Белясова Н.А. Биохимия и молекулярная биология
Подождите немного. Документ загружается.


90
Рис 4.9. Движение молекул воды в соответствии с закономерностями осмоса. Рас-
твор А содержит более высокую концентрацию глюкозы, чем раствор В, и явля-
ется по отношению к нему гипертоническим. Соответственно раствор В по отно-
шению к раствору А является гипотоническим. Результатом осмотического пере-
мещения воды становится уравнивание концентраций и воды, и
глюкозы: растворы становятся изотоническими по отношению друг к другу
Скорость диффузии веществ через мембраны зависит от многих причин:
растворимости вещества в мембране, коэффициента диффузии в мембране, а
также разности концентрации вещества снаружи и внутри клетки (градиента
концентрации) (рис. 4.10).
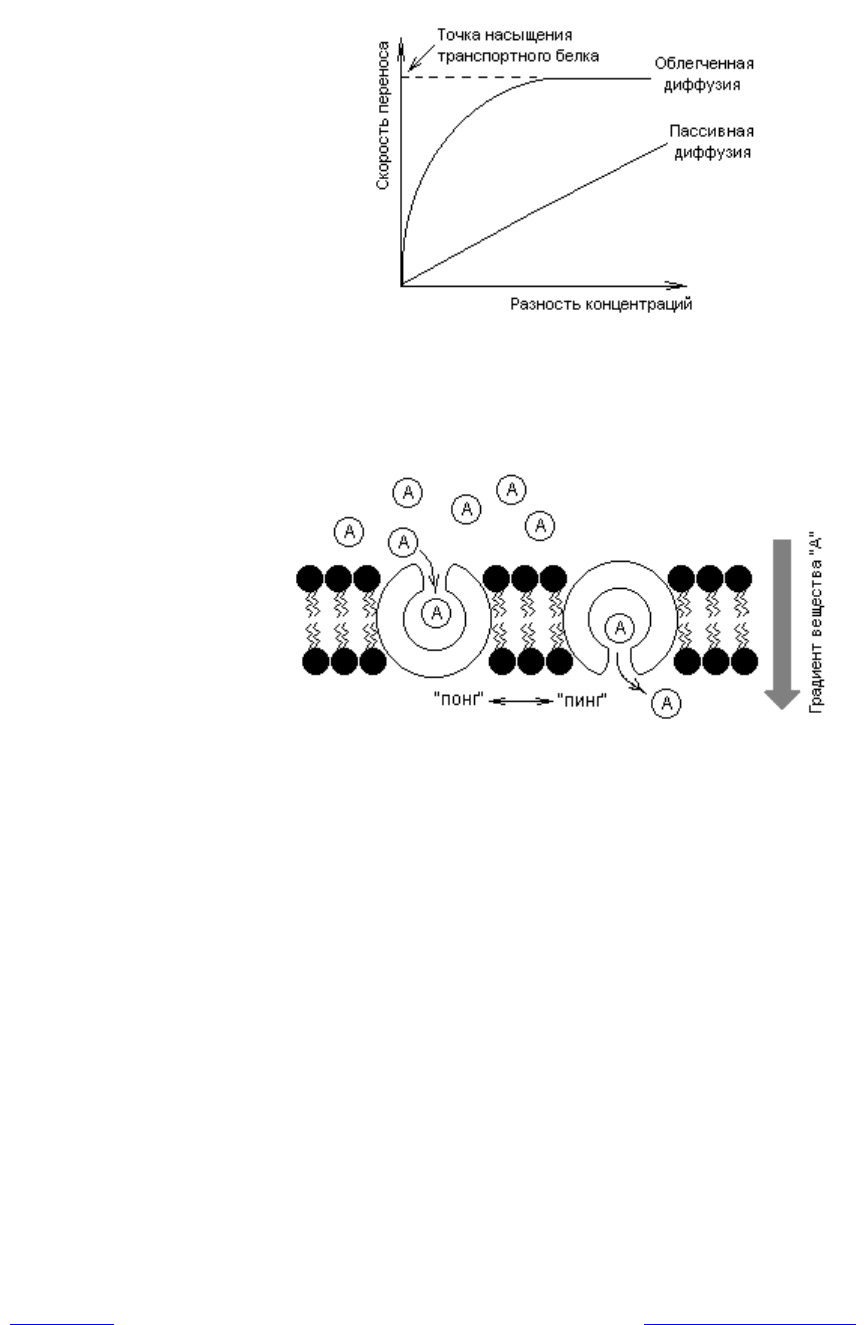
Облегченная диффузия. Этот вид транспорта осуществляется с помощью
транспортных белков по электрохимическому градиенту (разность элек-
трических потенциалов и концентраций веществ) без затрат энергии. Это се-
лективный перенос веществ — вещество будет транспортировано через мем-
брану лишь в том случае, если для него в мембране имеется функционирую-
щий транспортный белок. Поскольку в облегченной диффузии задействованы
белки, этот процесс, в отличие от пассивной диффузии, может достигать эф-
фекта насыщения. Стадия насыщения (рис. 4.10) характеризует состояние,
когда все транспортные белки для данного вещества насыщены субстратом и
скорость транспорта этого вещества достигает максимума.
С помощью облегченной диффузии через мембрану транспортируются
многие вещества, в том числе гидрофильные молекулы: углеводы, аминокис-
лоты, нуклеотиды, различные ионы и др. При этом скорость транспорта зна-
чительно превышает скорость пассивной диффузии (рис. 4.10). Принципи-
ально возможны два пути переноса веществ и ионов через мембрану: с по-
мощью переносчиков и каналов. Поскольку трансмембранное перемещение
белков в биомембранах не обнаружено, предложена модель, описывающая
