Белясова Н.А. Биохимия и молекулярная биология
Подождите немного. Документ загружается.


279
триптофаном при репрессии), комплексы сАМР-САР, гуанозинтетрафосфат, а
в некоторых случаях таким действием обладают белки — продукты экспрес-
сии собственных генов. Особое значение в данных процессах имеют такие
важные сигнальные молекулы, как сАМР и гуанозинтетрафосфат. Можно
сказать, что сАМР сигнализирует клетке об энергетическом голоде —
отсутствии глюкозы. В ответ на это увеличивается частота транскрипции
структурных генов, отвечающих за катаболизм других источников углерода и
энергии (активация катаболитных оперонов, катаболитная репрессия, гла-
ва 3). Гуанозинтетрафосфат (гуанозин-5ў-дифосфат-3ў-дифосфат) является
сигналом аминокислотного голодания. Этот нуклеотид связывается с РНК-
полимеразой и изменяет ее сродство к промоторам различных генов. В ре-
зультате экспрессия генов, ответственных за биосинтез углеводов, липидов,
нуклеотидов и др. уменьшается, а экспрессия других генов, в частности де-
терминирующих процессы протеолиза белков, наоборот, повышается.
Процесс транскрипции чаще регулируется с помощью изменения частоты
событий инициации транскрипции, но, кроме этого, могут регулироваться
скорость элонгации транскрипции и частота ее преждевременной термина-
ции. На события элонгации и терминации первостепенное влияние оказывает
конформационное состояние ДНК или самой мРНК (наличие «стоп-
сигналов», шпилечных структур).
Аллостерическая регуляция активности ферментов. Этот тип регу-
ляции является одним из самых быстрых и гибких, он осуществляется с по-
мощью молекул-эффекторов, взаимодействующих с аллостерическим цен-
тром фермента (глава 6). Аллостерической регуляции, как и оперонной, под-
вержены ключевые ферменты тех или иных метаболических путей. Таким
образом, скорость всего биосинтетического или катаболического процесса
зависит от одной, реже нескольких реакций, катализируемых ключевыми
ферментами.
Особое значение регуляция имеет для процессов биосинтеза протеиноген-
ных аминокислот. Поскольку их 20, и каждая в суммарном клеточном белке у
разных организмов представлена в определенном отношении, требуется
очень четкая регуляция, координирующая процессы синтеза отдельных ами-
нокислот. Такой контроль исключает перепроизводство аминокислот, и выде-
ление их из клетки возможно лишь у микроорганизмов с нарушенной регуля-
цией.
Пример регуляции биосинтеза аминокислот семейства аспартата у энте-
робактерий представлен на рис. 19.3. Четыре аминокислоты имеют общий
предшественник — аспарагиновую кислоту. Ее превращение в аспартилфос-
фат у бактерий E.coli катализируют три изоферментные формы аспартокина-
зы, каждая из которых испытывает репрессию и/или ингибирование со сторо-
ны разных конечных продуктов данного разветвленного метаболического
пути. Аналогичным способом регулируется синтез гомосериндегидрогеназы.
