Страйер Л. Биохимия. Том 3
Подождите немного. Документ загружается.

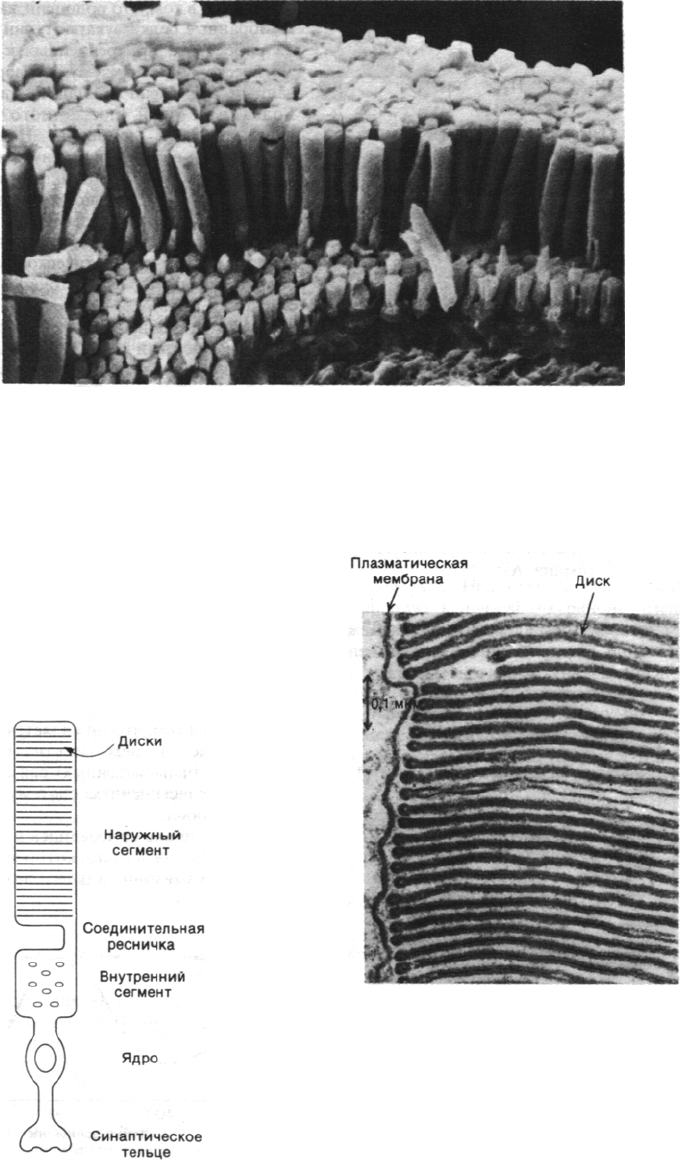
Рис. 37.21. Микрофотография клеток па-
лочек сетчатки, полученная
в сканирующем электронном
микроскопе. (Печатается с лю-
безного разрешения д-ра Deric
Bownds.)
(хромофора). Фоточувствительным веще-
ством палочек является родопсин, состоя-
щий из белка опсина и простетической
группы, представленной 11-цис-ретиналем
(рис. 37.24). Родопсин представляет собой
трансмембранный белок массой 38 кДа. Его
Рис. 37.22. Схематическое изображение
палочки сетчатки.
N-конец расположен в водной фазе внутри
диска, а С-конец - на другой стороне мем-
браны диска, в цитозоле. N-концевая
область родопсина содержит две олигосаха-
ридные единицы, ковалентно присоеди-
ненные к аспарагиновой боковой цепи.
Рис. 37.23. Наружный сегмент палочки
сетчатки под электронным ми-
кроскопом. Видны уложенные
стопкой диски. (Allen J. M., ed.,
Molecular organization and
biological function, Harper and
Row, 1967.)