работу переносчиков — механизм «пинг-понг». Согласно этому механизму,
транспорт веществ связан с конформационными изменениями в структуре
белка-переносчика, которые индуцируются связыванием транспортируемого
вещества (рис. 4.11).
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
91
Рис. 4.10. Зависимость скорости процессов пассивной и облегченной диф-
фузии от разности концентраций транспортируемого вещества
Рис. 4.11. Модель работы интегральных транспортных белков по типу
«пинг-понг»
Работу каналов можно рассмотреть на примере ацетилхолинового рецеп-
тора. Этот интегральный белок находится в основном в мембранах нервно-
мышечных соединений скелетных мышц. Он состоит из пяти субъединиц че-
тырех типов и открывается в ответ на связывание ацетилхолина (нейроме-
диатор). При взаимодействии с ацетилхолином канал открывается, что свя-
зано с изменением конформации субъединиц, и пропускает определенные
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

92
ионы (Na
+
, K
+
, Ca
++
и некоторые другие), остается в таком положении 1 мс, а
затем закрывается. Селективное перемещение катионов изменяет трансмем-
бранный потенциал, в результате чего происходит электрическое возбужде-
ние мышечной клетки, что приводит к сокращению мышцы. Изучение струк-
туры ацетилхолинового рецептора показало, что пять белковых субъединиц
встроены в бислой определенным образом: они организованы вокруг цен-
тральной поры диаметром 3 нм, через которую и транспортируются катионы.
Непроницаемость канала для анионов и в три раза большую проницаемость
для катионов, чем для незаряженных молекул, можно объяснить электроста-
тическими взаимодействиями, возникающими благодаря присутствию в во-
ротах канала биполярных или отрицательно заряженных групп.
Особым типом транспорта веществ в ходе облегченной диффузии являет-
ся использование ионофоров, действие которых изучено на искусственных
мембранах. Под ионофорами понимают низкомолекулярные вещества пеп-
тидной природы, избирательно транспортирующие через мембраны ионы.
