Березов Т.Т., Коровкин Б.Ф. Биологическая химия
Подождите немного. Документ загружается.

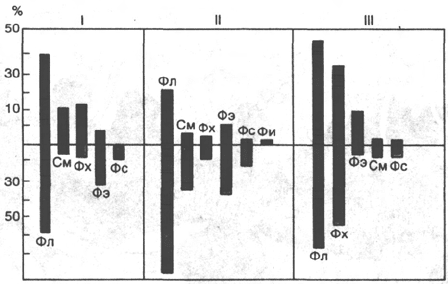
Рис. 9.3. Распределение липидов между наружной (а) и внутренней (б) сторонами
бислоя в мембранах эритроцитов (I), вируса гриппа (II) и саркоплазматического
ретикулума кролика (III).
Фл - общие фосфолипиды; Фх - фосфатидилхолин; Фэ - фосфатидилэтаноламин; Фс - фосфати-
дилсерин; См - сфингомиелин; Фи - фосфатидилинозит.
ные цепи, метальные концы которых соединены друг с другом ковалент-
ными связями. Такие «шпильки» обеспечивают повышенную прочность
липидного бислоя. (Подробнее о жирных кислотах см. главу 11.)
Липиды в составе бислоя распределяются асимметрично. Это свойство
диктуется особенностями строения их молекул: фосфатидилхолину, фосфат-
идилсерину, сфингомиелину присуща цилиндрическая форма, фосфатидил-
этаноламину – форма конуса, а лизофосфолипидам (получаются в резуль-
тате отщепления от молекулы одной жирнокислотной цепи) – форма пере-
вернутого конуса. Природные мембраны также обладают исходной асим-
метрией (рис. 9.3).
Белки взаимодействуют с мембранным бислоем, в результате чего они
либо ассоциируются с поверхностью мембраны – периферические белки,
либо пересекают бислой один или несколько раз, прочно интегрируясь
в него,– это интегральные белки. Интеграция оказывается возможной, если
в первичной структуре белка имеются достаточно протяженные участки,
содержащие гидрофобные аминокислотные последовательности. В таком
случае белковые молекулы способны самопроизвольно встраиваться в би-
слой. При ассоциации рибосом с мембранными структурами встраивание
гидрофобных белков в мембрану осуществляется синхронно с их синтезом
при участии специальных механизмов, потребляющих энергию АТФ.
Участки белка, которые обращены во внеклеточную среду, могут под-
вергаться гликозилированию. В мембранах растений и бактерий полисахара
играют самостоятельную роль, образуя наружную оболочку. В клетках
животных, в которых наружный слой включает углеводы, имеется внутрен-
ний цитоскелет, состоящий из актина и других легко полимеризующихся
белков; он имеет регулярную связь с мембранными белками и выполняет
формообразующую и опорную функцию (рис. 9.4).
Фазовое состояние мембранных липидов. Мембранные липиды могут
находиться в нескольких фазовых состояниях, т. е. они обладают мезомор-
физмом. Два основных ламеллярных состояния, характерных для мембран-
ных липидов в клеточных системах: кристаллическое и жидкокристалличе-
ское – различаются плотностью упаковки и подвижностью находящихся
в бислое белковых молекул. При более плотной упаковке ацильные цепи
301
а
б
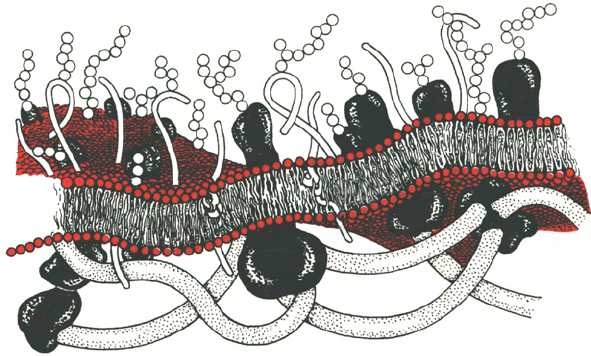
Рис. 9.4. Схематическое изображение клеточной мембраны.
липидов расположены под углом, близким к 90°, и все С—С-связи нахо-
дятся в транс-конформации (максимально вытянуты). Фазовый переход
приводит к увеличению подвижности ацильных цепей в бислое, увеличению
угла их наклона и уменьшению плотности упаковки. Латеральная подвиж-
ность мембранных белков после фазового перехода возрастает, увеличива-
ется вероятность образования их ассоциатов.