Обращает на себя внимание существование механизма обратной связи,
который заключается в том, что конечные продукты метаболических процес-
сов регулируют уровень синтеза и/или активность ферментов, катализирую-
щих первые этапы образования этих метаболитов.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
280
Аллостерическими эффекторами могут выступать самые различные веще-
ства: субстраты и конечные продукты метаболических путей, иногда
А
с
п
а
р
т
а
т
Гомосерин
Лизин
Метионин
Треонин
Изолейцин
Аспартокиназа
Гомосерин
дегидрогеназа
Гомосеринкиназа
Треонин
синтаза
Дигидродипиколинат
синтетаза
Треонин
дезаминаза
репрессия
ингибирование
Аспартил-4-фосфат
4-Полуальдегид-аспартат
1, 2, 3 - изоферментные формы
аспартокиназы и
гомосериндегидрогеназы
1
1
2
2
3
-
-
-
-
-
-
Рис. 19.3. Схема регуляции синтеза и активности ферментов, участвующих в
пути биосинтеза аминокислот семейства аспартата у бактерий E.coli. Гены,
кодирующие аспартокиназу 1, гомосеринкиназу и треонинсинтетазу, со-
ставляют треониновый оперон, поливалентно репресси-
руемый треонином и изолейцином
— промежуточные метаболиты; в катаболических процессах — нуклео-
зиддифосфаты и нуклеозидтрифосфаты, а также переносчики восстанови-
тельных эквивалентов; в каскадных реакциях — сАМР и сGMP, которые ре-
гулируют активность ферментов (например, протеинкиназ), участвующих в
ковалентной модификации белков; ионы металлов и множество иных соеди-
нений. Примеры аллостерической регуляции ферментов приведены в главе 6
и др. разделах.
Ковалентная модификация ферментов. Этот тип регуляции активности
ферментов иначе называют взаимопревращениями ферментов, поскольку
суть данного процесса состоит в превращении активных форм ферментов в
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

281
неактивные и наоборот. Особенности и примеры ковалентной модификации
описаны в главе 6. Эти процессы находятся под разнообразным контролем, в
том числе и гормональным. Классическим примером взаимопревращений
ферментов является регуляция метаболизма гликогена в печени.
Скорость синтеза этого резервного полисахарида находится под контро-
лем гликоген-синтазы, а расщепление катализируется гликоген-
фосфорилазой. Оба фермента могут пребывать в активной и неактивной
формах. При голодании или в стрессовых ситуациях в кровь выделяются
гормоны — адреналин и глюкагон, которые связываются с рецепторами на
плазматических мембранах клеток и активируют при посредничестве G-
белков фермент аденилатциклазу (катализирует синтез сАМР). сАМР связы-
вается с протеинкиназой А и активирует ее, что приводит к фосфорилирова-
нию гликоген-синтазы и переводу ее в неактивную форму. Гликоген переста-
ет синтезироваться. Кроме этого, протеинкиназа А в ходе каскадных реакций
вызывает фосфорилирование гликоген-фосфорилазы, которая в результате
активируется и начинает расщеплять гликоген. На процессы синтеза и распа-
да гликогена действует также другой гормон —инсулин. В этом примере сиг-
нальными молекулами служат гормоны, а посредниками — G-белок и сАМР.
Взаимопревращения ферментов осуществляются в ходе фосфорилирования—
дефосфорилирования.
Гормональная регуляция. Этот тип регуляции метаболизма предусмат-
ривает участие гормонов — сигнальных веществ, образующихся в клетках
эндокринных желез, поэтому гормональная регуляция свойственна только
высшим организмам. Выше описано действие гормонов на процесс обмена
гликогена, в котором регулируется активность ферментов на уровне кова-
лентной модификации. Кроме этого, гормоны способны оказывать воздействие
на скорость транскрипции (оперонная регуляция).
Из специализированных клеток, где происходит синтез гормонов, послед-
ние поступают в кровь и переносятся к клеткам-мишеням, имеющим рецеп-
торы, способным связывать гормоны и тем самым воспринимать гормональ-
ный сигнал. Связывание гормона рецептором запускает каскад реакций с уча-
стием молекул-посредников, которые завершаются клеточным ответом. Ли-
пофильные гормоны связываются с внутриклеточным рецептором (белок) и
регулируют транскрипцию определенных генов. Гидрофильные гормоны дей-
ствуют на клетки-мишени за счет связывания с рецепторами на плазматиче-
ской мембране.
Кроме гормонов, аналогичным действием обладают другие сигнальные
вещества: медиаторы, нейромедиаторы, ростовые факторы. Четкой границы,
позволяющей отличать гормоны от перечисленных веществ, нет. Медиатора-
ми называют сигнальные вещества, которые продуцируются не железами
внутренней секреции, а различными типами клеток. К медиаторам относят
гистамин, простагландины, которые обладают гормоноподобным действием.