Различают ионофоры-каналообразователи (грамици-дин А, амфотерицин В
и др.) и ионофоры-переносчики (валиномицин, энниатины, боверицин).
Валиномицин представляет собой антибиотик депсипептидной природы,
организованный по типу ионной «ловушки». В неполярных растворителях
конформация валиномицина напоминает собой браслет, внутренняя полость
которого точно подогнана под размеры ионов калия. Внешняя сфера валино-
мицина гидрофобна, в результате чего он способен перемещаться в липидном
бислое и транспортировать через него ионы.
Хорошо изученным примером ионофоров-каналообразователей служит
грамицидин А. Это антибиотик пептидной природы, состоящий из
15 аминокислот. Две молекулы грамицидина могут пронизывать мембрану в
виде двойной спирали или образуя димер «голова к голове». В таких кон-
формациях молекулы грамицидина А формируют полый цилиндр, по которо-
му могут перемещаться ионы металлов.
В биологических мембранах ионофорный тип транспорта до сих пор не
обнаружен.
Активный транспорт. Это сопряженный с потреблением энергии пере-
нос молекул или ионов через мембрану против электрохимического градиен-
та, в котором задействованы транспортные белки. Благодаря активному
транспорту в жизнеспособных клетках между двумя сторонами мембраны
поддерживается разность потенциалов, т. е. электрический заряд, при этом у
большинства изученных клеток внутреннее содержимое заряжено отрица-
тельно по отношению к внешней среде.
Активный транспорт сопряжен со значительными затратами энергии: не-
которые клетки тратят более трети всей запасенной энергии для создания
ионного градиента на мембране. Это необходимо для таких жизненно важ-
ных процессов, как осморегуляция, генерация и передача нервных импульсов,
перенос в клетки питательных веществ (сахаров, аминокислот и др.).
Разнообразные системы активного транспорта отличаются друг от друга,
в первую очередь тем, что служит для них источником энергии: АТР, ионный
градиент, фосфоенолпируват, видимый свет. Наиболее хорошо изученной
системой активного транспорта является натрий-калиевая (Na
+
/K
+
)-АТР-аза,
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

93
функционирующая в плазматических мембранах животных клеток. Этот ин-
тегральный белок состоит из двух субъединиц: бульшая представлена поли-
пептидом, имеющим участки связывания для ионов натрия и АТР на цито-
плазматической поверхности, а ионов калия — на наружной; меньшая субъе-
диница является гликопротеином. Работа (Na
+
/K
+
)-АТР-азы заключается в
следующем: при гидролизе одной молекулы АТР из клетки выкачивается
3 иона Na
+
, а извне в клетку проводится 2 иона K
+
, т. е. выводится больше
положительных ионов, чем проводится внутрь клетки. Так на внутренней
стороне мембраны создается избыточный отрицательный заряд, и клетка ста-
новится электрогенной. В мембранах обычно присутствуют проводящие пути
для облегченной диффузии ионов натрия и калия по электрохимическому
градиенту, и этот транспорт, хотя и с малой скоростью, совершается. Однако
в жизнеспособной клетке не происходит уравнивания концентраций ионов,
создающих электрохимический градиент на мембране, благодаря постоянной
работе первичных активных переносчиков, таких, как (Na
+
/K
+
)-АТР-аза. Та-
ким образом, ионные насосы, принимающие участие в первичном активном
транспорте, осуществляют перемещение заряда на мембране и создают на
ней электрохимический градиент, в котором заключена энергия.