37. Возбудимые мембраны
и сенсорные системы
341
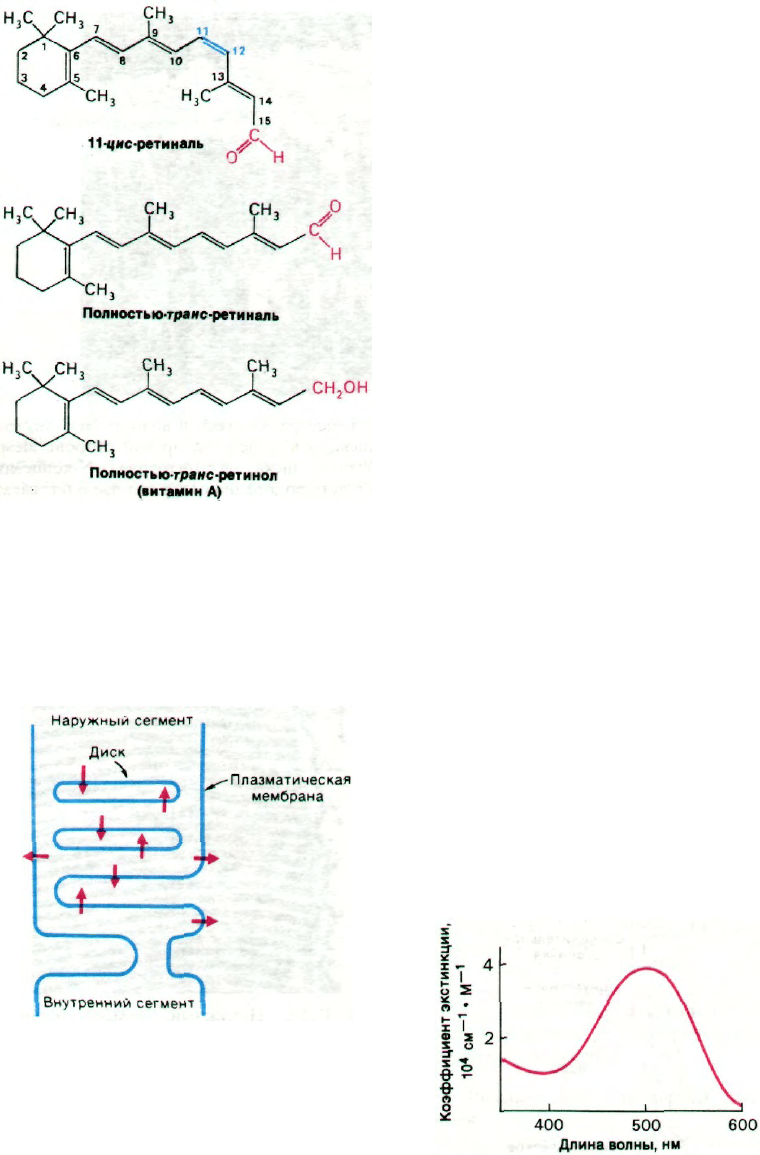
Рис. 37.24. Структура 11-цис-ретиналя, по-
лностью-транс-ретиналя и по-
лностью-транс-ретинола (ви-
тамина А).
Этим сахарам, по-видимому, принадлежит
важная роль в направленном передвижении
родопсина из внутреннего сегмента к ди-
Рис. 37.25. Образование дисков путем ин-
вагинации плазматической
мембраны. Стрелками показа-
на полярность молекул ро-
допсина.
342
Часть V.
Молекулярная физиология
скам. Дело в том, что родопсин, как и другие
мембранные белки эукариот, синтезируется
на рибосомах, прикрепленных к эндоплаз-
матическому ретикулуму. Новосинтезиро-
ванный родопсин попадает в аппарат
Гольджи и лишь после этого достигает
плазматической мембраны. Новые диски
образуются в основании внутреннего сег-
мента путем инвагинации плазматической
мембраны, и именно поэтому углеводные
единицы родопсина оказываются локализо-
ванными внутри диска, хотя первоначально,
входя в состав плазматической мембраны,
они обращены во внеклеточное простран-
ство (рис. 37.25).
Опсин, подобно другим белкам, ли-
шенным простетических групп, не погло-
щает видимого света. Цвет родопсина и его
чувствительность к свету определяются
присутствием 11-цис-ретиналя, являющего-
ся высокоэффективным хромофором. Бла-
годаря 11-цис-ретиналю родопсин обладает
широкой полосой поглощения в видимой
области спектра с максимумом при 500 нм,
что прекрасно соответствует солнечному из-
лучению. Примечательна также интенсив-
ность поглощения видимого света родопси-
ном. Коэффициент экстинкции родопсина
при 500 нм очень высок, а именно
4•10
4
см
-1
•М
-1
(рис. 37.26). Суммарная
сила поглощения видимого света родопси-
ном приближается к максимальным значе-
ниям для органических соединений. Высо-
кие хромофорные качества 11-цис-ретиналя
обусловлены тем, что он является полиеном.
Чередование в нем шести одинарных
и двойных (ненасыщенных) связей создает
длинную ненасыщенную систему для пере-
носа электрона.
11-цис-ретиналь присоединен к родопсину
через шиффово основание, которое образует-
ся при связывании альдегидной группы
Рис. 37.26. Спектр поглощения родоп-
сина.
