Boggs S. Principles of Sedimentology and Stratigraphy
Подождите немного. Документ загружается.

14.4 The Basis for Biostratigraphic Zonation: Changes in Organisms Through Time
487
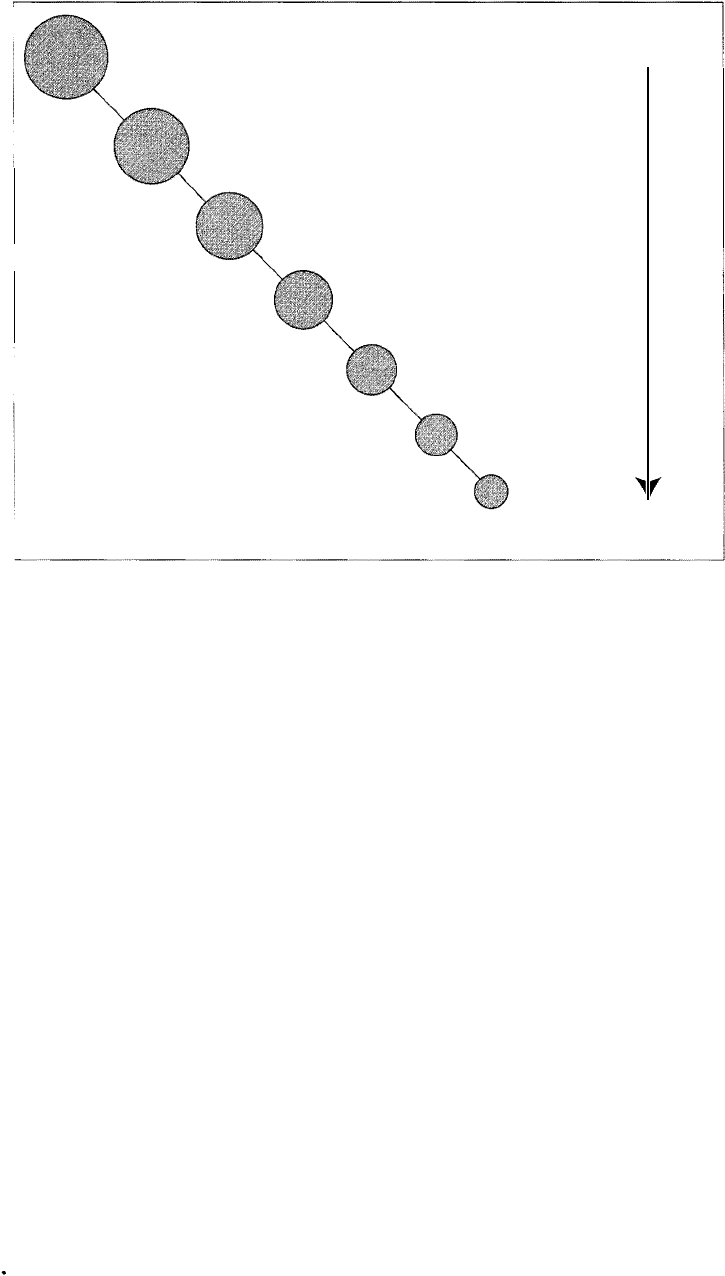
Kingdom (five)
Phylum (-ninety)
Class
Order
Family
Genus
Species
(millions)
l
E
o
E
·mE
0
�
u
£0
characterized mainly on the basis of shell, or skeletal, morphology. Because the
skeletal morphology of different members of the same species can be quite vari
able, determination of fossil species must be made by taxonomic specialists. Such
determination may require quantitative measurements of shell parameters and
computer analysis of measurement data to provide statistical rigor to fossil species
idenfication.
Changes in Species Through Time
The importance of species in biostratigraphic study lies in the fact that species do
not remain immutable for all time. If environmental conditions remained ab
solutely constant through time, perhaps species would change very little. The fact
is that environments do change and, as they change, species also change, although
environments do not directly cause species to change. Both gene mutation, or
gene pool combinations, and shifting environmental conditio are essential to
the evolution of species. Most species are well adjusted to their normal environ
ments, but if an appropriate variation appears in a species just at the time when it
is becoming inadaptive a chang environment, the force of natural selection
may preserve this novel variant (e.g., Shaw, 19). Thus, species have evolved
through time as a result of natural selection of those random, chance mutaons
that brought the species into better adjustment with changing environmental con
ditions.