В липидном бислое могут также образовываться гексагональные струк-
туры (вывернутые мицеллы). При их образовании в мембране возникают
дефекты регулярной упаковки, что позволяет проникать через мембрану
крупным молекулам, а также обеспечивает обмен компонентами монослоев
в бислойной мембране.
Фазовые переходы мембранных липидов могут быть вызваны изменени-
ем температуры среды. Значение температуры, при котором наблюдается
фазовый переход, называется критической температурой фазового перехода,
или разделения фаз, если различные участки мембраны вследствие гетеро-
генности липидного состава по-разному отвечают на изменения темпера-
туры. Ионы Са
2+
, изменение числа ненасыщенных жирнокислотных цепей
мембранных фосфолипидов и некоторые другие факторы также могут
индуцировать фазовые переходы в бислое. Обычно критическая температу-
ра фазовых переходов приближена к температуре тела гомойотермных
животных (или к температуре среды обитания пойкилотермных животных).
Таким образом, достаточно незначительного изменения условий, чтобы
изменить упаковку мембраны.
Специфические свойства биологических мембран. Благодаря указанным
особенностям биологические мембраны имеют присущие им характерные
черты. Они образуют протяженные бислойные структуры малой толщины
(6–10 нм), объединяющие белковые и липидные компоненты с различными
свойствами.
Целостная структура мембраны создается за счет гидрофобных и элект-
ростатических взаимодействий, а не за счет ковалентных связей между
составляющими ее молекулами белков и липидов. Гидрофобный липидный
бислой представляет естественную преграду для проникновения полярных
молекул. Мембраны асимметричны по своему исходному строению, что
302
Гликокаликс
Цитоскелет
Мембранный бислой
с встроенными в него
белками
обеспечивает градиент кривизны и спонтанное образование замкнутых
структур.
Мембранный бислой обладает относительно малой микровязкостью.
Другими словами, мембраны рыхло упакованы, что позволяет отдельным
компонентам проявлять высокую подвижность в латеральном направ-
лении.
Наружные мембраны клеток отличаются от внутренних по липидному
составу (последние почти не содержат стеринов, имеют соотношение
ФХ/ФЭ > 1) и обладают специфическим набором ферментов и рецепторов.
Как правило, белки плазматических мембран со стороны внеклеточной
среды обильно гликозилированы. Внутриклеточные мембраны содержат
мало гликопротеинов и гликолипидов и характеризуются меньшей микро-
вязкостью. Благодаря этому они могут образовывать органеллы малого
размера. Мембранные белки выполняют различные специфические функ-
ции: рецепторные, транспортные, ферментативные, энергопреобразующие
и т.д. (см. далее).
Функции биологических мембран. Как отмечалось, клеточные мембраны
отграничивают содержимое клетки (или клеточной органеллы) от окружаю-
щей среды. Благодаря наличию специальных рецепторов они воспринима-
ют сигналы из внешней среды (например, молекулы гормонов, называемые
первичными мессенджерами, или посредниками), в ответ на которые образу-
ются вторичные мессенджеры, высвобождающиеся внутрь клетки. Так осу-
ществляется преобразование сигналов, изменяющих клеточный метаболизм
в соответствии с изменяющимися условиями среды (см. главу 8).
Мембранные рецепторы выполняют функции узнавания (иммунокомпе-
тентная система), адгезии (обеспечение межклеточных контактов, формиро-
вание тканей), регуляции активности ионных каналов (электрическая возбу-
димость, создание мембранного потенциала). Мембранные ферменты в со-
ставе бислоя приобретают большую стабильность и способность к осуще-
ствлению реакций, которые в гидрофильном окружении протекали бы
с весьма малой скоростью. Липидное окружение предоставляет таким
белкам «привилегированные» условия функционирования, но и накладыва-
ет ограничения на поведение белковых ассоциатов: последнее сильно
зависит от плотности упаковки (микровязкости) мембран. Поэтому факто-
ры, влияющие на липидный состав и свойства клеточной мембраны,
оказывают регулирующее влияние на функции мембранных белков.
Мембранные белки часто образуют олигомерные ансамбли, взаимодей-
ствия между которыми (или длительность их существования в бислое)
оказывается под контролем их мембранного окружения. Изменения микро-
вязкости мембран в таком случае позволяют контролировать активность
этих надмолекулярных структур.
