Белясова Н.А. Биохимия и молекулярная биология
Подождите немного. Документ загружается.

190
расходуются восстановительные эквиваленты и ATP, а фиксированный в
цикле Кальвина углерод бесполезно теряется в виде СО
2
. Показано, что фото-
дыхание может приводить к реокислению и выделению до 50% углерода,
фиксированного в цикле Кальвина. До сих пор не разгадана целесообразность
этого столь неэкономного и ограничивающего эффективность роста растений
процесса.
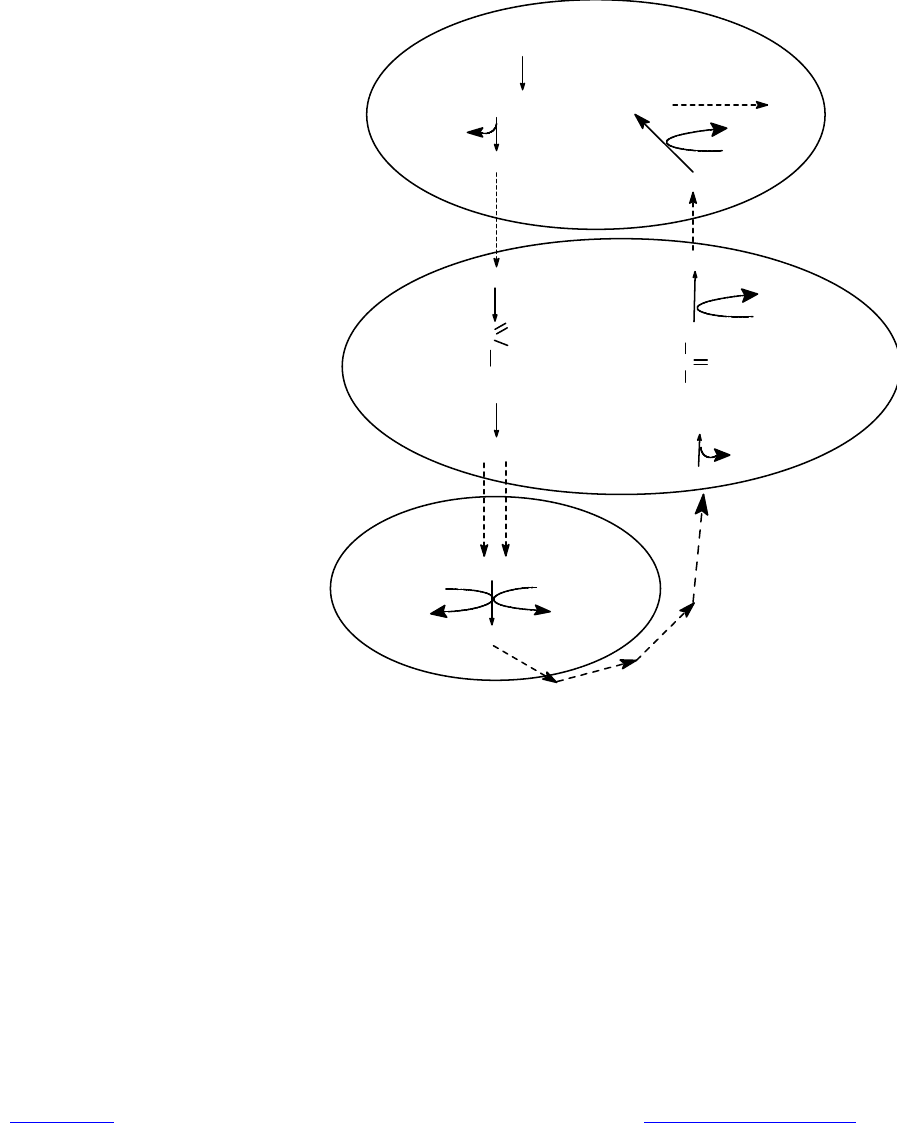
Последовательность событий фотодыхания изображена на рис. 13.7.
Можно видеть, что отдельные стадии этого процесса осуществляются в раз-
ных органеллах. Итогом фотодыхания является образование одной
Рибулозодифосфат + О
2
Фосфогликолат + 3-Фосфоглицерат
Гликолат
Гликолат
+ О
2
P
i
C
COOH
Глиоксилат
+ NH
3
Глицин
Глицин + Глицин
1/2 O
2
CO
2
ADP
ATP
Серин + NH
3
Серин
COOH
C
CH
2
OH
O
NH
3
Гидроксипируват
NADH
NAD
+
Глицерат
Глицерат
ATP
ADP
Цикл
Кальвина
Хлоро-
пласт
Перокси-
сома
Мито-
хондрия
O
H
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

191
Рис. 13.7. Фотодыхание
молекулы фосфоглицерата (3 атома «С») из двух молекул гликолата (4 атома
«С»), а участие кислорода приводит к окислению четвертого атома углерода в
углекислоту. По-видимому, назначение сложной последовательности превра-
щений в фотодыхании состоит в том, чтобы вернуть в цикл хотя бы часть углеро-
да из гликолата, который накапливается в избытке.
