxon-range and interval biozones. Note that several
pes of interval zones are used here for correlation. For ex
ample, Zone 1 is defined by the total vertical range of
Species A (taxon-range zone); Zone 2 is an interval zone
defined by the last appearance of Species A and the first
appearance of Species B; Zone 4 is formed by the overlap
ping ranges of Species B and C; and so forth.
Zone 3
Zone 2
Zone 1
biozones of most individual taxa. Correlation can be made also between stra
graphic sections simply on the basis of first or last appearances of specific taxa,
wiout correlating entire zones. other words, a correlation line can be drawn
from the stratiaphic position represented by the first appearance of a particular
taxon, called e first appearance datum or FAD, to the FAD of the same taxon in
another stratigraphic section. Similarly, correlation can be made between the last
appearance datums, LADs, of a given taxon in different stratigraphic sections.
FADs and LADs are further discussed in the Chapter 15.
Gra
p
hic Method fo r Correlating by Ta xon-Range Biozone
Although interval biozones can be used to define units of strata deposited during
relatively short periods of time, they do not necessarily yield precise time-strati
graphic correlations. Organisms may migrate laterally and appear other areas
at somewhat later times than their true first appearance (Fig. 14.7), or they may
migrate out of a local area before their final extinction elsewhere. These variables
of behavior make the boundaries between interval zones inherently "fuzzy." The
exact boundary between biozones can never be known because such boundaries
are determined empirically. Additional collecting a new area always holds the
possibility of extending the known range of previously defined species or taxa, be
cause
ey may have appeared earlier or persisted longer in the new area than
the origally defined areas. One way to minimize the problem of fuzzy zonal
boundaries is to treat range data statistically, utilizing the first and last appear
ances of all the species present in a stratigraphic section rather than the ranges of
just one or two species. A. B. Shaw in 1964 was the first to propose a graphical
method for establishing time equivalence of strata in two stratigraphic sections by
plotting first and last appearances of all the species in one section against the rst