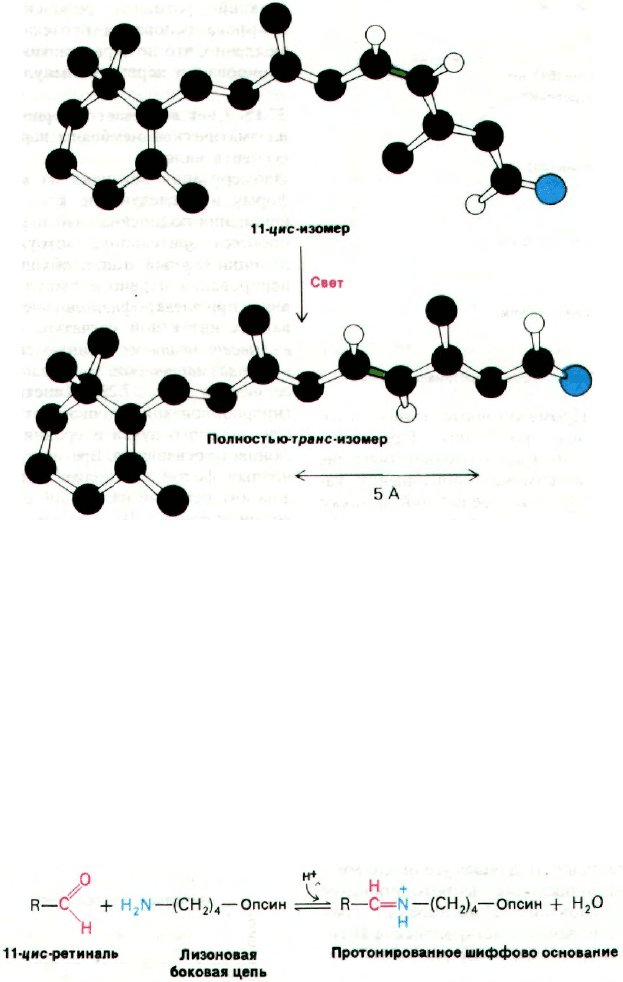
Рис. 37.27. Первичный акт при возбужде-
нии светом - это изомеризация
11-цис-изомера шиффова осно-
вания, образуемого ретиналем.
в полностью-транс-форму.
Двойная связь между С-11
и С-12 показана зеленым цве-
том.
11-цис-ретиналя с ε-аминогруппой специфи-
ческого остатка лизина в опсине. Спек-
тральные свойства родопсина свидетель-
ствуют о том, что шиффово основание нахо-
дится в протонированной форме.
Предшественником 11-цис-ретиналя слу-
жит полностью-транс-ретинол (витамин А),
который в организме млекопитающих не
может синтезироваться de novo. Пол-
ностью-транс-ретинол (рис. 37.24) превра-
щается в 11-цис-ретиналь в два этапа. Сна-
чала происходит окисление спиртовой
группы в альдегидную в присутствии рети-
нол-дегидрогеназы и NADP
+
в качестве ак-
цептора электронов. Затем под действием
ретиналь-изомеразы двойная связь между
С-11 и С-12 изомеризуется из транс-формы
в цис-форму. Недостаточность витамина
А приводит к ночной («куриной») слепоте
и в конечном итоге к повреждению на-
ружных сегментов палочек.
37.14. Свет вызывает изомеризацию
11-цис-ретиналя
Первичный акт в процессе зрительного воз-
буждения известен. Как показал Джордж
Уолд (George Wald), свет вызывает изомери-
зацию 11-цис-ретиналъной группы родопсина
в полностью-транс-ретиналь. В результате
изомеризации сильно меняется геометрия
ретиналя (рис. 37.27). Образованное ретина-
лем шиффово основание передвигается по
отношению к области кольца хромафора
примерно на 5 А. В сущности, поглощенный
фотон преобразуется в движение атомов.