Нейромедиаторами считают сигнальные вещества, продуцируемые клет-
ками центральной нервной системы.
Изменение концентрации метаболитов. Важным условием, обеспечи-
вающим высокую скорость того или иного метаболического пути, является
концентрация субстратов. Она может зависеть от интенсивности протекания
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

282
других процессов, в которых также расходуются эти субстраты (конкурен-
ция), или от скорости транспорта данных веществ через мембраны (плазма-
тическую или органелл). В частности, у эукариотических клеток появляется
возможность регулировать метаболизм, перераспределяя метаболиты по от-
дельным компартментам.
Кроме этого, скорость метаболических процессов определяется концен-
трацией кофакторов. Например, гликолиз и ЦТК регулируются доступностью
ADP (глава 10, 11) на уровне изменения активности ключевых аллостериче-
ских ферментов.
Посттранскрипционная и посттрансляционная модификация мак-
ромолекул. Эти процессы также описаны в соответствующих разделах (гла-
ва 3). Модификация и/или процессинг первичных РНК-транскриптов осуще-
ствляются с разной скоростью, от чего зависит концентрация зрелых молекул
РНК, способных транслироваться, а значит, и интенсивность белкового син-
теза. В свою очередь, пептиды, прежде чем превратиться в зрелый белок,
также должны модифицироваться, и если это касается ферментов, то речь
идет об их ковалентной модификации.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

283
Часть пятая. ОСНОВЫ ГЕНЕТИЧЕСКОЙ
ИНЖЕНЕРИИ
Глава 20. СОЗДАНИЕ И АНАЛИЗ КЛОНОТЕК ГЕНОМОВ
Генетическая инженерия как основа современной биотехнологии зароди-
лась в семидесятые годы ХХ ст. в недрах молекулярной биологии. К этому
времени были расшифрованы механизмы основных матричных процессов,
происходящие в клетках прокариот — репликация, транскрипция и трансля-
ция, кроме того, эти процессы были воспроизведены in vitro. Была определе-
на структура генетического кода и синтезирован первый ген (аланиновой
тРНК дрожжей). Удалось выделить и изучить свойства некоторых ферментов,
использующих в качестве субстрата ДНК (рестриктазы, лигазы, ДНК-
полимеразы). Эмбриологи освоили методологию пересадки ядер соматиче-
ских клеток животных (лягушки) взамен удаленного гаплоидного ядра, что
позволило воспроизводить животных неполовым путем, т. е. клонировать
(получать генетически идентичные особи).
Так, впервые появилась надежда на возможность замены определенных
генов (например, дефектных) в зародышевых клетках на другие —
полноценные, иными словами, осуществления генной терапии. Однако для
этого необходимо было освоить методологию выделения генов, их идентифи-
кации, тиражирования, устойчивого культивирования в составе автономно
реплицирующихся молекул, а также определения свойств продуктов этих ге-
нов. Все эти вопросы решает генетическая инженерия, под которой подразу-
мевают комплекс молекулярно-генетических методов, позволяющих осуще-
ствить целенаправленное конструирование организмов путем манипуляций с
их наследственным аппаратом.
Поскольку микроорганизмы, в частности бактерии, как одни из наиболее
просто организованных форм живого, изучены гораздо лучше с генетической
точки зрения, чем макроорганизмы, первые эксперименты по получению ре-
комбинантных ДНК были осуществлены именно с бактериальными клетками.
И вскоре обнаружилась еще одна важная перспектива использования дости-
жений генетической инженерии, а именно конструирование высокопродук-
тивных штаммов микроорганизмов—продуцентов биологически активных
веществ, обладающих комплексом заданных свойств.