Вторичные активные переносчики используют электрохимические гради-
енты в качестве движущей силы для транспорта растворимых веществ. Этот
процесс можно проследить на примере клеток эпителия кишечника. Образуе-
мые в кишечнике при переваривании пищи строительные блоки (аминокис-
лоты, глюкоза и др.) поступают в кровь при диффузии через мембраны кро-
веносных сосудов, и эта диффузия осуществляется в ходе симпорта (однона-
правленного транспорта) с ионами натрия. Ионы натрия стремятся возвра-
титься в клетку согласно закономерностям облегченной диффузии и как бы
тянут с собой молекулы питательных веществ. В мембранах обнаружены спе-
цифические переносчики сахаров и разных аминокислот, которые функцио-
нируют в системе активного транспорта, накапливая в клетке эти вещества,
извлекая их даже из очень разбавленных растворов, т. е. против химического
градиента. Эти же транспортные системы могут участвовать и в облегченной
диффузии, если вещества транспортируются по химическому градиенту.
Кроме описанного выше примера симпорта питательных веществ вместе с
возвращающимися в клетку ионами натрия, существует и разнонаправленный
транспорт — антипорт. Например, белок полосы 3 эритроцитов осуществля-
ет сопряженный транспорт Cl
-
и HCO
3
-
в противоположных направлениях
через эритроцитарную мембрану.
У аэробных бактерий транспорт питательных веществ в клетку осуществ-
ляется в ходе симпорта не с ионами Na
+
, а с протонами. Наилучшим образом
охарактеризованным примером подобного переносчика служит лактозопер-
меаза кишечной палочки. Этот интегральный белок использует протонный
электрохимический градиент, созданный на мембране в результате окисли-
тельного фосфорилирования, для симпорта лактозы: с каждым возвращен-
ным в клетку протоном переносится одна молекула лактозы.
Следует отметить, что АТР-азы представляют собой ферменты, катализи-
рующие взаимообратимые реакции: при гидролизе АТР ионы транспортиру-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

94
ются против электрохимического градиента, а перенос ионов по электрохи-