При фотодыхании происходит потеря одного атома углерода из каждых
четырех, причем следует учитывать, что на фиксацию этого атома углерода
уже затрачена энергия в цикле Кальвина. Кроме этого, при окислении глици-
на и дезаминировании серина выделяется аммиак, на включение которого в
состав аминокислот снова придется затрачивать энергию. В результате фото-
дыхание снижает потенциальную урожайность С
3
-растений на 30—40%.
13.2. Зрительное восприятие
Органы зрения представляют собой высокоразвитые рецепторы, в основе
функционирования которых лежат фотохимические реакции светочувстви-
тельных пигментов (фоторецепторов). Для улавливания наибольшего количе-
ства света фоторецепторами светочувствительные клетки глаза (палочки и
колбочки сетчатки) устроены особым образом: их внешние сегменты бук-
вально «напичканы» специализированными мембранами, в которых и нахо-
дятся рецепторные молекулы (рис. 13.8). Каждая такая клетка соединяется с
помощью посредника с аксонами зрительного нерва, в мембранах которых в
результате каскадных реакций в ответ на фотохимические превращения фо-
торецепторов возбуждаются нервные импульсы. Таким образом, свет различ-
ной длины волны, отражаемый объектами, расположенными на различном
расстоянии, воспринимается глазом и преобразуется в нервные импульсы. По
аксонам зрительного нерва эти импульсы поступают в особый отдел головно-
го мозга (зрительную кору) и интерпретируются там, порождая изображение.
В палочках (высокочувствительные клетки сетчатки, отвечающие за
«черно-белое» зрение), в их внешнем, обращенном к окружающей среде сег-
менте, располагается порядка 500 параллельно уложенных дисков диаметром
~ 2 мкм (рис. 13.8). Каждый диск образован парой мембран, разделенных