В данной главе охарактеризованы основные методы, позволяющие полу-
чать определенные гены, вводить их в состав векторных молекул для клони-
рования в клетках-реципиентах, идентифицировать гены в клонотеках, опре-
делять последовательность нуклеотидов в ДНК. Знание данной методологии
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

284
необходимо для современных биотехнологов, использующих для создания
организмов — продуцентов нужных веществ достижения генетической инже-
нерии.
Для конструирования организмов с заданными свойствами требуется
иметь набор генов, детерминирующих желаемые функции (утилизация опре-
деленных субстратов, биосинтез и секреция определенных продуктов, устой-
чивость к определенным физическим и химическим факторам, деградация
ксенобиотиков и т. п.). Такие гены можно выделить из любых организмов,
обладающих соответствующими свойствами, но для этого вначале следует
получить клонотеку генома данного организма и охарактеризовать ее. Под
клонотекой генома понимают набор клонов бактерий или бактериофагов, ка-
ждый из которых содержит один тип рекомбинантной ДНК, а в совокупно-
сти — весь геном изучаемого организма, фрагменты которого распределены
по отдельным клонам.
20.1. Получение генов
Используется три основных способа получения нужных генов: выделение
из состава ДНК, химико-ферментативный синтез (in vitro), транскрипция
изолированной из клетки матричной РНК с помощью обратной транскрипта-
зы (РНК-зависимой ДНК-полимеразы, ревертазы).
Выделение генов из ДНК. Большинство методик в генетической инжене-
рии основано на вырезании из молекул ДНК определенных фрагментов и
соединении их с другими фрагментами для получения рекомбинантных
(химерных) ДНК. При этом фрагментация выделенной из клеток ДНК осу-
ществляется с помощью ферментов, которые способны расщеплять ДНК в
строго определенных сайтах. В качестве таких ферментов используются эн-
донуклеазы рестрикции (рестриктазы), общая характеристика которых дана в
главе 2. В генетической инженерии используются рестриктазы, образующие в