мическому градиенту через каналы АТР-азы может запускать синтез АТР.
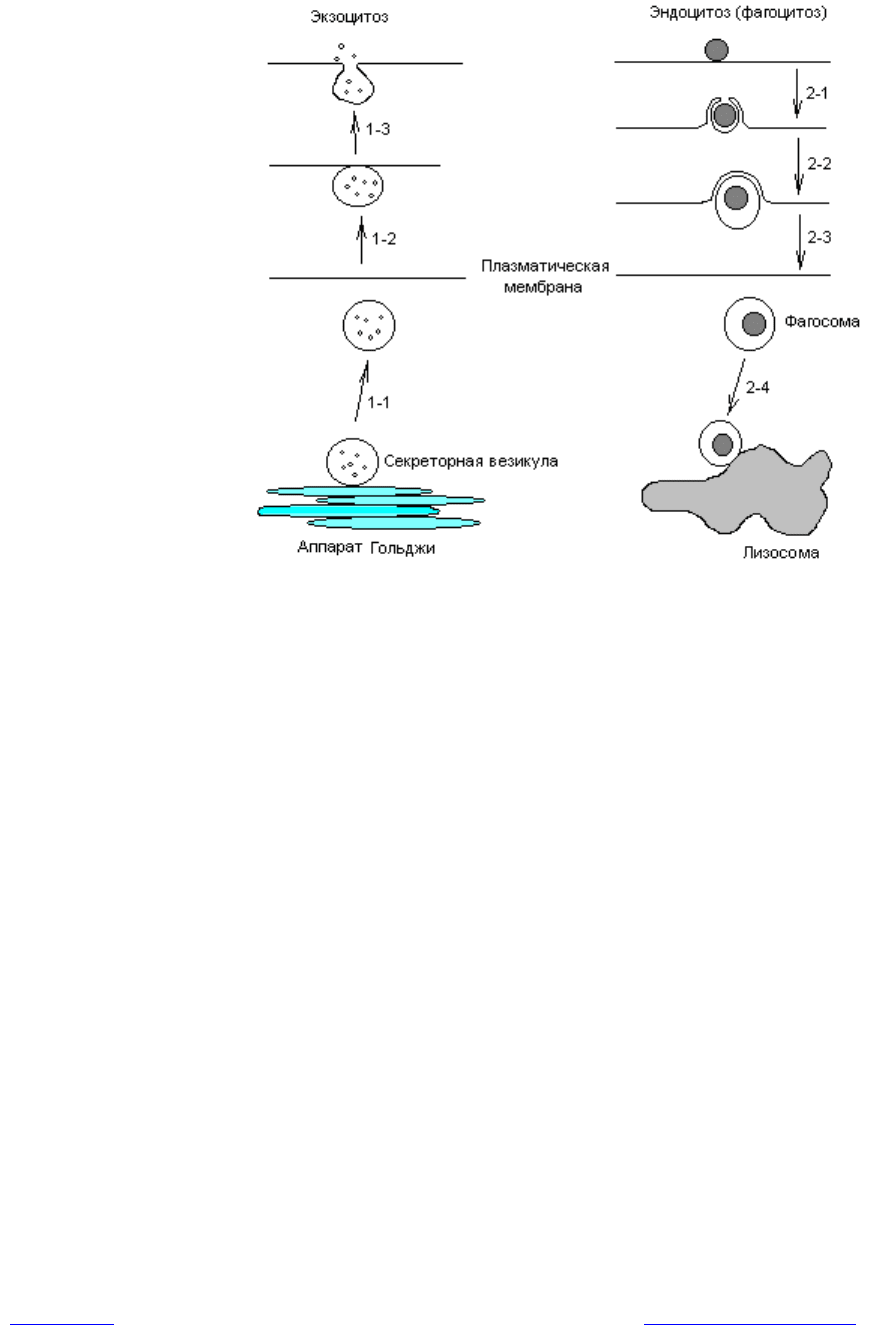
Эндоцитоз и экзоцитоз. Эти способы переноса веществ через мембраны
связаны с образованием впячиваний (инвагинаций) мембраны и формиро-
ванием особых мембранных везикул, обеспечивающих прохождение через
мембрану крупных макромолекул и частиц. При этом эндоцитоз обеспечивает
поглощение клеткой веществ, а экзоцитоз — выделение из клетки. Принято
делить цитозы еще на два типа: пиноцитоз и фагоцитоз. Пиноцитоз — это
механизм, с помощью которого через мембрану проводятся белки и другие
макромолекулы в жидкой фазе. Фагоцитоз представляет собой поглощение
клеткой крупных частиц, например бактерий, вирусов. Эти виды транспорта
характерны в основном для эукариотических клеток, причем у животных фа-
гоцитоз осуществляют только специализированные клетки, такие, например,
как макрофаги. Для многих простейших, например амеб, фагоцитоз является
основным способом питания.
Важной особенностью цитозов является последовательное образование и
слияние везикул, в которых заключено транспортируемое вещество, причем
секретируемые и поглащаемые молекулы локализуются в везикулах и не
смешиваются с другими макромолекулами или органеллами клетки. С помо-
щью не установленного пока механизма каждый пузырек сливается только со
специфическими мембранными структурами (рис. 4.12).
В основе цитозов лежит еще одно характерное свойство липидных слоев
биомембран — способность к агрегации, в результате чего мелкие везикулы
объединяются в более крупные или происходит объединение везикул с плаз-
матической мембраной клетки. Такой механизм основан на универсальности
структуры биомембран, участвующих в формировании клеточных органелл и
протопластов. Аналогичное явление можно наблюдать в пенах, где мыльные
пузыри, также состоящие из амфифильных молекул (мыла — соли жирных
кислот), обладают тенденцией к объединению с образованием более крупных
структур. Способность мембран к агрегации лежит в основе такого широко
распространенного способа переноса генетической информации, как слияние
протопластов.