узким пространством. Мембраны на 60% состоят из белка, основная часть
которого приходится на родопсин — светочувст-вительный хромопротеин.
Хромофором родопсина служит 11-цис-ретиналь, он через e-аминогруппу
остатка лизина связан с белковой частью — опсином (рис. 13.8). Родопсин
присутствует в мембранах дисков в виде трансмембранного белка.
При поглощении молекулой родопсина кванта света происходит изомери-
зация 11-цис-ретиналя в полностью транс-ретиналь (рис. 13.8). В результате
этой фотохимической реакции родопсин переходит в активную форму и сти-
мулирует G-белок, циркулирующий в цитоплазме палочек. В свою очередь,
G-белок запускает каскад передачи сигнала, который и приводит к генериро-
ванию нервного импульса в мембранах нервных клеток.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
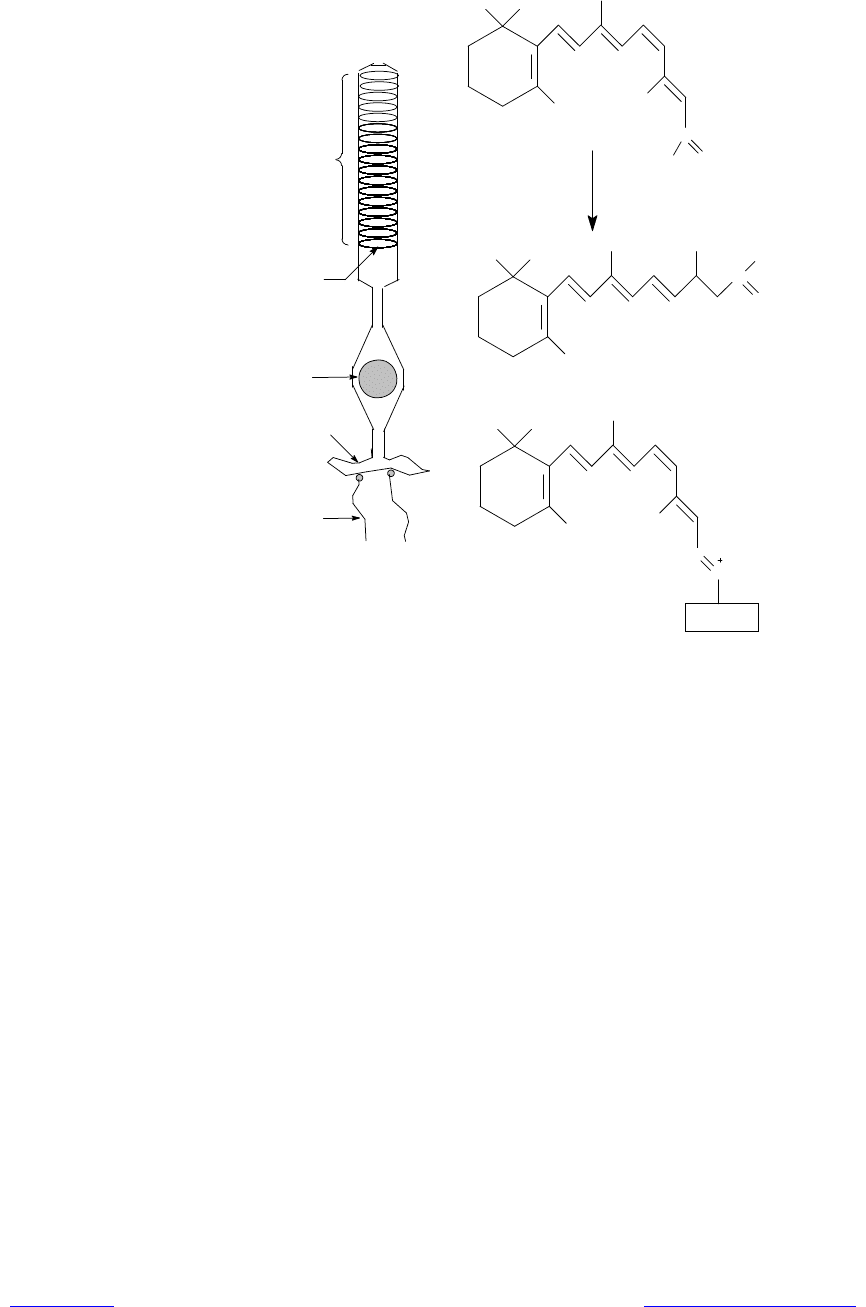
192
Свет
Наружный
сегмент
Мемб-
ранные
диски
Синапс
Отростки
нервных
клеток
Палочка сетчатки глаза
ядро
CH
3
H
3
C
CH
3
C
CH
3
H
3
C
H
O
11-цис-ретиналь
hn
CH
3
CH
3
H
3
C
CH
3
C
CH
3
H
O
11
12
12
11
Полностью транс-ретиналь
12
11
H
3
C
CH
3
CH
3
H
3
C
CH
3
HC
NH
опсин
Связанная с белком
форма цис-ретиналя
Рис. 13.8. Схематическое строение палочки. Родопсин. Изомеризация ретина-
ля из цис-формы в транс-форму
Транс-ретиналь характеризуется низким сродством к опсину, и спустя ко-
роткое время активированный родопсин диссоциирует на опсин и полностью
транс-ретиналь. Специальная ретиналь-изомераза катализирует превращение
транс-ретиналя в его цис-форму, после чего происходит самопроизвольная
ассоциация цис-ретиналя с опсином, обусловленная высоким сродством этих
структурных частей друг к другу, и снова образуется родопсин. Все перечис-
ленные события циклически повторяются и известны под названием зри-
тельный цикл (рис. 13.9).