All indications from the geologic record suggest that species variations are
one-directional and nonreversible. Once a species has become extinct, it does not
reappear
in the fossil record. As members of a new species increase in numbers,
they may eventually become abundant and widespread enough to show up in the
geologic record as the first appearance of the species. When the species is no
longer able to adjust to shifting environmental conditions, its members decrease
Figure 14.5
Schematic representation of
the hierarchical Linnaean
system for classifying or
ganisms. All organisms are
currently grouped into five
kingdoms, about ninety
phyla; numerous classes,
orders, families, and gen
era; and millions of
species. Note that organ
isms at the species level
share many common char
acteristics whereas those at
higher levels share fewer
characteristics.
488
Chapter 14 I Biostratigraphy
Figure 14.6
in number and eventually disappear-the extinction, or last appearance, of e
species. Extinction refers to the disappearance by death of every individual mem
ber of a species or higher taxonomic group so that the lineage no longer exists. Pa
leontologists recognize also that a species may experience pseudoextinction.
Pseudoextinction, or phyletic extinction, refers to an evolutionary process where
by
a species evolves into a different species. Thus, the original species becomes ex
tinct, but the lineage continues in the daughter species.
Some species exist for only a fraction of a geologic period. Others may per
sist for longer periods of time. Organisms that were abundant and geographically
widespread and had relatively short ranges have the greatest time-stratigraphic
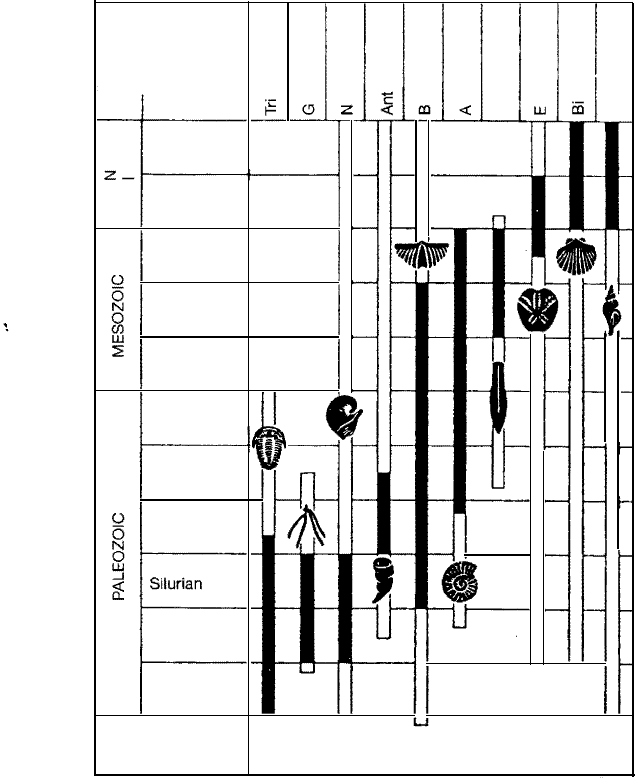
utility, that is, the greatest usefulness for biostratigraphic study (Fig. 14.6). Figure
14.6 shows some of the more important groups of macrofossils that are useful for
biostratigraphic zonation. Many other fossil groups, including microfossil groups
such as foraminifers, are also important (see Figure 11.4. for example). Particular
ly important and useful fossils for biostratigraphic purposes are called guide fos
sils or index fossils. Ideally, index fossils should be (1) independent of their
environment, (2) fast evolving, (3) geographically widespread, (4) abundant, (5)
readily preserved, and (6) easily recognizable (e.g., Doyle, Beett, and Baxter,
1994, p. 37).
Geological
Time Scale
Eras Periods
�
§
0
.
Quaterna
6
z_
� 2 Tertiary
Cretaceous
Jurassic
Triassic
0
Q
0
0
N
E
0
>
�
�
z
--
-
-
-
Permian
�-
-
Carbon
iferous
Devonian
'
'
0
0
N
0
E
0
·
E
E
E
E
>
i
-
•
-
-
-
-
-
-
-
-
-
- -
Some of the more important macrofossil groups
of marine invertebrate organisms for biostrati
graphic zonation. The white columns show the
time span of distribution, the black columns the
time span in which the organisms are important
as index fossils. [From Thenius, E., 1973, Fossils
and the life of the past, Fig. 50, p. 79, reprinted
by permission of Springer-Verlag.]