Значительная изомеризация ретиналя
происходит практически в первые несколько
37. Возбудимые мембраны
и сенсорные системы
343
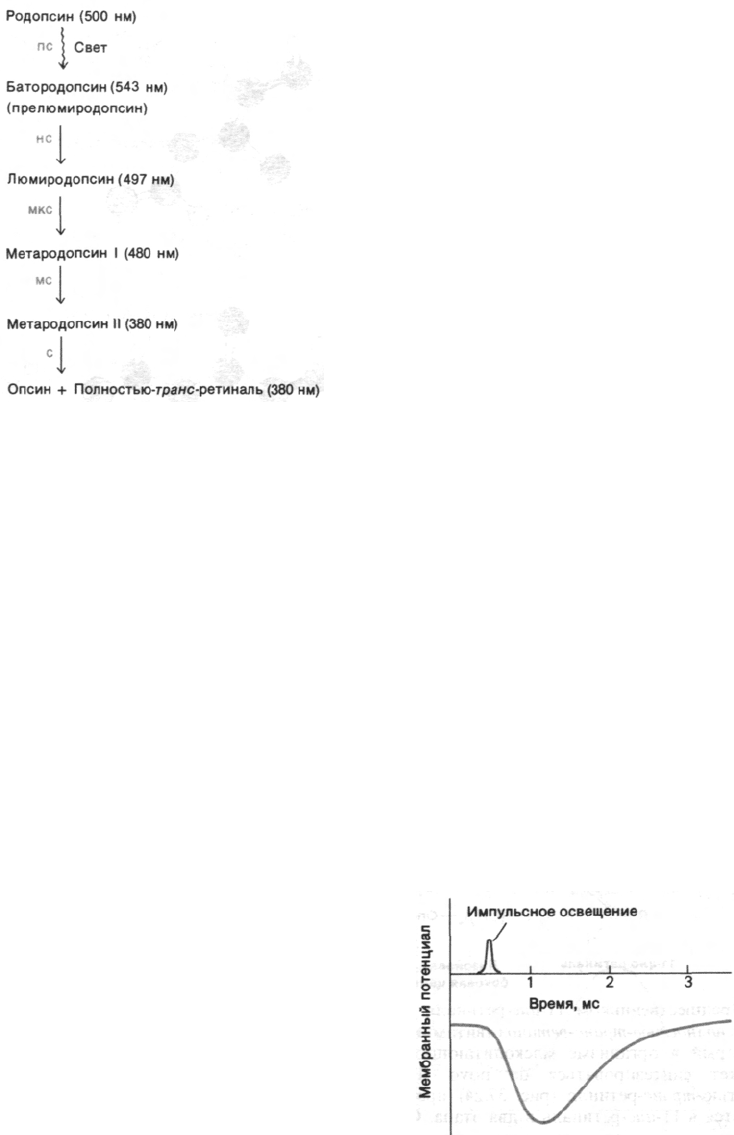
Рис. 37.28. Промежуточные этапы фото-
лиза родопсина. Приведены
длины волн, соответствующие
максимумам поглощения ка-
ждого из соединений, а также
постоянная времени каждого
из превращений.
пикосекунд после поглощения фотона, о чем
можно судить по появлению новой полосы
поглощения после сильного импульсного
освещения лазером. Образующийся при
этом промежуточный продукт фотолиза, на-
зываемый батородопсином (или прелюмиро-
допсином), содержит напряженную пол-
ностью-транс-форму хромофора. Далее
и ретиналь, и белок продолжает изменять
свою конформацию, что приводит к образо-
ванию ряда промежуточных продуктов, раз-
личающихся по спектральным свойствам
(рис. 37.28). При переходе от метародопсина
I к метародопсину II, длящемуся около мил-
лисекунды, происходит депротонирование
шиффова основания. Лишенное протона
шиффово основание в метародопсине II ги-
дролизуется в течение примерно одной ми-
нуты с образованием опсина и полностью-
транс-ретиналя; последний путем диффузии
отделяется от опсина, поскольку не соответ-
ствует участку связывания 11-цис-изомера.
Полностью-транс-ретиналь в темноте изо-
меризуется снова в 11-цис-ретиналь, ко-
торый связывается с опсином, и таким пу-
тем регенерируется родопсин. В отличие от
344
Часть V.
Молекулярная физиология
реакций фотолиза родопсина гидролиз
шиффова основания протекает настолько
медленно, что не играет никакой роли в ге-
нерировании нервного импульса.
37.15. Свет вызывает гиперполяризацию
плазматической мембраны наружного
сегмента палочек
Изомеризация ретиналя из цис- в транс-
форму и последующие конформационные
изменения родопсина - это первичные акты
процесса зрительного возбуждения. Сле-
дующий важный этап, необходимый для ге-
нерирования нервного импульса, был вы-
явлен при электрофизиологических исследо-
ваниях интактной сетчатки. Квант света
вызывает кратковременную гиперполяриза-
цию плазматической мембраны наружных
сегментов (рис. 37.29). Кинетика процесса
гиперполяризации зависит от интенсивно-
сти светового пучка и уровня устойчивого
фонового освещения. Время реакции на еди-
ничный фотон составляет около секунды,
а на интенсивный падающий свет - несколь-
ко миллисекунд. В палочках не возникает
потенциала действия. Их реакция на свет
может быть различной силы. Величина сиг-
нала, идущего от наружного сегмента
к синапсу, зависит от числа поглощенных
фотонов. Если взять обладающие полной
чувствительностью адаптированные к тем-
ноте палочки, то полумаксимальный уро-
вень гиперполяризации наблюдается при
поглощении всего лишь 30 фотонов на-
ружным сегментом палочки, содержащим
40•10
6
молекул родопсина (рис. 37.30). По-
глощение единичного фотона адаптирован-
ной к темноте палочкой вызывает гиперпо-
ляризацию порядка 1 мВ, что улавливается
синапсом и передается на другие нейроны
сетчатки. Исключительная чувствитель-
Рис. 37.29. Под действием света происхо-
дит гиперполяризация палочек
сетчатки.