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
193
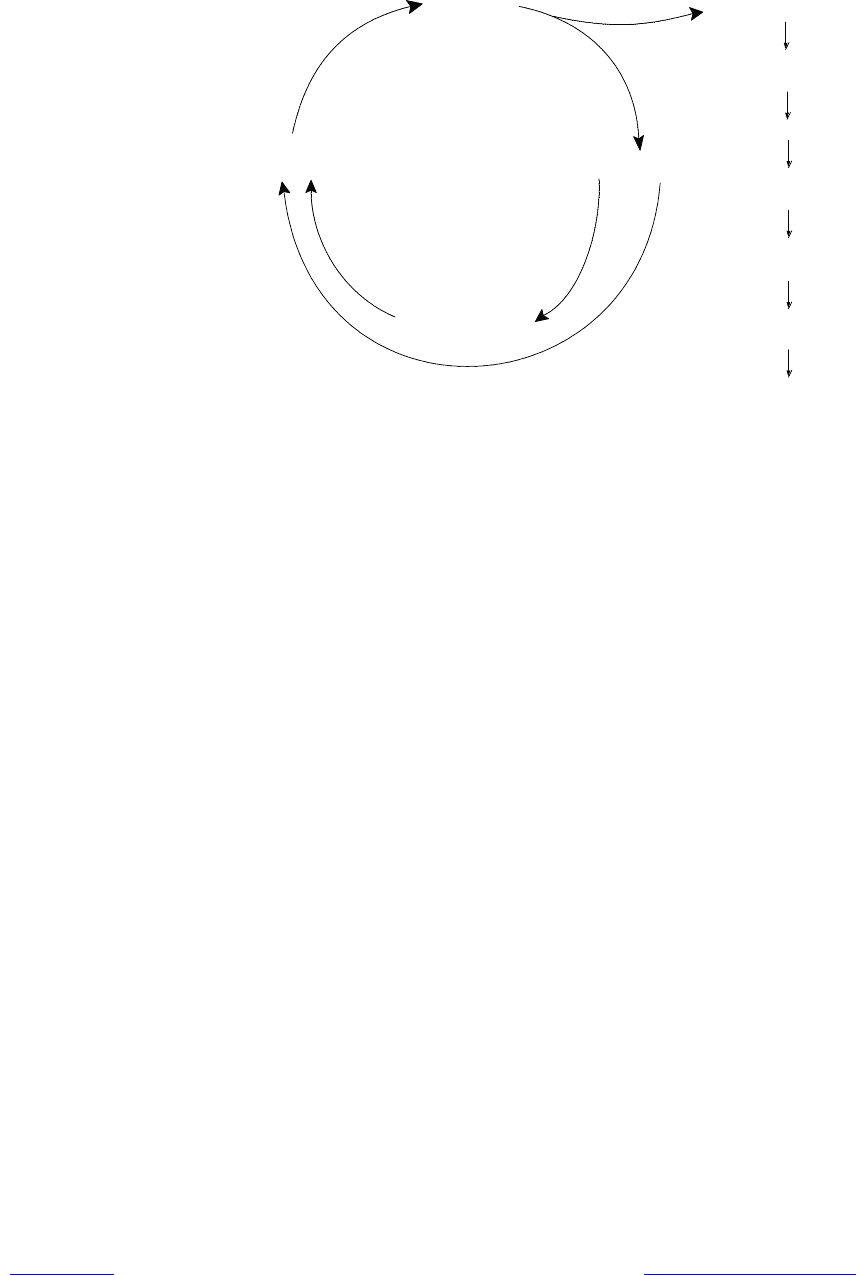
Для понимания механизма генерирования нервного импульса в ходе кас-
кадных событий, вызванных изомеризацией ретиналя под действием
Родопсин
( )
Родопсин*
(транс ретиналь)
Транс ретиналь опсин
Стимуляция G белка
hn
Ретиналь
изомераза
Активация GMP
фосфодиэстеразы
Падение уровня cGMP
Закрывание катионных
каналов
Гиперполяризация
мембраны палочки
Остановка выброса
нейромедиатора
Генерация нервного
импульса в мембранах
нейронов
11-цис-ретиналь
11-цис-ретиналь
-
-
-
-
-
+
Рис. 13.9. Зрительный цикл и связанные со световой активацией родопсина
каскадные реакции, приводящие к генерации нервного импульса в мембранах
нейронов (родопсин* — активированная форма родопсина;
cGMP- циклический гуанозинмонофосфат)
света, следует учитывать некоторые особенности функционирования свето-
чувствительных клеток. В мембране палочки находятся катионные насосы,
которые постоянно выкачивают ионы Na
+
и Ca
2+
из клетки, создавая на ней
ионный градиент. В темноте внутрь палочек постоянно течет поток ионов
натрия и кальция, которые пользуются cGMP-зависимыми каналами для воз-
вращения в клетку согласно закономерностям облегченной диффузии. Эти
каналы остаются открытыми при связывании с ними циклического гуанозин-
монофосфата (cGMP).
При освещении происходят следующие события. Активный родопсин, в
составе которого цис-ретиналь подвергнулся фотохимическому превращению
в полностью транс-ретиналь, связывается с G-белком. При этом в составе G-
белка GDP обменивается на GTP и белок диссоциирует на субъединицы, одна
из которых представляет собой активную GTP-a-субъединицу. Эта структура
активирует фермент сGMP-фосфодиэстеразу, которая катализирует гидролиз
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

194
циклического гуанозинмонофосфата. Уровень сGMP быстро понижается, что
приводит к перекрыванию катионных каналов. В результате концентрация
ионов Na
+
и Ca
2+
на внутренней поверхности мембраны палочки резко падает,
поскольку продолжают работать катионные насосы: происходит гиперполя-
ризация клетки. Гиперполяризация замедляет высвобождение из палочек воз-
буждающего нейромедиатора (глутамата), и в ответ на это опосредованным
образом в аксонах зрительного нерва возникает потенциал действия (гла-
ва 4).