Скорость цитозов удивительно высокая. Показано, что клетки печени по-
глощают путем эндоцитоза за 1 ч количество жидкости, составляющее не
менее 20% их объема, и количество мембранного материала, по площади
превышающее в пять раз площадь их плазматической мембраны. Сходство
цитозов с другими способами транспорта веществ через биомембраны состо-
ит в том, что переносимое вещество должно быть «узнано» мембранными
компонентами, иными словами, и в этом случае проявляется избирательная
проницаемость мембран для различных соединений.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
95
Рис. 4.12. Схематическое изображение цитозов. Стадии: 1–1 — от аппарата
Гольджи отшнуровывается секреторная везикула; 1–2 — везикула диффунди-
рует к плазматической мембране и слипается с ней; 1–3 — слияние везикулы
с мембраной и выход содержимого везикулы в окружающую среду; 2–1 —
частица обволакивается мембраной; 2–2 — фагосома отшнуровывается от
мембраны; 2–3 — фагосома диффундирует к лизосоме;
2–4 — фагосома слипается с лизосомой и сливается с ней
4.6. Рецепторные функции мембран
Все клетки должны обладать системами, позволяющими определять со-
стояние и изменения окружающей среды, чтобы адаптироваться к ним. Эти
системы представляют собой разнообразные рецепторные молекулы, которые
располагаются в поверхностных структурах, чаще всего — в плазматических
мембранах, реже — в клеточных стенках, причем у грамотрицательных бак-
терий — в наружной мембране. Функция рецепторных молекул и их ассоциа-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

96
ций состоит во взаимодействии с внеклеточными компонентами и иницииро-
вании специфического клеточного ответа.
Рецепторные молекулы в большинстве случаев представлены белками, но
эту роль могут выполнять и другие молекулы, например гликолипиды, глико-
протеины или сфинголипиды. Так, показано, что ганглиозиды служат местом
связывания холерного и столбнячного токсинов, а также участвуют в регуля-
ции процессов клеточного роста и дифференцировки.
Среди огромного разнообразия клеточных рецепторов можно выделить
несколько основных типов. В поверхностных структурах бактериальных,
дрожжевых, животных клеток присутствуют рецепторы, определяющие спо-
собность клеток распознавать друг друга, взаимодействовать, образуя скоп-
ления, а также связываться с нерастворимыми компонентами внеклеточного
матрикса. Примером рецепторов указанного типа служат белковые ворсинки,
обнаруженные у патогенных штаммов E. coli, которые вызывают инфекцион-
ные заболевания мочевых путей человека. Ворсинки крепятся в наружной
мембране и содержат на конце рецепторный белок — адгезин, способный
специфически связываться с дигалактозидсодержащими гликолипидами. Эти
липиды присутствуют на поверхности эпителиальных клеток, выстилающих
мочевые пути, где размножаются бактерии.
Другой класс рецепторов представлен молекулами, расположенными в
плазматических мембранах организмов и связывающими питательные веще-
ства и метаболиты. Эти рецепторы участвуют в процессах эндо- и экзоцитоза,
определяя специфичность этих видов транспорта.
Более сложные рецепторные реакции сопровождаются связыванием ре-
цептора с метаболитом, гормоном или нейромедиатором, передачей сигнала
внутрь клетки и следующим затем клеточным ответом. К подобному классу
рецепторов относятся, например, белки бактерий, ответственные за хемотак-
сис. В составе плазматической мембраны E. coli присутствует рецептор для
аспартата, который представляет собой трансмембранный белок. Этот белок
осуществляет связывание аспартата, что влечет за собой конформационное
изменение в той части молекулы, которая обращена в цитоплазму. Это изме-
нение и служит сигналом, заставляющим опосредованным образом (через
фосфорилирование другого белкового компонента системы) вращаться жгу-
тики. В результате клетка перемещается по градиенту концентрации аспарта-
та, получая возможность использовать его в качестве питательного субстрата.