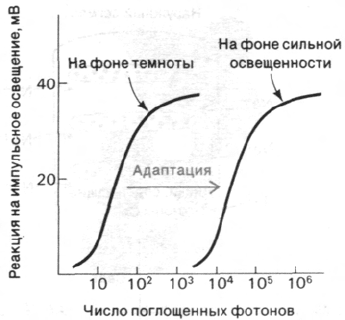
ность - не единственное выдающееся свой-
ство палочек. Их вторая поразительная осо-
бенность состоит в том, что реакция
световоспринимающего аппарата фотоде-
тектора на импульсное освещение зависит
от уровня фоновой освещенности. Для воз-
буждения палочек, освещенных постоянным
светом, требуется значительно больше фо-
тонов, чем для палочек, находящихся в тем-
ноте (рис. 37.30). Благодаря этому свойству,
называемому адаптацией, палочки сетчатки
способны функционировать при уровнях
фоновой освещенности, различающихся на
много порядков величины.
Каков ионный механизм индуцированной
светом гиперполяризации? Плазматическая
мембрана наружного сегмента палочки
в темноте высоко проницаема для Na
+
. Это
обстоятельство наряду с наличием высоко-
го трансмембранного градиента концентра-
ций Na
+
приводит к тому, что в темноте
ионы натрия быстро входят внутрь наруж-
ного сегмента. Этот градиент поддержи-
вается (Na
+
+ К
+
)-АТРазой, локализован-
ной в плазматической мембране внутренне-
го сегмента. Таким образом, в темноте
ионы натрия входят в наружный сегмент,
диффундируют далее во внутренний сег-
мент и затем выводятся с помощью насоса,
работающего за счет энергии АТР. Свет ка-
ким-то образом блокирует Na
+
-каналы
в плазматической мембране наружного сег-
мента. В результате ток Na
+
, направленный
внутрь клетки, снижается и мембрана с вну-
тренней стороны становится более элек-
троотрицательной. Другими словами, мем-
бранный потенциал палочек при освещении
сдвигается в сторону равновесного
К
+
-потенциала. Далее индуцированная све-
том гиперполяризация вблизи освещенных
дисков пассивно передается по плазматиче-
ской мембране на синаптическое тельце.
37.16. Медиаторы передают сигнал
от фотолизированного родопсина
на плазматическую мембрану
Изменение проницаемости плазматической
мембраны для Na
+
и последующая гипер-
поляризация - это многократно усиленные
реакции наружного сегмента на свет.
В самом деле, поглощение всего лишь одно-
го фотона адаптированной к темноте палоч-
кой блокирует ток более чем миллиона ио-
нов натрия. Как возникает усиление такого
масштаба? Прежде всего следует обратить
внимание на то, что мембраны дисков, со-
держащие основную массу молекул родоп-
Рис. 37.30. Чувствительность палочек сет-
чатки к импульсному освеще-
нию зависит от фонового уров-
ня освещенности.
сина, не соприкасаются с плазматической
мембраной палочек и не сопряжены с ней
электрически. Более того, молекула родоп-
сина, поглотившая фотон, может быть уда-
лена от натриевого канала плазматической
мембраны на несколько тысяч ангстрем. Все
это исключает возможность прямого взаи-
модействия между дисками и плазматиче-
ской мембраной. Практически не может
быть сомнений в том, что сигнал от фотоли-
зированного родопсина (Рд*) в мембранах
дисков передается на плазматическую мем-
брану с помощью диффундирующих медиа-
торов. При этом для обеспечения той высо-
кой степени усиления, которая реально
наблюдается, фотолиз одной молекулы ро-
допсина должен сопровождаться образова-
нием (или распадом) большого числа моле-
кул медиатора.
Природа медиатора не установлена еще
окончательно; проведенные исследования
позволили выдвинуть на эту роль двух до-
статочно вероятных кандидатов: ионы Са
2+
(рис. 37.31) и циклический GMP (рис. 37.32).
В
пользу
Са
2+
как
медиатора
свидетель-
ствуют следующие экспериментальные
данные.