Активная GTP-a-субъединица G-белка существует очень непродолжи-
тельное время и инактивируется за счет гидролиза GTP. Образуется свобод-
ная a-субъединица, ассоциирующая с остальными субъединицами с образо-
ванием G-белка, способного снова перейти в активную форму за счет связы-
вания с активным родопсином.
Бактериородопсин. Белок, похожий по структуре на родопсин глаза
(бактериородопсин), обнаружен в клетках некоторых архебактерий, где он
тоже используется для фоторецепции. Высокоспециализированная в физиоло-
гическом отношении группа галобактерий (Halobacterium, Halococcus, Natro-
coccus, Natrobacterium) содержит необычные пурпурные мембраны, окра-
шенные в ярко оранжевые — ярко красные тона за счет содержания в них
бактериородопсина. Эти бактерии уникальны тем, что способны за счет по-
глощения энергии света генерировать на мембране протонный градиент, ко-
торый используется в качестве движущей силы при синтезе АТР. Другой от-
личительной особенностью галобактерий является способность развиваться в
концентрированных растворах солей (галофилия).
Бактериородопсин представляет собой интегральный белок пурпурной
мембраны, в которой он формирует полый цилиндр. Хромофором бактерио-
родопсина служит ретиналь. В темноте ретиналь находится в полностью
транс-форме и его альдиминная группа (= N
+
H -) протонирована (рис. 13.8).
При освещении ретиналь перегруппировывается в 13-цис-форму, а альди-
минная группа отдает протон, который через канал выводится на наружную
поверхность мембраны. Так на мембране создается протонный градиент. За-
кономерности синтеза АТР за счет энергии протонного градиента такие же,
как при окислительном фосфорилировании. Таким образом, бактериородоп-
син в пурпурных мембранах выполняет роль протонного насоса, приводимого
в действие светом.
Энергия, запасенная в ходе такого необычного способа «фотосинтеза»,
дополняет энергию, которую галобактерии запасают в ходе дыхания.
13.3. Биолюминесценция
Под биолюминесценцией понимают способность живых организмов ис-
пускать видимый свет. В основе данного явления лежит ферментативное
аэробное окисление веществ особого семейства, называемых люциферина-
ми. Катализирующие этот процесс ферменты называют люциферазами.
Энергия, выделяющаяся в процессе окисления люциферинов, превращается в
энергию электронного возбуждения молекул, способных испускать ее в виде
фотонов. Свечение может иметь самые разные тона: от голубого до красного.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

195
К биолюминесценции способны многие организмы: бактерии, грибы, про-
стейшие, моллюски, насекомые (светляки), кишечнополостные животные,
ракообразные, рыбы и др. Люциферины этих организмов сильно различаются
по структуре и свойствам. Большинство из них испускают свет при окисле-
нии, взаимодействуя с О
2
. Однако некоторые люциферины, например такие,
как фотобелок медуз, активируются ионами кальция.
У светящихся бактерий роль люциферина выполняет FMNH
2
. При его
окислении в присутствии альдегида с длинной цепью (не менее 8 атомов уг-
лерода) происходит одновременное окисление альдегида в карбоновую ки-
слоту и люциферина в FMN:
FMNH
2
+ O
2
+ R—CHO + люцифераза ®
® FMN + H
2
O + R—COOH + фотон
Эта реакция протекает, видимо, через стадию образования пероксида фла-
винмононуклеотида, а свечение характеризуется длиной волны ~490 нм.
Светящиеся организмы могут использовать явление биолюминесценции
для опознавания особей своего вида, приманки добычи, в качестве брачных
сигналов, отпугивания хищников и отвлечения их внимания, для «высвечи-
вания» жертвы и т. д. Менее понятна роль биолюминесценции для низкоор-
ганизованных организмов, например бактерий.
Существует точка зрения, согласно которой биолюминесценция возникла
на стадии перехода от анаэробных форм жизни к аэробным как защитная
реакция по отношению к мощному окислителю — молеку-лярному кислоро-
ду.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

196
Часть четвертая. МЕТАБОЛИЗМ.
ПРОЦЕССЫ, ТРЕБУЮЩИЕ ПРИТОКА ЭНЕРГИИ
Глава 14. ОСОБЕННОСТИ БИОСИНТЕЗА.