ДНК однонитевые (липкие) концы (при разрезе уступом), а также те, что
формируют в ДНК двухнитевые (тупые) концы (при расщеплении по сере-
дине узнаваемого участка нуклеотидных пар). Примером рестриктазы перво-
го типа, образующей липкие концы, является Eco R1 (рис. 2.1), а рестрикта-
зой второго типа является, например, Hind II.
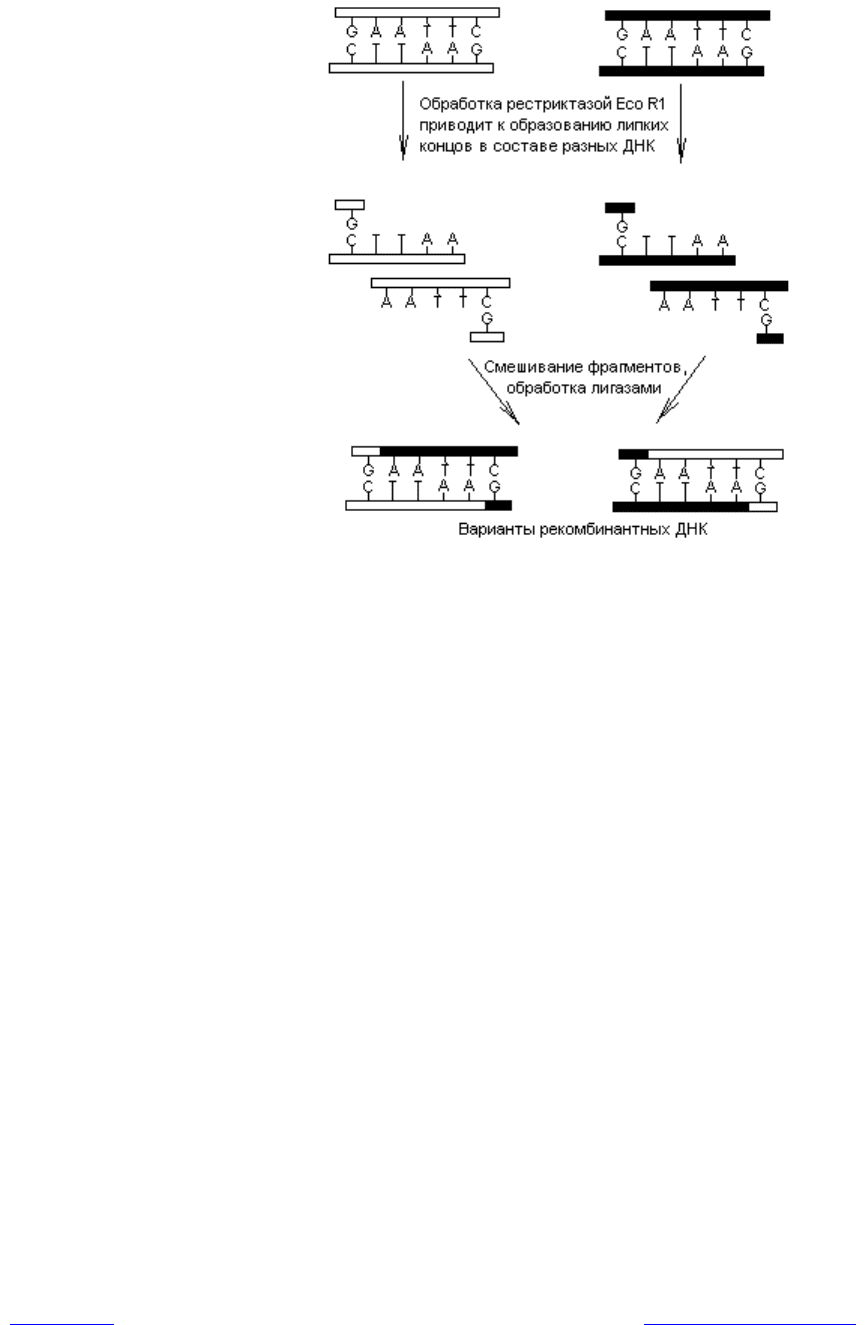
Образование липких концов при расщеплении ДНК имеет преимущество,
которое состоит в возможности реассоциации образованных фрагментов по
гомополимерным липким концам (содержат комплементарные нуклеотиды).
В таком случае появляется возможность формирования ассоциатов из фраг-
ментов, принадлежащих разным молекулам ДНК, что и лежит в основе
большинства генноинженерных манипуляций по получению рекомбинантных
ДНК (рис. 20.1). Образующиеся спонтанно ассоциаты могут быть превраще-
ны в целые молекулы путем «сшивания» с помощью ДНК-лигаз.
Следует указать, что в обычных условиях липкие концы в составе одной
молекулы могут удерживаться друг относительно друга с помощью водород-
ных связей между комплементарными основаниями. Однако
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
285
Рис. 20.1. Схема образования рекомбинантных ДНК при расщеплении
разных молекул ДНК рестриктазами, обнажающими во фрагментах
липкие концы
комплементарные цепочки легко разделить при небольшом нагревании рас-
творов ДНК (денатурация ДНК). При охлаждении липкие концы гибриди-
зуются вновь за счет восстановления водородных связей при соблюдении
принципа комплементарности (отжиг). В результате отжига набора фрагмен-
тов, полученных при воздействии на разные молекулы ДНК одной и той же
рестриктазой, могут образоваться как исходные молекулы ДНК, так и их гиб-
риды — рекомбинантные ДНК.
В 1972 г. в Стэнфорде Дж. Мерц и Р. Дэвис осуществили первый подоб-
ный эксперимент. Позднее оказалось, что не все геномы могут содержать
сайты рестрикции для используемых рестриктаз. Особенно это касается не-
больших молекул: плазмид и профагов, которые настолько малы, что содер-
жат лишь по несколько сайтов рестрикции для небольшого числа рестриктаз.