Транспортная функция является одной из важных функций клеточных мембран
(рис. 9.5). Мембрана создает существенные ограничения для проникновения различ-
ных веществ, однако она не является полностью непроницаемой: небольшие
нейтральные молекулы могут проникать через бислой в области структурных
дефектов. Этот процесс осуществляется по градиенту концентрации переносимого
вещества - из области, где его содержание высоко, в область с более низким
содержанием. Такой процесс называется простой диффузией, он осуществляется
неизбирательно и с низкой скоростью.
При облегченной диффузии вещества также переносятся в направлении их
концентрационного градиента, но с использованием специальных структур - пере-
носчиков или каналов, увеличивающих скорость и специфичность переноса. Извест-
303
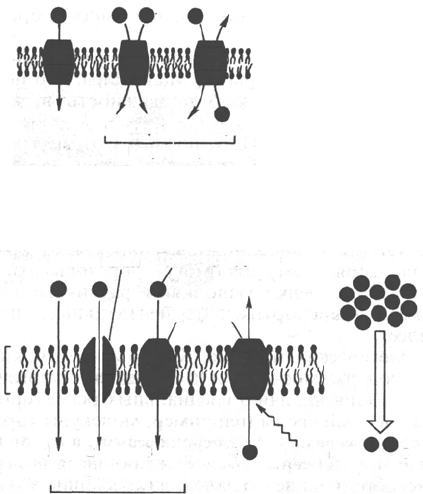
Рис. 9.5. Перенос веществ через мембрану.
а - виды переноса; б - пассивный и активный транспорт: 1 - пассивная диффузия; 2 - диффузия
с помощью канала; 3 - диффузия с помощью переносчика; 4 - активный транспорт; 5 - вторич-
но-активный транспорт.
ны высокоспецифические транслоказы - белковые молекулы, переносящие аденило-
вые нуклеотиды через внутреннюю мембрану митохондрий: Nа
+
/Са
2+
-обменник -
белок, входящий в состав плазматических мембран многих клеток; низкомолекуляр-
ный пептид бактериального происхождения валиномицин - специфический перенос-
чик для ионов К
+
.
Облегченная диффузия, осуществляемая с помощью каналов, не обладает высо-
кой специфичностью (специфичность определяется лишь размерами канала), но
протекает с большей скоростью, а процесс переноса не достигает насыщения
в широком диапазоне концентраций переносимого вещества. Функционирование
каналов в меньшей степени зависит от фазового состояния мембраны, чем функцио-
нирование переносчиков. Все эти примеры относятся к пассивному транспорту через
мембрану.
Активный транспорт веществ осуществляется такими же механизмами, но проте-
кает против концентрационного градиента и для своего осуществления должен быть
сопряжен с энергодающим процессом. Основным источником энергии для активно-
го транспорта является АТФ. Поэтому, как правило, эти системы представляют
собой АТФазы. Примером систем активного транспорта ионов является Na
+
/K
+
-
АТФаза плазматических мембран животных клеток, которая «выкачивает» из
клетки ионы натрия в обмен на ионы калия, затрачивая на выполнение этой работы
АТФ в стехиометрии 3Nа
+
/2К
+
/1АТФ. Са
2+
-АТФаза осуществляет активный
транспорт кальция через мембрану со стехиометрией 2Са
2+
/1АТФ.
В так называемых сопрягающих мембранах имеются протонные насосы,
работающие как Н
+
-АТФазы. В результате их функционирования на мембране
304
Унипорт
Симпорт
Антипорт
Котранспорт
а
Бислой
Канал
Транспортируемая
молекула
Энергия
Диффузия
б
Переносчик
1
2
3
4
5
возникает разность концентраций протонов (ΔрН) и разность электрических потен-
циалов, в совокупности образующие протонный электрохимический потенциал,
обозначаемый ΔμН
+
(см. далее). За счет работы Н
+
-АТФазы создается кислая среда
в некоторых органеллах клетки (например, лизосомах, хромаффинных клетках
надпочечников). В митохондриальной мембране Н
+
-АТФаза работает в обратном
направлении, используя ΔμН
+
, создаваемый в дыхательной цепи, для образования
АТФ.