БИОСИНТЕЗ УГЛЕВОДОВ
Биосинтез клеточных веществ составляет одну из основных «затратных
статей» метаболизма: на эти нужды расходуется самая большая доля энергии,
запасенной в составе АТР, восстановительных эквивалентов и строительных
блоков. Биосинтетические реакции относят к анаболизму, особенности кото-
рого и взаимосвязь с другими гранями метаболизма описана в главе 8. К этой
информации стоит добавить еще несколько важных закономерностей биосин-
тетических процессов: 1) пути биосинтеза сложных клеточных соединений
часто осуществляются параллельно соответствующим путям катаболизма, в
которых формируются строительные блоки — предшественники биосинтеза.
Однако последовательности реакций в этих взаимообращенных путях нико-
гда полностью не совпадают, так же как не совпадают наборы ферментов,
принимающих участие в этих процессах; 2) регуляция путей биосинтеза и
катаболитных процессов осуществляется независимо, на уровне разных фер-
ментов; 3) вступление веществ в пути биосинтеза требует их активации, что
чаще всего осуществляется в ходе фосфорилирования при участии АТР;
4) процессы биосинтеза родственных по структуре и функциям соединений
часто представляют собой «разветвленные» пути, в которых продукт одной
серии реакций становится субстратом для синтеза других продуктов; 5) роль
предшественников в биосинтезе большинства клеточных веществ выполняет
ограниченное количество субстратов, которые образуются в ходе основных
катаболических и анаболических процессов и принадлежат к ключевым
промежуточным соединениям клетки.
Большинство перечисленных закономерностей биосинтеза можно обна-
ружить, например, при сравнении процессов гликолиза и глюконеогенеза.
Последний будет рассмотрен в данной главе.
14.1. Закономерности обмена и биосинтеза моносахаридов
Ключевым промежуточным соединением в обмене углеводов можно на-
звать глюкозо-6-фосфат. Это вещество может образовываться из экзогенной
глюкозы при ее фосфорилировании в процессе транспорта через мембраны
или сразу после этого. Кроме того, глюкозо-6-фосфат образуется в клетках
автотрофных организмов, способных к фиксации СО
2
, в процессе глюконео-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

197
генеза из глицеральдегид-3-фосфата (рис. 13.5). Другими источниками глю-
козо-6-фосфата являются: пируват (образуется в результате большинства ка-
таболитных путей расщепления углеводов), некоторые промежуточные про-
дукты ЦТК, углеродные скелеты гликогенных аминокислот, лактат (продукт
молочнокислого брожения), глицерол (образуется при расщеплении липидов).
Все эти вещества могут вовлекаться в процесс глюконеогенеза — синтеза
глюкозы из неуглеводных предшественников. Кроме этого, в клетках расте-
ний и многих микроорганизмов глюкоза может образовываться из ацетил-
СоА — продук-та расщепления жирных кислот. В этом случае используются
два специфических фермента глиоксилатного цикла, отсутствующие в клет-
ках высших животных. Эти ферменты (изоцитрат-лиаза и малат-синтаза)
участвуют в превращении ацетил-СоА в сукцинат, который затем преобразу-
ется в оксалоацетат (рис. 11.7) и вступает в процесс глюконеогенеза. Нако-
нец, еще одним источником глюкозы, а значит, и глюкозо-6-фосфата являют-
ся резервные полисахариды.
Расходуется глюкозо-6-фосфат в клетках следующим образом. Во-первых,
он является субстратом разнообразных катаболических путей, в которых за-
пасается энергия, образуются восстановительные эквиваленты и строитель-
ные блоки (глава 9). Во-вторых, глюкозо-6-фосфат способен изомеризоваться
в глюкозо-1-фосфат и служить субстратом для биосинтеза гликогена либо
иных запасных и структурных полисахаридов. Глюкозо-6-фосфат может де-
фосфорилироваться с образованием глюкозы, которая способна трансформи-
роваться в клетках в другие моносахариды, например галактозу, маннозу,
фруктозу, глюкуроновую кислоту. Названные соединения принимают участие
в процессах образования сложных полисахаридов клеточных стенок или дру-
гих структур.
Большинство перечисленных выше способов образования и расходования
моносахаридов уже охарактеризовано в предыдущих разделах. Здесь внима-
ние будет уделено одному из центральных путей образования ключевого про-
дукта углеводного обмена (глюкозо-6-фосфата) —глюконеогенезу.