1. Натриевые каналы в плазматической
мембране закрываются при повышении со-
держания Са
2+
в цитозоле и открываются
при его понижении.
2. При введении в цитозоль хелатирую-
37. Возбудимые мембраны
и сенсорные системы
345
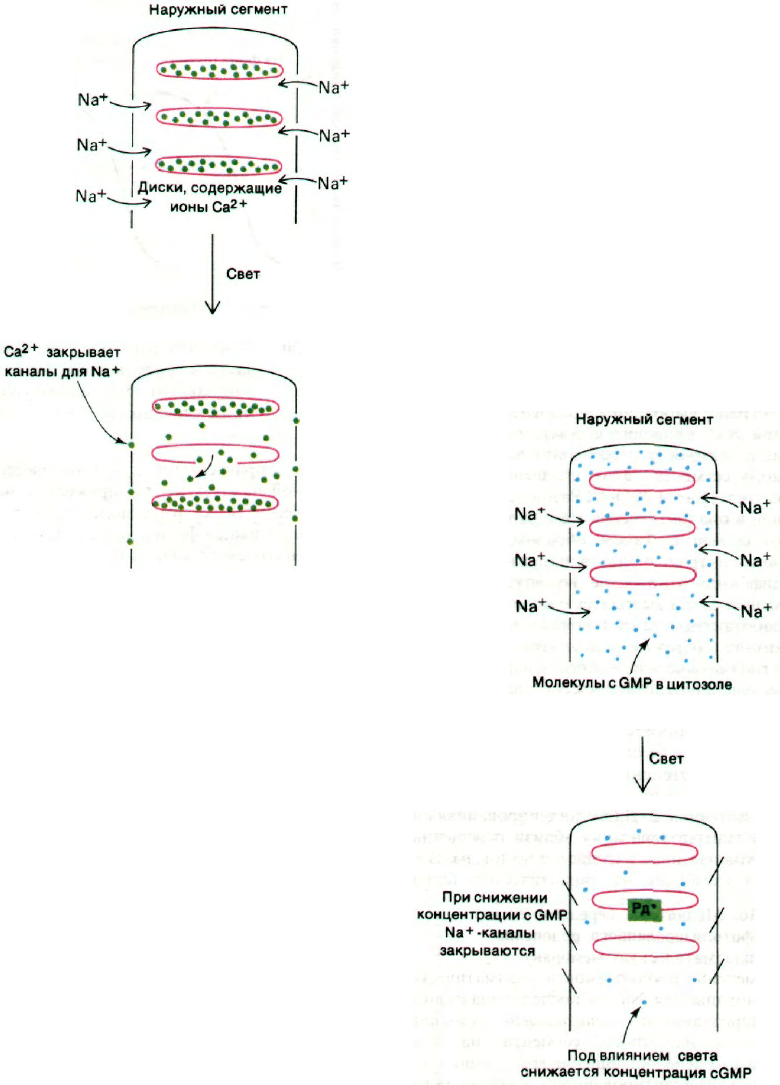
Рис. 37.31. Схематическое изображение
гипотезы о роли ионов кальция
как медиаторов при зритель-
ном возбуждении.
щих агентов, специфически связывающих
Са
2+
,
чувствительность палочек
к
свету
уменьшается. Такая десенсибилизация
указывает на то, что фотолиз одной моле-
кулы родопсина ведет к высвобождению
в цитозоль нескольких сотен ионов Са
2+
.
3. После импульсного воздействия све-
том из наружных сегментов палочек выво-
дится много Са
2+
.
Однако, судя по результатам других экс-
периментов, медиатором может быть
cGMP. Решающее значение для этого за-
ключения имеют следующие данные.
1. Натриевые каналы плазматической
мембраны открываются при повышении со-
держания cGMP в цитозоле и закрываются
при снижении содержания этого нуклеоти-
да.
346
Часть V.
Молекулярная физиология
2. Содержание cGMP регулируется све-
том. В частности, под действием света про-
исходит активация фосфодиэстеразы, ги-
дролизующей cGMP, что будет описано
несколько ниже.
3. Фотолиз одной молекулы родопсина
ведет к быстрому гидролизу 10
5
молекул
cGMP.
37.17. Свет снижает содержание
циклического GMP путем активации
фосфодиэстеразы
Как показывают приведенные эксперимен-
тальные данные, возбудимость палочек за-
висит и от Са
2+
, и от cGMP. Взаимодей-
ствие этих агентов может играть решаю-
щую роль в зрительном возбуждении. Что
касается молекулярного механизма индуци-
рованного светом высвобождения Са
2+
Рис. 37.32. Схематическое изображение
гипотезы о роли cGMP как
медиатора при зрительном
возбуждении. Рд* - фотолизи-
рованный родопсин.