Данное обстоятельство существенно ограничивает возможности метода, по-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
286
этому была разработана методология введения в геном фрагментов ДНК, со-
держащих необходимые сайты рестрикции.
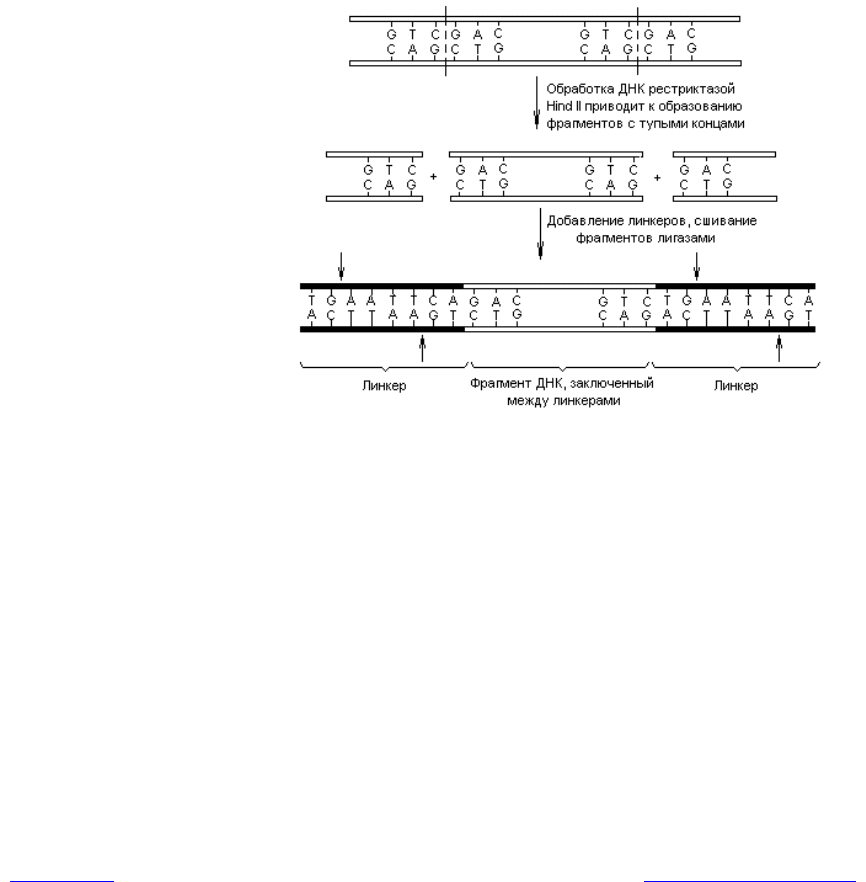
Для этого используют искусственно синтезированные олигонуклеотидные
ДНК с тупыми концами (линкеры). Линкеры синтезируют таким образом,
чтобы в их составе содержались известные сайты рестрикции (предваритель-
но требуется установить последовательность нуклеотидов в этих сайтах).
ДНК для введения линкеров в ее состав расщепляют одной из рестриктаз,
образующих тупые концы, а затем «сшивают» полученные фрагменты с лин-
керами с помощью лигаз (рис. 20.2). Альтернативным методом получения
тупых концов является механический разрыв крупных молекул ДНК при бы-
стром перемешивании раствора или продавливании его через узкое отвер-
стие. В результате этих манипуляций фрагменты ДНК приобретают сайты
рестрикции внутри добавочных последовательностей (линкеров).
Теперь полученный фрагмент ДНК, содержащий требуемые сайты рест-
рикции, можно присоединить к другой молекуле ДНК (например, к вектору),
обработанной такой же рестриктазой, либо превратить в кольцевую форму
путем сшивания взаимно комплементарных концов.