Глюконеогенез. Этот процесс характерен для представителей всех царств
живых организмов, но наиболее важное значение имеет для клеток высших
животных. Дело в том, что эмбриональные ткани, мозг, семенники, эритро-
циты в качестве источника углерода способны использовать только D-
глюкозу. Если в рационе недостает углеводов, в печени индуцируется распад
гликогена, но и этого источника может оказаться недостаточно (мозг челове-
ка в сутки потребляет более 120 г глюкозы). В таком случае глюкоза синтези-
руется в организме из неуглеводных предшественников в ходе глюконеогене-
за. Наиболее активно глюконеогенез осуществляется у животных в клетках
печени и почек.
Реакции глюконеогенеза в большой степени тождественны обратным ре-
акциям гликолиза, и многие из них катализируются теми же ферментами,
которые задействованы в гликолизе (риc. 14.1).
Итак, в гликолизе имеется три практически необратимые реакции, взамен
которых в глюконеогенезе существуют обходные пути. Первый обходной
путь представляет собой превращение пирувата в фосфоенол-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
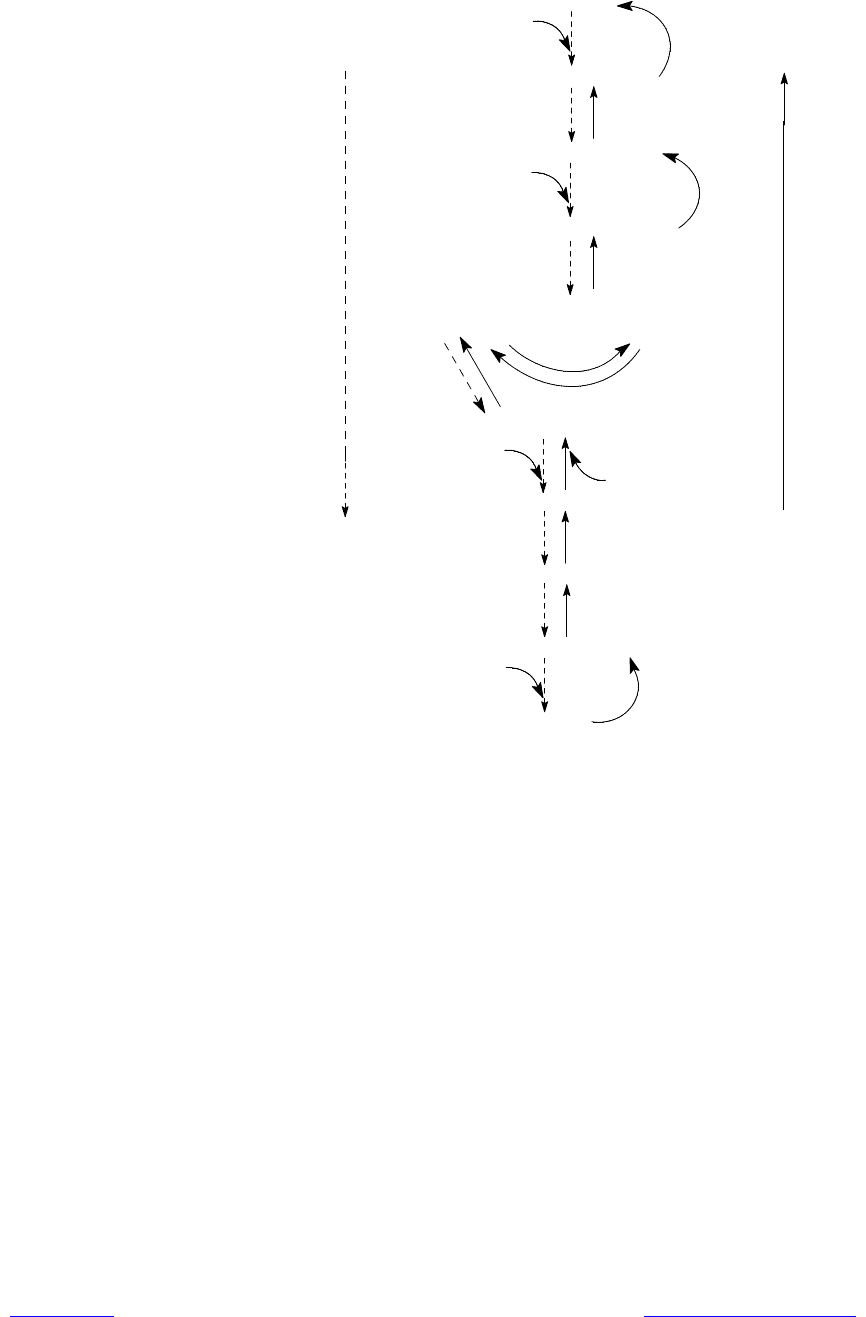
198
Г
л
ю
к
о
з
а
Глюкозо-6-фосфат
Фруктозо-6-фосфат
Фруктозо-1,6-дифосфат
Глицеральдегид-
-3-фосфат
Дигидрокси-
ацетонфосфат
1,3-Дифосфоглицерат
3-Фосфоглицерат
2-Фосфоглицерат
Фосфоенолпируват
Пируват
Глюкозофос-
фатизомераза
Фруктозодифос-
фатальдолаза
Глицераль-
дегид-3-фос-
фатдегид-
рогеназа
Триозофосфат-
изомераза
Фосфоглице-
раткиназа
Фосфоглице-
ратмутаза
Енолаза
Обходной
путь
ADP
+
АТР
АТР
АТР
ADP
Обходной
путь
Обходной
путь
Г
л
и
к
о
л
и
з
Г
л
ю
к
о
н
е
о
г
е
н
е
з
Рис. 14.1. Последовательность реакций гликолиза (пунктирные стрелки) и
глюконеогенеза (сплошные стрелки). Подписаны названия ферментов, общих
для двух путей. Стадии, по которым процессы различаются, обо-
значены в глюконеогенезе, как обходные пути
пируват. Для непосредственного перевода пирувата в фосфоенолпируват не-
достаточно энергии расщепления АТР, поэтому данная стадия осуществляет-
ся в ходе нескольких реакций. Вначале пируват, образующийся преимущест-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

199
венно в цитоплазме (из лактата, аминокислот, в гликолизе), переводится в
митохондрии и там карбоксилируется в оксалоацетат (рис. 14.2). Катализиру-
ет реакцию пируваткарбоксилаза, использующая в качестве кофактора био-
тин. Эта анаплеротическая реакция и характеристика свойств фермента опи-
саны в разделе 11.2.
Оксалоацетат в митохондриях восстанавливается в малат (митохондри-