Ordovician
Cambrian
- -
14.4 The Basis for Biostratigraphic Zonation: Changes in Organisms Through Time
489
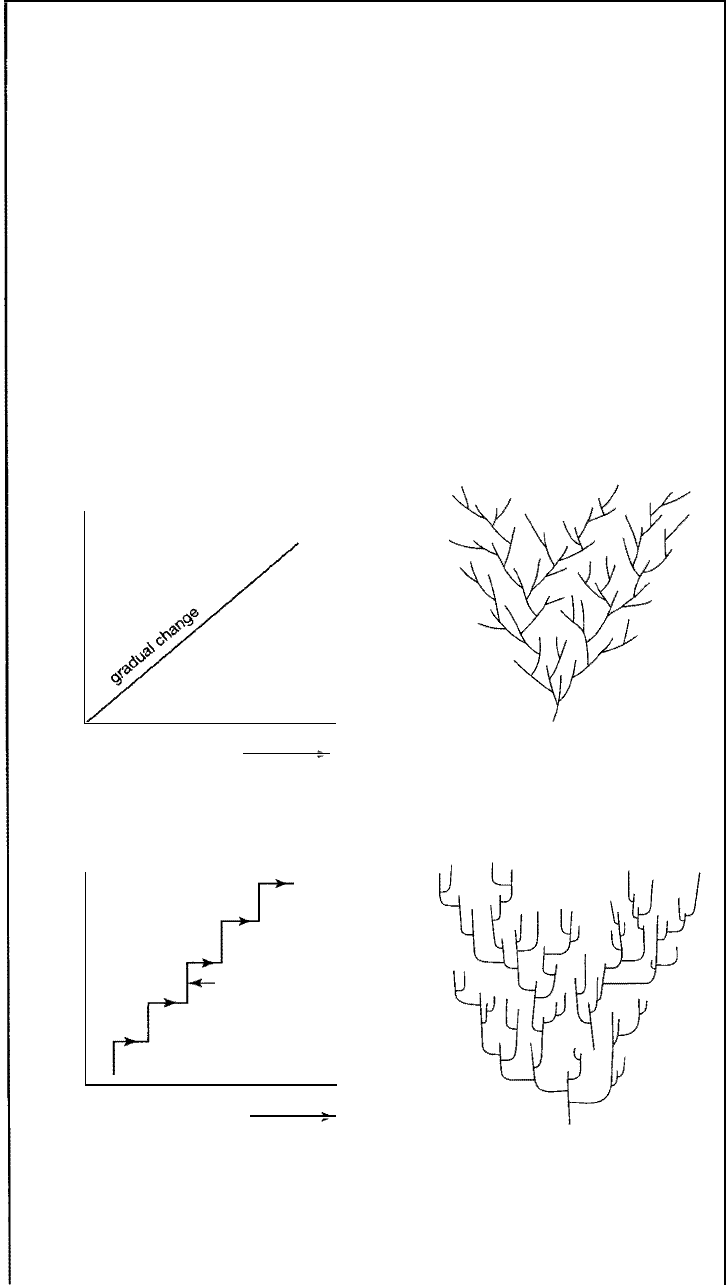
Box 14.1 Models and Rates of olution
There is currently considerable controversy among paleontologists conceing
the mode of change in organic evolution. Tw o principal points of view prevail.
e view states that evolution proceeds mainly as a gradual change by slow,
steady transformation of well-established leages-phyletic evolution, or
gradualism. The gradualist concept has been the traditional view of species
evolution. The second view holds that many species arise very rapidly from
small populations of organisms that have become isolated from the parental
range and then subsequently change very little after their successful origin.
This latter view represents evolution by speciation or branching of lineages,
the so-called punctuated equiliba model of Eldredge and Gould (19).
Thus,
according to this theory, fossil populations are in stable equilibrium for
long periods of time and change very little (called stasis), punctuated by sud
den introduction of new species. Dierences in these two postulated modes of
evolution are illustrated graphically in Figure 14.1.1. In the punctuational
model, speciation, or branching of spees, is viewed as a very rapid process,
A
.