Описанные методы выделения генов в составе фрагментов ДНК с помо-
щью рестрикционных ферментов широко распространены, но имеют ряд не-
достатков. Во-первых, не всегда удается подобрать рестриктазы, позволяю-
щие вырезать из ДНК именно тот участок, в котором содер-
Рис. 20.2. Присоединение линкеров, содержащих сайты рестрикции
(в данном случае для рестриктазы Eco R1 — обозначены
стрелками), к фрагментам ДНК с тупыми концами
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

287
жится интересующий ген. Во-вторых, в составе вырезанного фрагмента ДНК
могут оказаться затрудняющие дальнейшее использование гена последова-
тельности, например интроны в эукариотических генах. В данном случае ре-
комбинантные ДНК не смогут экспрессироваться в прокариотических клет-
ках, поскольку последние не обладают способностью к сплайсингу (глава 3).
Химико-ферментативный синтез генов. С помощью этого метода син-
тезированы, а впоследствии клонированы гены, определяющие структуру та-
ких гормонов, как инсулин и соматостатин, а также лейкоцитарный интерфе-
рон человека. Синтез гена интерферона осуществлен в СССР в 1984 г. под
руководством академика М.Н. Колосова.
Суть метода сводится к следующему: in vitro осуществляют химический
синтез коротких (8—16 нуклеотидов) одноцепочечных фрагментов ДНК, ко-
торые затем соединяют с помощью лигаз и отжигают (дают возможность об-
разоваться двухнитевым молекулам ДНК). Для этого метода необходимо
знать последовательность нуклеотидов в гене, поскольку синтез осуществля-
ется без матрицы. Ее устанавливают обычно исходя из аминокислотной по-
следовательности соответствующего полипептида, однако из-за вырожденно-
сти генетического кода определить истинную нуклеотидную последователь-
ность гена оказывается невозможным. Установить истинную структуру гена
можно методом секвенирования ДНК, однако для этого требуется выделить и
клонировать соответствующий ген.
Этап химического синтеза олигонуклеотидов в настоящее время полно-
стью автоматизирован. Метод основан на специфической защите 5ў или 3ў-
конца моно- или олигонуклеотида, предотвращающей его вступление в хими-
ческие реакции. При необходимости блокирующие группы, с помощью кото-
рых осуществляют модификацию, можно удалить обработкой кислотой либо
щелочью. Цикл химического синтеза ДНК включает конденсацию нуклеоти-
дов, удаление той или иной блокирующей группы и дальнейшую конденса-
цию. Модификацией метода является прикрепление первого нуклеотида к
твердому носителю и добавление следующих нуклеотидов по одному после
промывки носителя на каждом таком этапе.
Воссоединение одноцепочечных фрагментов с помощью лигаз требует
фосфорилирования 5ў-концов, что осуществляется с использованием фермен-
та полинуклеотидкиназы и АТР. Одноцепочечные ДНК можно превратить в
двухцепочечные либо в процессе отжига с комплементарной антипараллель-
ной цепью, также синтезированной химическим путем, либо при достройке
комплементарной цепи ферментом (обычно используют ДНК-полимеразу I).
При сочетании химического синтеза и ферментативных этапов, например, из
66 коротких синтетических фрагментов был воссоздан ген инсулина длиной
514 п. н.
Ферментативный синтез генов. Возможны два принципиально разли-
чающихся способа ферментативного синтеза ДНК. Один из них не требует
присутствия матрицы и осуществляется по программе, задаваемой экспери-
ментатором. Такой синтез катализирует бактериальный фермент полинуклео-
тидфосфорилаза, специфичный в отношении рибонуклеотидов, но способный
с меньшей скоростью и к полимеризации цепей ДНК. Для такого синтеза не-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
288
обходим праймер, включающий не менее 3 нуклеотидов. Реакции полимери-
зации имеют некоторые ограничения и с трудом поддаются контролю.