альная малатдегидрогеназа), который с помощью специфических переносчи-
ков транспортируется в цитоплазму. В цитоплазме малат вновь окисляется в
оксалоацетат (цитоплазматическая малатдегидрогеназа), который с помощью
GTP-зависимой фосфоенолпируваткарбоксилазы декарбоксилируется в фос-
фоенолпируват (РEP).
Второй обходной путь в глюконеогенезе представляет собой превращение
фруктозодифосфата во вруктозо-6-фосфат (рис. 14.2). В гликолизе фосфоф-
руктокиназная реакция, сопровождающаяся гидролизом АТР, является необ-
ратимой. В глюконеогенезе функционирует другой фермент —
фруктозодифосфатаза, которая катализирует практически необратимое от-
щепление фосфатной группы от первого атома углерода. Фруктозодифосфа-
таза, как и пируваткарбоксилаза, является аллостерическим ферментом. Его
активность ингибируется с помощью АМР и активируется при участии АТР.
Третий обходной путь — дефосфорилирование глюкозо-6-фосфата, не
может произойти с помощью прямого обращения гексокиназной реакции. Эту
реакцию катализирует глюкозо-6-фосфатаза, которая локализована на внут-
ренней поверхности мембран гладкого эндоплазматического ретикулума
(ЭР). Поэтому для осуществления данной реакции глюкозо-6-фосфат транс-
портируется в ЭР, где дефосфорилируется в свободную глюкозу (рис. 14.2).
Следует отметить, что глюкозо-6-фосфатаза отсутствует в таких тканях, как
мышцы и мозг, поэтому они не могут поставлять в кровь свободную глюкозу.
Суммарное уравнение глюконеогенеза выглядит следующим образом:
2Пируват + 4АТР + 2GTP +2NADH + 2H
+
+ 4H
2
O ®
® Глюкоза + 2NAD
+
+ 4ADP + 2GDP + 6 P
i
Из приведенного баланса следует, что на образование одной молекулы
глюкозы в процессе глюконеогенеза расходуется шесть высокоэнергетиче-
ских фосфатных связей, а также две молекулы NADH. Важно отметить, что
регуляция скорости синтеза глюкозы в этом пути осуществляется с помощью
ферментов, не принимающих участие в гликолизе. При этом глюконеогенез
наиболее интенсивно протекает в условиях повышенного содержания в клетке
топливных молекул, в частности ацетил-СоА, и достаточного количества
АТР.
Глицерол включается в путь глюконеогенеза через дигидроксиацетонфос-
фат, в который он превращается после фосфорилирования (с участием АТР) и
дегидрирования.
Аминокислоты поступают в путь через такие метаболиты, как пируват и
оксалоацетат, образующиеся в ходе перестроек их углеродных скелетов. Лак-
тат перед вступлением в глюконеогенез должен окислиться до пирувата.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)