Клеточный ответ на сигнал, обусловленный рецепцией специфического веще-
ства, может выражаться также в активации транскрипции отдельных генов. В
такую рецепторную систему входит белок-регулятор, находящийся, по-
видимому, в цитоплазме в растворимой форме. Считается, что рецепторы
каким-то образом модифицируют регуляторные белки, и затем последние
активируют транскрипцию.
Аналогичным образом происходит передача сигнала при связывании
лиганда (нейромедиатора или гормона) со специфическим рецептором на
наружной поверхности мембраны животной клетки. Это событие инициирует
конформационный переход в молекуле рецептора и следующий затем каскад
событий в клетке, который может включать открывание канала (никотиновый
ацетилхолиновый рецептор), фосфорилирование клеточных белков, сопрово-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

97
ждающееся изменением их активности, образование комплекса с G-белками.
В последнем случае G-белки активируются, высвобождаются из комплекса и
диффундируют к клеточным мишеням, вызывая специфический ответ. Одной
из наиболее распространенных мишеней G-белков является аденилатциклаза
(катализирует образование сАМР). Конформационное изменение этого фер-
мента приводит к изменению внутриклеточной конценрации сАМР, который,
как известно (глава 3), служит вторым посредником, влияя на множество
внутриклеточных процессов.
Наконец, многие клетки имеют в составе мембран рецепторы, способные
в ответ на стимул (внешний сигнал) генерировать нервный импульс. Нервный
импульс, возникший в мембране специализированной рецепторной клетки
передается через синапсы по отросткам центростремительных нервных кле-
ток к центральной нервной системе, а затем по отросткам центробежных
нервных клеток — к мышце или железе. В клетках скелетных мышц при этом
возбуждается ацетилхолиновый рецептор и возникает потенциал действия, а
через короткий промежуток времени (около 35 мс) происходит сокращение за
счет движения актина и миозина внутриклеточных миофибрилл.
Специализированные рецепторные клетки у высших животных и человека
могут формировать органы чувств. Работа этих органов основана на измене-
нии электрических характеристик рецепторных клеток в ответ на специфиче-
ский стимул, т. е. на свойстве клеток генерировать нервный импульс. Более
подробно эти процессы рассмотрены в главе 13 на примере функционирова-
ния органа зрения.
4.7. Генерация и проведение нервных импульсов
Благодаря непрерывной работе мембранных ионных насосов между внут-
ренним содержимым невозбужденной клетки и внешней средой постоянно
поддерживается разность потенциалов. Этот параметр можно измерить вве-
дением электрода в крупные нервные клетки, диаметр которых у кальмаров,
например, может достигать 1 мм. Определяемая величина называется потен-
циалом покоя и достигает в невозбужденных клетках (-60) — (-70) мВ (ми-
нус указывает на то, что внутренняя поверхность мембраны электроотрица-
тельна по отношению к внешней). Потенциал покоя характеризует состояние
динамического равновесия в перемещении ионов натрия и калия через мем-
брану.
При воздействии на мембрану в каком-либо локусе электрическим током
или другим раздражителем в этом месте открываются специфические натрие-
вые каналы и ионы Na
+
устремляются внутрь, нейтрализуя отрицательный
заряд на внутренней поверхности мембраны в данном локусе. Это явление
носит название «деполяризация». Если пороговая величина импульса, вы-
звавшего деполяризацию, достаточно велика (более 10 мВ), то в мембране
открываются натриевые каналы, работающие на потенциал действия. По-
тенциал действия представляет собой импульс, проходящий вдоль мембраны
нервной клетки и специфически изменяющий за доли секунды (в нервах мле-
копитающих приблизительно за 0,5 мс) мембранный потенциал. Особенно-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

98
стями потенциала действия являются: высокая скорость распространения
вдоль мембраны нервной клетки — до 100 м/с и неизменная интенсивность.