Наконец, в клетках широко представлен вторично-активный транспорт, в процессе
которого градиент одного вещества используется для транспорта другого. С по-
мощью вторично-активного транспорта клетки аккумулируют сахара, аминокисло-
ты и выводят некоторые продукты метаболизма, используя градиент Na
+
, созда-
ваемый в ходе работы Na
+
/К
+
-АТФазы (см. рис. 9.5).
БИОЭНЕРГЕТИКА
С позиций термодинамики (см. главу 4) метаболизм представляет собой
совокупность процессов, в которой реакции, потребляющие энергию из
внешней среды (эндэргонические), сопрягаются с энергодающими (экзэрго-
ническими) реакциями, что позволяет живым существам оказывать посто-
янное сопротивление нарастанию энтропии. Выяснение биохимических
механизмов, приводящих к генерации различных форм биологической
энергии, является предметом биоэнергетики. Источником энергии служат
реакции, в ходе которых соединения, содержащие атомы углерода в высо-
ковосстановленном состоянии, подвергаются окислению, а специальные
дыхательные переносчики присоединяют протоны и электроны (восстанав-
ливаются) и в таком виде транспортируют атомы водорода к дыхательной
цепи.
Биологические виды энергии. Энергетические превращения в живой клет-
ке подразделяют на две группы: локализованные в мембранах и протекаю-
щие в цитоплазме. В каждом случае для «оплаты» энергетических затрат
используется своя «валюта»: в мембране это ΔμН
+
или ΔμNa
+
, а в цито-
плазме – АТФ, креатинфосфат и другие макроэргические соединения. Не-
посредственным источником АТФ являются процессы субстратного и окис-
лительного фосфорилирования. Процессы субстратного фосфорилирования
наблюдаются при гликолизе и на одной из стадий цикла трикарбоновых
кислот (реакция сукцинил-КоА —> сукцинат; см. главу 10). Генерация ΔμН
+
и ΔμNa, используемых для окислительного фосфорилирования, осуществ-
ляется в процессе транспорта электронов в дыхательной цепи энергосопря-
гающих мембран.
Энергия разности потенциалов на сопрягающих мембранах может
обратимо превращаться в энергию АТФ. Эти процессы катализируются
Н
+
-АТФ-синтазой в мембранах, генерирующих протонный потенциал, или
Na
+
-АТФ-синтазой (Na
+
-АТФазой) в «натриевых мембранах» алкалофиль-
ных бактерий, поддерживающих ΔμNa
+
[Скулачев В.П., 1989]. На рис. 9.6
представлена схема энергетики живых клеток, использующих ΔμН
+
в каче-
стве мембранной формы конвертируемой энергии. На схеме видно, что свет
или энергия субстратов дыхания утилизируется ферментами фотосинтети-
ческой или дыхательной редокс-цепи (у галобактерий – бактериородопси-
ном). Генерируемый потенциал используется для совершения полезной
работы, в частности для образования АТФ. Будучи макроэргическим
соединением, АТФ выполнняет функцию аккумулирования биологической
энергии и ее последующего использования для выполнения клеточных
функций. «Макроэргичность» АТФ объясняется рядом особенностей его
305
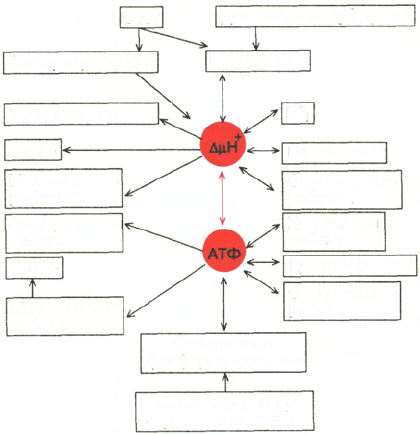
Рис. 9.6. Взаимозаменяемость различных видов биологической энергии при выпол-
нении клеточной работы [Скулачев В.П., 1989].
Красной стрелкой показана взаимозаменяемость в клетке двух клеточных видов энергии - АТФ
и ΔμН
+
, для которых имеются также специальные буферные системы: креатинфосфат для АТФ
(клетки животных) и градиент ионов Na (алкалофильные бактерии).
молекулы. Это прежде всего высокая плотность зарядов, сконцентрирован-
ная в «хвосте» молекулы, обеспечивающая легкость диссоциации терми-
нального фосфата при водном гидролизе. Продукты этого гидролиза
представляют собой АДФ и неорганический фосфат и далее – АМФ и неор-
ганический фосфат. Это обеспечивает высокую величину свободной энергии
гидролиза терминального фосфата АТФ в водной среде.