в цитозоль, то в этом вопросе еще много не-
ясного. Что же касается регуляции светом
содержания cGMP в наружных сегментах
палочек, то в изучении этого вопроса в по-
следние годы был достигнут значительный
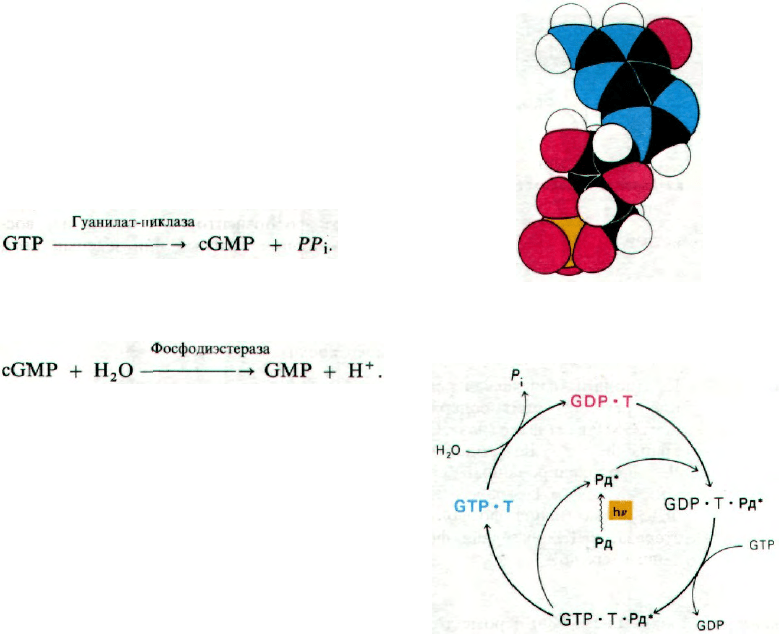
прогресс. На гуанилат-циклазу - фермент,
катализирующий синтез
cGMP,-
свет,
по-
видимому, не оказывает существенного
влияния:
Зато на фосфодиэстеразу, гидролизующую
cGMP, свет оказывает поразительное по си-
ле действие:
В результате освещения активность фосфо-
диэстеразы возрастает в несколько сотен
раз. Стимуляция этого фермента фотолизи-
рованным родопсином опосредована регу-
ляторным белком, называемым трансдуци-
ном. В темноте трансдуцин содержит про-
чно связанную молекулу GDP. При освеще-
нии фотолизированный родопсин образует
комплекс с GDP-трансдуцином и катализи-
рует обмен GDP на GTP (рис. 37.34). Возни-
кающий комплекс GTP-трансдуцин активи-
рует фосфодиэстеразу. Крайне важное зна-
чение имеет то обстоятельство, что всего
лишь одна фотолизированная молекула ро-
допсина катализирует обмен GDP на GTP
на нескольких сотнях молекул трансдуцина,
что в свою очередь активирует сотни моле-
кул фосфодиэстеразы. Следовательно, если
число оборотов фосфодиэстеразы соста-
вляет примерно 10
3
с
-1
, то на свету в тече-
ние секунды гидролизуется уже более 10
5
молекул cGMP в расчете на одну молекулу
фотолизированного родопсина. Система
возвращается в исходное темновое состоя-
ние благодаря встроенной в нее GTP-азной
активности. Присоединенный к трансдуцину
GTP подвергается медленному гидролизу
с образованием GDP-трансдуцина, неспо-
собного активировать фосфодиэстеразу. Та-
ким образом, весь этот цикл протекает за
счет свободной энергии гидролиза GTP.
Здесь мы видим пример того, как для усиле-
ния сигнала используется ~Р. Описанный
каскад реакций, регулирующих содержание
cGMP (рис. 37.35), очень напоминает каскад
реакций, опосредующих действие β-адре-
нергических гормонов, в частности адрена-
лина (разд. 35.4).
Рис. 37.33. Молекулярная модель цикли-
ческого GMP.
Рис. 37.34. Фотолизированный родопсин
(Рд*) катализирует образова-
ние комплекса GTP с трансду-
цином; этот комплекс в свою
очередь активирует фосфодиэ-
стеразу циклического GMP.
[Fung В. К.-К., Stryer L., Ргос.
Nat. Acad, Sci., 77, 2503 (1980).]