Phyletic gradualism
1
E
Morphology
(body characteristics)
B. Punctuated equilibrium
1
E
stasis (unchanging
morphology)
� punctuation (speciation)
Morphology
(body characteristics)
Figure 14.1.1
Schematic 'family tree'
Schematic 'family tree'
Diagrammatic representation of gradualistic and punctuated models of evolution.
[Schematic family trees after Stanley, W. H., 1979, Macroevolution, patterns and
processes, W. H. Freeman and Company.]

490
Chapter 14 I Biostratigraphy
requiring only tens of thousands of years or possibly as little as a few hundred
years (e.g., Stanley, 1979) after a population becomes reproductively isolated
from the parent population. Although the duration of species from first ap
pearance to extinction may be measured in mions of years (Table 14.1.1),
species are believed by the punctuationalists to change morphologically very
little and only very slowly after initial speciation. This concept is stated very
succinctly by Eldredge and Gould (1977), who emphasize the importance of
speciation (splitting) and claim "that most morphological differences between
two species appear in conjunction with the speciation process itself, whereas
most of a species' history involves little further change, at least of a progres
sive nature."
Whether species evolution takes place mainly by gradual evolutionary
change, mainly by punctuated speciaon, or by both, is still a much discussed
and debated issue (see, for example, Gould and Eldredge, 1993, and Sheldon,
1996). Bo sides of the controversy contue to be aired in e paleontological
literature. Some workers propose that certain groups of organisms, such as
mammals, tend to evolve by gradual transformation whereas others, such as
many marine invertebrates, tend to evolve by punctuated equilibrium. Sheldon
(1996) suggests that organisms on land in e tropics and those in the deep sea
may tend to undergo connuous, gradualistic evolution, whereas organisms in
temperate zones and shallow water tend more toward stasis and occasional
punctuations. Because the majority of the fossil record comes from dynamic
shallow-marine environments, many fossil lineages thus show approximate sta
sis and occasional punctuations. Dierent groups of species are known to evolve
at greatly dierent rates. Stanley (1985) indates, for example, that marine bi
valve groups evolve at a rate that yields only ree or four species in 20 million
years. By contrast, mammalian families evolve at a rate that yields roughly 80
species in 20 million years. From a practical point of view, the task of delineang
Biological group
Marine diatoms
Benthic foraminifers
Planktonic foraminifers
Bryophytes
Marine bivalves
Marine gastropods
Higher plants
Ammonites
Freshwater fish
Graptolites
Beetles
Snakes
Mammals
Trilobites
Source: Stanley (1985).
Estimated mean species duration (Ma)
25
•* 20-30
>20
>20
11-14
10-14
8->20
-5 (but with a mode in the 1-2 Ma range)
3
2-3
>2
>2
�1-2
>1

14.4 The Basis for Biostratigraphic Zonation: Changes in Organisms Through Time
491
the boundaries of species is more difcult if evolution occurs by phylec grad
ualism because a chain of intermediate species is present in the geologic record
and the boundaries between successive species are arbitrary. Thus, it is diffi
cult to pick points in the evolutionary sequence at which distinct species
boundaries are recognizable. If, on the other hand, evolution occurs by specia
tion (punctuated equilibrium), most morphological change presumably occurs
at branch intersections in the evolutionary line (Fig. 14.1.1B), which represent
discrete points in time. Thus, the task of picking species boundaries should
theoretically be easier and less error prone if evolution occurs by speciation
rather than by phyletic gradualism. On the other hand, the initial appearance
of a new species in different provinces may show a time lag owing to lags in
migration, which makes identification of the first-appearance species bound
ary more difficult.
The practicality of identifying species boundaries, and of establishing the
boundaries of biostratigraphic zones, is further complicated by problems in
volving the following: (1) sampng intervals (How small must they be to en
sure that species boundaries are detected?), (2) changes in the fossil record
induced by burial and the vagaries of preservation, (3) constancy and rates of
sedimentation (smaller sampling intervals are required for sediments that ac
cumulated very slowly vs. those that accumulated very rapidly), and (4) inter
mittent or punctuated patterns of sedimentation and erosion that yield an
incomplete stratigraphic record, thus giving the appearance of punctuated
speciation. [For some comparatively recent views on punctuated equilibrium,
see Gould (2001) and Kemp (1999, Chapter 7).]