Другой способ ферментативного синтеза ДНК предполагает участие мат-
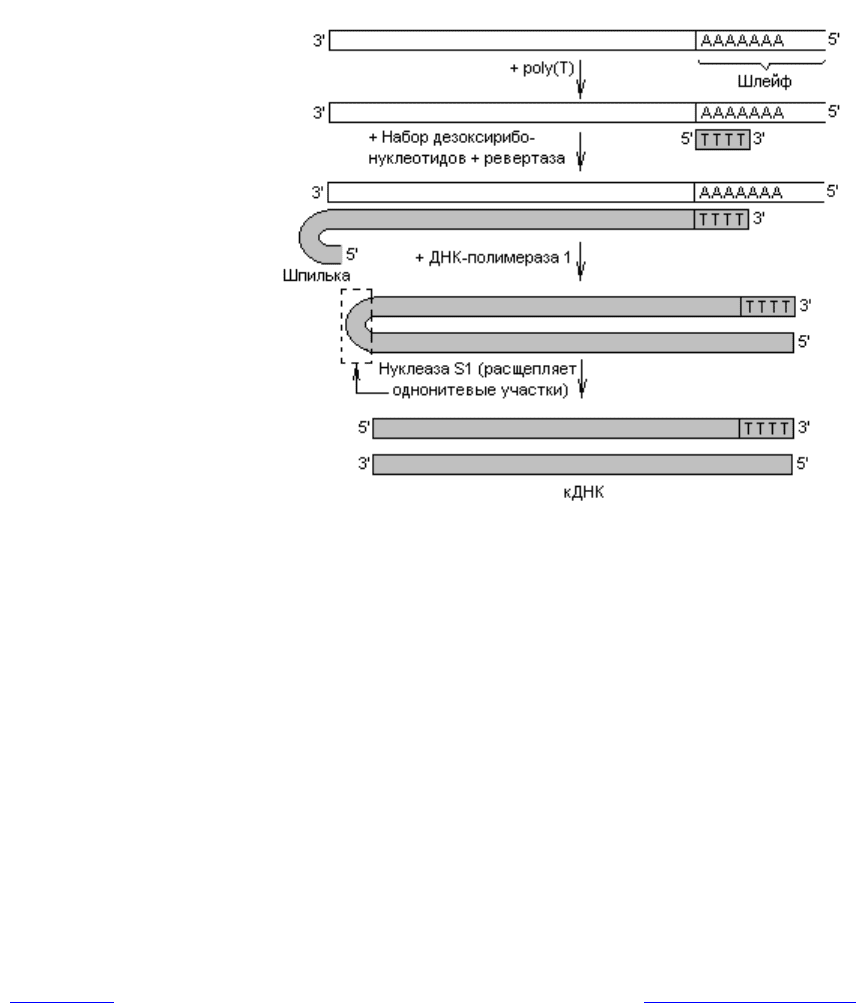
рицы, которой на первом этапе служит мРНК, выделенная из клетки. Практи-
чески все мРНК эукариот имеют на 3ў-конце «шлейф» (полиаденилатную по-
следовательность). Этот участок используют для образования затравки для
комплементарной цепи ДНК: к мРНК добавляют короткие последовательно-
сти, состоящие из тимидилатов, которые в результате отжига гибридизуются
с полиаденилатами (рис. 20.3). Обратная транскриптаза в присутствии набора
дезоксирибонуклеотидов катализирует их присоединение к затравке в после-
довательности, определяемой мРНК, в результате чего образуется двухните-
вый гибрид РНК-ДНК. По невыясненным пока причинам новосинтезирован-
ная цепь ДНК имеет на конце шпильку (рис. 20.3), которая возникает только
при реакции in vitro (по-видимому, из-за того, что фермент «поворачивает
вспять»). Эту шпильку используют в качестве затравки для синтеза вто-
Рис. 20.3. Образование кДНК в ходе ферментативного синтеза на
основе МРНК
рой цепи ДНК. На следующем этапе осуществляют деградацию РНК с помо-
щью рибонуклеаз либо в ходе щелочного гидролиза, а оставшуюся одноните-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)