Тканевое дыхание и биологическое окисление. Распад органических соеди-
нений в живых тканях, сопровождающийся потреблением молекулярного
кислорода и приводящий к выделению углекислого газа и воды и образова-
нию биологических видов энергии, называется тканевым дыханием. Ткане-
вое дыхание представляют как конечный этап пути превращений моносаха-
ров (в основном глюкозы) до указанных конечных продуктов, в который на
разных стадиях включаются другие сахара и их производные, а также
промежуточные продукты распада липидов (жирные кислоты), белков
(аминокислоты) и нуклеиновых оснований. Итоговая реакция тканевого
дыхания будет выглядеть следующим образом:
С
6
Н
12
О
6
+ 6O
2
= 6СO
2
+ 6Н
2
O + 2780 кДж/моль.
(1)
Впервые сущность дыхания объяснил А.-Л. Лавуазье (1743-1794), обративший
внимание на сходство между горением органических веществ вне организма и дыха-
нием животных. Постепенно становились ясными принципиальные различия между
этими двумя процессами: в организме окисление протекает при относительно
низкой температуре в присутствии воды, и его скорость регулируется обменом
веществ. В настоящее время биологическое окисление определяется как совокупность
реакций окисления субстратов в живых клетках, основная функция которых - энерге-
306
Свет
Дыхательные субстраты
Бактериородопсин
Редокс-цепи
Трансгидрогеназа
РР
i
Тепло
Механическая
работа
Электрическая
работа
Тепло
Механическая
работа
Na-градиент
Осмотическая
работа
Химическая
работа
Креатинфосфат
Осмотическая
работа
Субстратное
фосфорилирование
Гликолитические суб-
страты, кетокислоты
тическое обеспечение метаболизма. В развитие концепций биологического окисле-
ния в XX в. важнейший вклад внесли А.Н. Бах, О. Варбург, Г. Крепс, В.А. Энгель-
гардт, В.И. Палладин, В.А. Белицер, С.Е. Северин, В.П. Скулачев.
Потребление кислорода тканями зависит от интенсивности реакций
тканевого дыхания. Наибольшей скоростью тканевого дыхания характери-
зуются почки, мозг, печень, наименьшей – кожа, мышечная ткань (в покое).
Уравнение (2) описывает суммарный результат многоступенчатого процес-
са, приводящего к образованию молочной кислоты (см. главу 10) и проте-
кающего без участия кислорода:
С
6
Н
12
О
б
= 2С
3
Н
6
О
3
+ 65 кДж/моль.
(2)
Этот путь отражает, по-видимому, энергетическое обеспечение простей-
ших форм жизни, функционировавших в бескислородных условиях. Совре-
менные анаэробные микроорганизмы (осуществляющие молочнокислое,
спиртовое и уксуснокислое брожение) получают для жизнедеятельности
энергию, производимую в процессе гликолиза или его модификаций.
Использование клетками кислорода открывает возможности для более
полного окисления субстратов. В аэробных условиях продукты бескисло-
родного окисления становятся субстратами цикла трикарбоновых кислот
(см. главу 10), в ходе которого образуются восстановленные дыхательные
переносчики НАДФН, НАДН и флавиновые коферменты. Способность
НАД
+
и НАДФ
+
играть роль промежуточного переносчика водорода
связана с наличием в их структуре амида никотиновой кислоты. При
взаимодействии этих кофакторов с атомами водорода имеет место обрати-
мое гидрирование (присоединение атомов водорода):
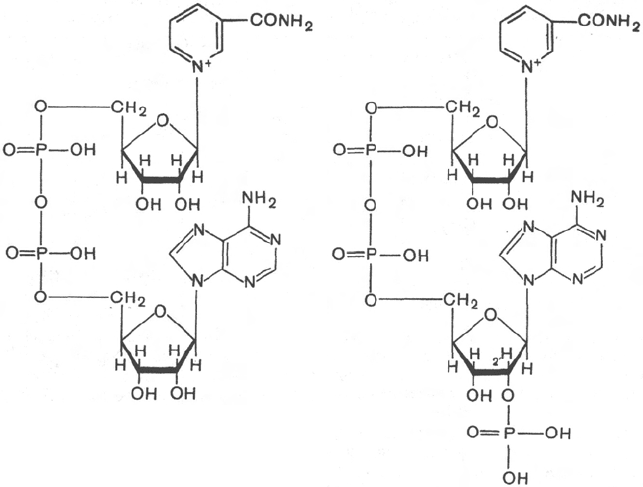
307
Никотинамидадениндинуклеотид
(НАД
+
)
Нинотинамидадениндинуклеотид-
фосфат
(НАДФ
+
)
При этом в молекулу НАД
+
(НАДФ
+
) включаются 2 электрона и один
протон, а второй протон остается в среде.