DETERMINISTIC VS. PROBABILISTIC EVOLUTION
An interesting side issue to the problem of evolutionary controls relates to the
question of whether or not such evolutionary events as adaptive radiation and
periods of mass extinction are deterministic or probabilistic. That is to say, are
evolutionary events explainable only in terms of causal factors, or are there
statistical laws or generalizations that can explain these events on the basis of
random variations or processes? Every human, for example, is destined from
the instant of his or her birth to age and eventually die. Is every species like
wise destined om the time of its birth (initial speciation) to eventually age
and become extinct? Raup (1991, p. 6) states that there is absolutely no basis
for equating the life span of species with those of humans and that there is no
evidence of aging in species or any known reason why a species could not live
forever. Nonetheless, in a subsequent chapter of his book entitled "Gambler's
Ruin and Other Problems," Raup discusses the probability of extinction of a
genus with a limited number of species (e.g., ten). If the chance of extinction is
identical to that of speciation (fifty-fifty), the number of species will uctuate
up and down as in a random walk but will finally reach zero. Therefore, the laws
of probability suggest that eventual extinction of the genus is inevitable (Raup,
1991, p. 49), although the greater the number of species in e genus the longer it
will take for extinction to occur.
Probabilistic evolutionary models are called stochastic models. Van
Val en (1973, p. 1) asserted, for example, that "all groups for which data exist go
extinct at a rate that is constant for a given group." Such statements should not
be taken to mean that extinctions occur without cause. Extinction of a species
may be the result of any number of specific causes and it may, therefore, be in
valid to attribute the death of individuals to chance. If frequency of death is
considered at population levels, however, it may be mathematically valid to
describe the frequency as being governed by random stochastic processes. In

492 Chapter 14 I Biostratigraphy
other words, individuals a given population of organisms may die owing to
various specific causes; however, the population as a whole will become ex
tinct at a constant rate, depending upon its size, regardless of the specific caus
es of death of the individuals. Thus, in the stochastic approach, the pattern of
evolution as a whole is perceived to be a random process, although individual
fluctuations in this patte can be explained by cause and effect.
Stochastic models may serve to separate tho
�
e features of the evolution
ary record that are amenable to deterministic explanations from those that do
not warrant a search for a specific cause. For example, the fairly rapid demise
of e dinosaurs at e end of Cretaceous time can probably be explained by
some specific environmental or catastrophic event such as dramatic climatic
change resulting from meteorite impact; however, the gradual decline of co
nodonts and their eventual extinction at e end of the Triassic appears to be
more difficult to attribute to a specific cause or causes. Probabilistic extinction
can be thout of as a kind of "background" extinction that has acted through