Механизм передачи нервного импульса состоит в следующем. Повышение
проводимости мембраны в конкретном локусе для ионов натрия и деполяри-
зация взаимно усиливают друг друга. В результате быстрого перемещения
ионов натрия на внутреннюю поверхность мембраны исходный потенциал (-
70 мВ) падает до нуля, а затем достигает значения (+20) — (+40) мВ. Таким
образом, в данном локусе мембраны создается область положительного заря-
да и в местной цепи возникает ток между этим активным участком и отрица-
тельно заряженной областью, находящейся непосредственно перед ним. От-
личительной особенностью мембран нервных клеток, очень быстро проводя-
щих нервные импульсы, является наличие миелиновой оболочки, которая
выполняет роль изолятора. Благодаря этой оболочке плазматическая мембра-
на контактирует с содержащей Na
+
внеклеточной жидкостью лишь в опреде-
ленных точках — перехватах Ранвье, и деполяризация мембраны происходит
толь-ко в области этих перехватов. Так как нервная клетка покрыта изолято-
ром — миелином, распространение области электрического тока до следую-
щего перехвата осуществляется в основном через цитоплазму клетки. Дойдя
до очередного перехвата, ток снижает мембранный потенциал в этой области,
в ней происходит деполяризация и повышается проницаемость для ионов на-
трия, т. е. возникает потенциал действия. Так, в виде волны деполяризации
потенциал действия распространяется по мембране в одном направлении.
Восстановление поляризации происходит в результате выхода из аксона
ионов К
+
. Пока реполяризация не закончится, перехват не может возбудиться
снова, а к тому времени, когда перехват будет способен к ответу, импульс уже
уйдет по мембране слишком далеко, чтобы снова открыть в рассматриваемом
перехвате аксона натриевые каналы. По этой причине импульс распространя-
ется по мембране нервной клетки только в одном направлении. Реполяриза-
ция заканчивается после того, как АТР-азы, задействованные в активном
транспорте ионов, восстановят нормальное соотношение концентраций Na
+
и
К
+
в перехвате.
Амплитуда потенциала действия постоянна для каждой нервной клетки и
не уменьшается по мере движения по мембране, так как изменение в каждой
точке мембраны происходит за счет локального запаса энергии в виде ионно-
го градиента, хотя и запускается изменением в соседнем перехвате.
Передача нервного импульса между нейронами и от нейронов к мышеч-
ным клеткам происходит в нервных окончаниях (синапсах) с помощью сиг-
нальных веществ — медиаторов. Одним из наиболее распространенных
нейромедиаторов (синтезируются нервными клетками) является ацетилхо-
лин. Выделение ацетилхолина синаптическими мембранами служит сигналом
для генерирования нервного импульса в расположенной по соседству нервной
клетке. Осуществляется это с помощью ацетилхолиновых рецепторов, кото-
рые представляют собой трансмембранные комплексы из пяти субъединиц,
формирующие проницаемый для ионов Na
+
и K
+
каналы.
Участки для специфического присоединения ацетилхолина расположены
на внеклеточной части a-субъединицы рецептора и их связывание приводит к
формированию в центре комплекса трансмембранного канала, который от-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

99
крывается на короткое время для прохождения ионов натрия и калия. Откры-
вание и закрывание канала происходит в результате аллостерического изме-
нения в заряженных участках полипептидных цепей рецептора. Перемещение
ионов через мембраны изменяет потенциал покоя нервной или мышечной
клетки, что стимулирует открывание потенциал-управляемых Na
+
-каналов, и
возникает потенциал действия.
Ацетилхолиновый рецептор может связывать различные лекарственные
вещества, например никотин, который в малых дозах активирует ацетилхо-
линовые рецепторы, а в больших — угнетает их. Кроме этого, многие нейро-
токсины змей способны взаимодействовать с ацетилхолиновым рецептором и
нарушать его функционирование. Известно также явление конкурентного ин-
гибирования ацетилхолина при связывании с рецептором ядом кураре (расти-
тельный алкалоид), которым южно-американские индейцы пропитывали на-
конечники для стрел.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)