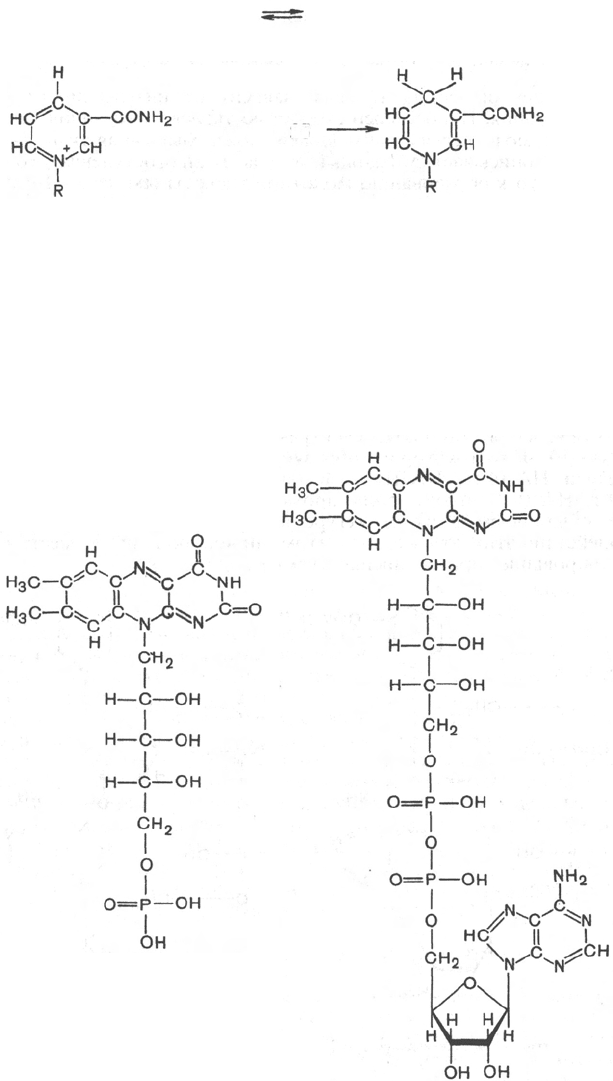
Во флавиновых коферментах (ФАД или ФМН), активной частью моле-
кул которых является изоаллоксазиновое кольцо, в результате восстанов-
ления чаще всего наблюдается присоединение 2 протонов и 2 электронов
одновременно:
308
Флавинмононуклеотид
(ФМН)
Флавинадениндинуклеотид
(ФАД)
Субстрат–Н
2
+ НАД(Ф)
Субстрат + НАД(Ф)Н + Н
+
НАД
+
НАДН
+ 2Н
+
+
+ H
+
2e
–
Восстановленные формы этих кофакторов способны транспортировать
водород и электроны к дыхательной цепи митохондрий или иных энерго-
сопрягающих мембран (см. далее).
Организация и функционирование дыхательной цепи. В клетках эукариот
дыхательная цепь расположена во внутренней мембране митохондрий,
у дышащих бактерий – в цитоплазматической мембране и специализирован-
ных структурах – мезосомах, или тилакоидах. Компоненты дыхательной
цепи митохондрий в порядке убывания окислительно-восстановительного
потенциала можно расположить, как показано в табл. 9.1.