out the fossil record, almost but not quite keeping pace wi the rise of new
species (global diversity of organisms has increased with time). From time to
time, however, major extinction events, called mass extinctions, have occurred
and demand a specific causal explanation.
MASS EXTINCTIONS
The fossil record shows that the diversity of both marine and continental life
has increased exponentially since the end of the Precamqrian (e.g., Benton,
1995; Miller, 2000); however, the record also shows that many groups of or
ganisms became extinct or suffered dramatic reductions in numbers and di
versity at particular times. During the past two decades, ese episodes of
mass extinction have assumed increasing importance to paleontologists and
other geologists, judging by the rapid appearance of new articles and books
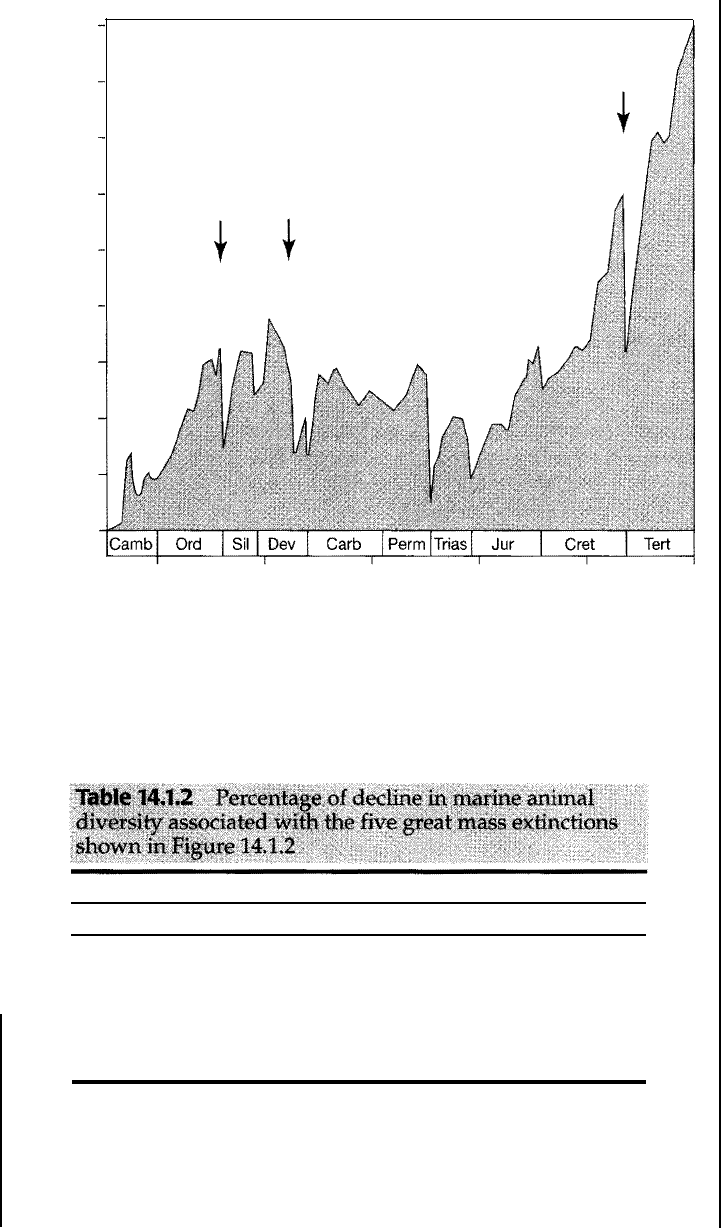
dealing wi mass extincons. Five extinction events have become so impor
tant and far reaching that they are now commonly referred to as the big five.
These major extinction episodes took place near the end of the Ordovician, De
vonian,
Permian, Triassic, and Cretaceous (Fig. 14.1.2), and the later extinc
tions affected both terresial and marine forms.
As shown in Table 14.1.2, 47-82 percent of extant mane animal genera
became extinct during these five major mass extinction episodes and 151
percent of the marine animal families became exnct. Such dramac extinc
tions demand an explanation linked to some specific cause or causes. Particu
larly important groups of organisms that became extinct include trilobites and
Fusulinid foraminifers (Late Permian), conodonts (Late Triassic), and am
monites and dinosaurs (Late Cretaceous). Many other groups also became ex
tinct or were greatly reduced in numbers. ese dramatic extinctions have
taxed the agination of paleontologists and other geologists to provide ac
ceptable causal explanations. The Late Permian extinction phase has received
particular attention because of the number of major groups affected and the
sharpness of the change wi which these groups disappeared from the geo
logic record at the end of the Permian.
Mass extinctions are of enormous interest to geolosts because of the
questions they raise, among oer things, about possible recurring catastrophic
events in Earth's history. e past few decades saw explosive growth in re
search into the pattes, rates, causes, and consequences of extinction but lit
tle overall agreement about the causes of extinction. Theories about extinction
fall to three groups: catastrophic extinction, gradual extinction, and stepwise
4000
3000
.
c
0
2000
E
z
1000
0
500
14.4 The Basis for Biostratigraphic Zonation: Changes in Organisms Through lime
493
t
t
400
300
200
100 0
Geologic time (millions of years)
Figure 14.1.2
Diversity of marine animal genera (number of genera) during Phanerozoic time. Arrows
point to the "big five," the five great mass extinctions. [After Sepkoski, ). )., Jr., 1995,
Patterns Phanerozoic extinction: a perspective from global data bases, in Wa lliser,
O.H. (ed.), Global events and event stratigraphy: Springer-Verlag, Berlin, Fig. 1, p. 38.]
Mass-Extinction Event
End-dovician
Late Devonian
End-Permian
End-Triassic
End -Cretaceous
urce: Sepkoski (1995).