Таблица 9.1. Окислительно-восстановительный потенциал компонентов дыхатель-
ной цепи в стандартных условиях (концентрация компонентов 1М, рН 7,25°С)
Восстановленная форма
НАДН + Н
+
ФАДН
2
Убихинол (KoQ-H
2
)
Цитохром b (Fe
2+
)
» с
1
(Fe
2+
)
» с (Fe
2+
)
» a (Fe
2+
)
» а
3
(Fe
2+
)
H
2
O
Окисленная форма
НАД
+
ФАД
+
Убихинон
Цитохром b (Fe
3+
)
» c
1
(Fe
3+
)
» с (Fe
3+
)
» a (Fe
3+
)
a
3
(Fe
3+
)
1
/
2
O
2
E°, В
–0,32
–0,05
+
0,04
+
0,07
+
0,23
+
0,25
+
0,29
+0,55
+
0,82
Молярные соотношения компонентов дыхательной цепи являются постоянными,
ее компоненты встроены в митохондриальную мембрану в виде 4 белково-липидных
комплексов: НАДН-КоQН
2
-редуктаза (комплекс I), сукцинат-КоQ-редуктаза (комп-
лекс II), КоQН
2
-цитохром c-редуктаза (комплекс III) и цитохром а-цитохромокси-
даза (комплекс IV) (рис. 9.7).
Если субстратом окисления служат α-кетокислоты, в переносе электронов на
НАД
+
участвуют липоатсодержащие дегидрогеназы. В случае окисления пролина,
глутамата, изоцитрата и других субстратов перенос электронов происходит непос-
редственно на НАД
+
. Восстановленный НАД в дыхательной цепи окисляется
НАДН-дегидрогеназой, содержащей железосерный белок (FeS) и ФМН и прочно
связанной с дыхательной цепью.
309
ФАД
(или ФМН)
ФАДН
2
(или ФМНН
2
)
НАДН + Н
+
+
Флавиновый фермент
(ФМН)
НАД
+
+
Флавиновый фермент
(ФМНН
2
)
+ 2Н
+
+ 2e
–

Рис. 9.7. Взаимное расположение компонентов дыхательной цепи с указанием мест
фосфорилирования и специфических ингибиторов.
KoQ (убихинон), необходимый компонент дыхательной цепи, является производ-
ным бензохинона с боковой цепью, которая у млекопитающих чаще всего представ-
лена 10 изопреноидными единицами (см. главу 7). Как любой хинон, KoQ способен
находиться и в восстановленном, и окисленном состоянии. Это свойство определяет
его роль в дыхательной цепи - служить коллектором восстановительных эквивален-
тов, поставляемых в дыхательную цепь через флавиновые дегидрогеназы. Содержа-
ние его значительно превосходит содержание других компонентов дыхательной
цепи.
Дополнительным участником дыхательной цепи является железосерный белок
FeS (негемовое железо). Он участвует в окислительно-восстановительном процессе,
протекающем по одноэлектронному типу. Первый участок локализации FeS нахо-
дится между ФМН и KoQ, второй - между цитохромами b и c
1
. Это соответствует
тому факту, что со стадии ФМН путь протонов и электронов разделяется: первые
накапливаются в митохондриальном матриксе, а вторые идут на гидрофобные
переносчики - KoQ и цитохромы.
Цитохромы в дыхательной цепи выстроены в порядке возрастания окислительно-
восстановительного потенциала. Они представляют собой гемопротеины, в которых
простетическая геминовая группа близка к гему гемоглобина (у цитохрома b иден-
тична). Ионы железа в составе гема при получении и отдаче электронов обратимо
изменяют свою валентность.
В процессах тканевого дыхания наиболее важную роль играют цитохро-
мы b, с
1
, с, а и а
3
. Цитохром а
3
представляет собой терминальный участок
дыхательной цепи – цитохромоксидазу, которая осуществляет окисление
цитохрома с и образование воды. Элементарный акт представляет собой
двухэлектронное восстановление одного атома кислорода, т.е. каждая
молекула кислорода одновременно взаимодействует с двумя электрон-
транспортными цепями. При транспорте каждой пары электронов во
внутримитохондриальном пространстве может накапливаться до 6 прото-
нов
(рис. 9.8).
Строение дыхательной цепи интенсивно исследуется. В числе послед-
них достижений молекулярной биохимии – установление тонкой структу-
310
Место сопряжения 1
Место сопряжения 2
Место сопряжения 3
Малонат
Сукцинат
Комплекс II
НАДН
Комплекс I
ФМН,
FеS
ФАД
FeS
Карбоксин
Антимицин А
H
2
S
CO
CN
-
Комплекс IV
Комплекс III
Q
Цит b,
FeS,
Цит с
1
Цит с
Цит а
Сu
Цитa
3
Сu
O
2
Амобарбитал
Ротенон
АДФ + Р
i
АТФ
АДФ + Р
i
АТФ
АДФ + P
i
АТФ