Percent decline in diversity
Genera Families
60
26
57
22
82
51
53
22
47
16
extinction (extinction that occurs in a series of discrete steps in the vicinity of
major stratigraphic bodaries, such as the Permian/Triassic boundary). ese

494
Chapter 14 I Biostratigraphy
various theories have been expounded in numerous research papers and a
number of recent books, some of which are listed at the end of ts chapter.
Causes proposed for the main mass-extinction events are summarized in
Figure 14.1.3. Proponents of the catastrophic theory, especially for the sharp
Cretaceous/Tertiary boundary event, suggest that the impact of extraterrestri
al objects called bolides (meteorites and comets) created major climatic change
(global winter) by throwing up huge clouds of dust and/or generated acid
rain, tsunamis, and wildfires that caused extinction of some taxonomic
groups. Alteatively, intense explosive volcanic activity may have adversely
aected climates through discharge of excessive gas douds (greenhouse
warming). Other geologists suggest that such extraterrestrial causes are not
needed to explain most exnction events. Gradual, progressive changes in cli
mate (either warming or cooling) together with changes in sea level are ade
quate, they say, to account for extinctions. For example, lowering of sea level
during major episodes of regression reduces habitats for shallow-water organ
isms and increases competition. On the other hand, widespread marine trans
gressions appear to be linked to development of anoxic (low oxygen) conditions
that adversely affect some organisms and cause extinction (Hallam and Wig
nall, 1997, p. 251). The actual causes of anoxia during phases of transgression
are poorly understood. Sll other geologists suggest that some extinctions
occur in a stepwise fashion by a series of pulses-some before, some at, and
some just after a major boundary. These extinctions are presumably the result
of a succession of events, such as brief showers of comets superimposed on a
background of progressive environmental deterioration.
In any event, worldwide extinctions of major groups of organisms, while
extremely interesting and significant, play only a limited role in biostratigra
phy because these major extinctions provide only a few correlation horizons.
Changing local environmental conditions are probably a more significant
cause of extinction of individual species, which form e most important basis
for biostratigraphy.
L
ate
Pr
ecamb
r
ian
-------------------------------------------------------------------------------•
L
ate
E
a
r
ly
Camb
ri
an
-----------------------------------------------------------•---------------•
L
ate Camb
r
ian
-------------------------------------------------------------------------------•
L
ate
O
r
dovician
---------------------------------------•------- ----•----------•--------------•
L
ate Devonian
------------------------------------------------------------------------------•
Dev
o
ni
a
n
-
Ca
r
bonife
r
ous
--------- -------------------------------------------------------------•
L
ate
P
e
r
mian
--------------------------------------------------------------------•
E
nd-
P
e
r
mian
-----------···-----------------•------------------··· -•------------------------•
E
n
d
-
T
r
iassic
-------------------------------------------- ----------------------- _ --------------
E
a
r
ly
J
u
r
assic
-----------------------------------------------------------------------------------•
L
ate C
r
etaceous
-----------------------------------------------------------------------------•
End
-
C
r
etaceous
-----------
•
-------------•---------•-----------------------•---------------0
End-
P
aleocene
----------------------------•
----------------------•---------------------------
•
L
ate
E
o
cene
-------------------------------------------•
•
st
r
on
g
lin
k
o possible l
in
k
Figure 14.1.3
Proposed causes for the main Phanerozoic extinction events. [After Hallam, A., and
P. B. Wignall, 1997, Mass extinctions and their aftermath: Oxford University Press,
Table 11.1, p. 248.]

14.5 Distribution of Organisms in Space: Paleobiogeography
495
14.5 DISTRIBUTION OF ORGANISMS IN SPACE:
PALEOBIOGEOGRAPHY
When d'Orbigny introduced the concept of stage, he believed that the fossil assem
blages upon which his stages were based had worldwide distribution. We now
know that the fossil species and assemblages that characterize biostratigraphic
units are not necessarily present everywhere that rocks of the appropriate ages
occur. Few species are distributed throughout the entire world. Most, in fact, are
restricted in their geographic range, although some fossil groups ranged widely
throughout whole ecological realms at times in the geologic past. The region with
in which a particular group or groups of plants or animals is distributed is called
a biogeographic province. Biogeographic provinces are separated by physical or
climatic barriers. Land areas are barriers to marine organisms; open marine water
is a barrier to land animals and plants; deep water is a barrier to shallow-water,
shelf-dwelling organisms; cold water is a barrier to warm-water organisms; fsh
water is a barrier to organisms adapted to saline marine conditions; and so forth.
A particular type of barrier may be impenetrable by one species of organism but
not by another. For example, benthonic organisms that do not have a long-lived,
juvenile planktonic larval stage find deep water a barrier to dispersal. By contrast,
planktonic organisms, which live in near-surface waters in the ocean, are distrib
uted widely throughout the oceans in both shallow and deep water.
Box 14.2 Dispersal of Organisms
Paleontologists regard species as the fundamental biologic units in nature.
They are the basic unit that undergoes evolution; the species niche is the basic
functional unit in ecological interactions; and species are the fundamental
units of biogeography, biostratigraphic zonation, and correlation. Because of
their central importance in biostratigraphy, it is essential that we understand
the factors that control the dispersal and distribution of species. Obviously,
factors affecting the dispersal of land organisms and plants are different from
those that control the dispersal of marine organisms. Also
,
the distribution of
invertebrate marine organisms is controlled by different factors than those that
control the distribution of vertebrate marine groups. Because of the overriding
importance of marine invertebrates in biostratigraphic studies, we shall con
fine our discussion of dispersal here to invertebrate organisms in the marine
setting, which consists of the pelagic realm (the water column) and the benth
ic realm (the seafloor, or bottom environment) (Fig. 14.2.1).
Marine invertebrate organisms can be divided into three fundamental
types on the basis of habitat: plankton, nekton, and benthos (Table 14.2.1).
Plankton are mainly microscopic-size organisms that live suspended at shal
low depths within the water column (pelagic realm) and have very weak or
limited ability to direct their own movements. They are distributed more or
less passively by currents and wave action and may be dispersed widely into
all types of open-ocean environments. Planktonic organisms are exceptionally
useful fossils for biostratigraphic zonation and correlation because of their
widespread distribution. They reflect e habitat of the pelagic realm and not
the bottom environment into which they fall upon death; therefore, their pres
ence in ancient marine sedimentary rocks is of limited value in environmental
interpretation, although they are useful in some paleoceanographic applica
tions (e.g., terpretation of water paleotemperature from oxygen isotopes). A
few plankton such as graptolites do have some value as indicators of bottom

496
Chapter 14 I Biostratigraphy
Pelagic
r
ealm: Planktonic and nekton
i
c o
r
ganisms
� Benth
i
c
r
ealm: Benthonic o
r
ganisms
Figure 14.2.1
0
1000
2000
3000
4000
5
000
6000
Subdivision of the marine environment into the pelagic (water column)
and benthic (bottom) realms. The pelagic realm is inhabited by planktonic
and nektonic organismsi benthonic organisms occupy bottom environ
ments of the benthic realm.
enviroents. Graptolites were too fragile to survive in high-energy, shallow
water environments and are hence preserved mainly in the facies of quiet
water environments. Thus, they constitute "facies fossils."
Nekton also inhabit the pelagic realm and include all animals that are
able to swim freely. Modern nekton are distributed in the ocean at depths rang
ing from the surface to thousands of meters and encompass many advanced
groups of animals such as fish, whales, and mammals. Nekton are less abun
dant in the fossil record than planktonic and benthonic organisms (below) and
thus overall appear to have somewhat less value in biostratigraphic studies.
Nektonic fossils include fish remains in some deep-sea clays, belemnites and
other mobile cephalopods, and probably conodonts. Conodonts are an inter
esting type of fossil, with considerable biostratigraphic significance, that oc
curs in rocks ranging in age from Cambrian to Triassic. They are tin
toothlike phosphate fossils whose origin remained an enigma until 1982. A
complete specimen of the conodont-bearing animal has been found in Lower
Carboniferous rocks of the Edinburgh district, Scotland (Briggs, Clarkson, and
Aldridge, 1983). The specimen is an elongate, soft-bodied animal 40 mm long
and 1.8 mm wide. The conodont apparatus occurs in the head or anterior re
gion of the body and may have served as teeth or possibly some type of inter
nal support.
Benthos are bottom-dwelling orgasms at live either on or below the
ocean floor (benthic realm). Benthos (benthonic organisms) with preservable
hard parts are particularly important for environmental interpretation because