37.18. Цветовое зрение опосредуется
фоторецепторами трех типов
В 1802 г. Томас Юнг (Thomas Young) выска-
зал предположение, что цветовое восприя-
тие опосредовано тремя основными рецеп-
торами. Как показали спектрофотометриче-
ские исследования интактной сетчатки, про-
веденные более 150 лет назад, в глазу
существует три типа клеток-колбочек,
а именно клетки, поглощающие синий, зе-
леный и красный свет. Для получения спек-
тра поглощения этих трех фоторецепторных
пигментов колбочки освещали лучом света
37. Возбудимые мембраны
и сенсорные системы
347

Рис. 37.35. Предполагаемый каскад реак-
ций, регулирующих содержа-
ние cGMP в сетчатке глаза. Со-
кращения: Рд - родопсин,
Рд* - фотолизированный ро-
допсин, Т - трансдуцин,
ФДЭн - неактивная фосфодиэ-
стераза, ФДЭ - активная фос-
фодиэстераза.
диаметром I мкм (рис. 37.36). Кроме того,
во многие колбочки вводили микроэлек-
троды. Спектры действия, основанные на
гиперполяризации плазматических мем-
бран, разделяются на три группы с макси-
мумами соответственно в синей, зеленой
и красной областях видимого света. У золо-
той рыбки максимумы поглощения трех
цветовых рецепторов приходятся на 455, 530
и 625 нм, тогда как родопсин характери-
зуется максимумом 500 нм.
Хромофором в колбочках всех трех типов
служит 11-цис-ретиналъ. В отсутствие бел-
ка протонированное шиффово основание
имеет максимум поглощения при 380 нм.
Следовательно, различные группировки на
опсине оказывают значительное действие на
хромофорные свойства связанного 11-цис-
ретиналя. Зависимость спектральных
свойств этого хромофора от белкового
окружения - частное проявление общего
принципа, а именно взаимодействие с бел-
ком оказывает модулирующее влияние на
свойства простетической группы. Другой
пример тому - функционирование гема в ка-
348
Часть V.
Молекулярная физиология
честве переносчика кислорода в гемоглоби-
не, переносчика электронов в цитохроме с
и катализатора в пероксидазе.
Большинство форм дальтонизма (цвето-
вой слепоты) обусловлено сцепленной с по-
лом рецессивной мутацией. Около 1% му-
жчин не видят красного цвета, около
2% - зеленого. Как показали спектральные
исследования, проведенные на интактном
глазе, у этих людей либо совсем отсут-
ствуют фоторецепторные молекулы, вос-
принимающие красный или зеленый цвет,
либо имеется измененный пигмент со сдви-
нутым спектром поглощения. Таким обра-
зом, дальтонизм обусловлен отсутствием
или дефектом одного из типов опсина
в колбочках.
37.19. 11-цис-ретиналь - хромофор всех
известных органов зрения
Только у трех типов животных - моллю-
сков, членистоногих и позвоночных - глаза
способны отображать образ предмета. Ана-
томически глаза этих трех типов устроены
совершенно по-разному и, по-видимому,
в ходе эволюции возникли независимо. Од-
нако во всех трех случаях хромофором в фо-
торецепторных молекулах служит 11-цис-
ретиналь. Это поразительный пример кон-
вергентной эволюции. Что же такого особен-
ного в 11-цис-ретинале? Во-первых, это
соединение обладает интенсивной полосой
поглощения, которая легко сдвигается в ви-
димую область спектра. Во-вторых, под
действием света 11-цис-ретиналь легко изо-
меризуется. Более того, в темноте скорость
изомеризации очень низка. В-третьих, изо-
меризация вызывает большие изменения
в структуре. В итоге поглощенный свет пре-
образуется в движение атомов такого масш-
таба, которое способно инициировать
генерирование нервного импульса. На-
конец, исходными предшественниками
Рис. 37.36. Спектры поглощения трех цве-
товых рецепторов.
Рис. 37.37. Вид клеток - палочек и колбо-
чек - фоторецепторного слоя
сетчатки в сканирующем элек-
тронном микроскопе. (Печа-
тается с любезного разрешения
д-ра William Miller.)
11-цис-ретиналя являются каротины (разд.
20.27), очень широко распространенные
в живой природе.
37.20. Хеморецепторы бактерий восприни-
мают специфические молекулы
и передают сигналы на жгутики
В конце XIX в. немецкий ботаник Виль-
гельм Пфеффер (Wilhelm Pfeffer) продемон-
стрировал, что подвижные бактерии скапли-
ваются вокруг отверстия тонкого капилля-
ра, содержащего какой-нибудь аттрактант,
например сахар (рис. 37.38). В случае же,
когда капилляр содержит репеллент (обыч-
но это вещество, повреждающее бактерии,
или продукт их выделения), бактерии дви-
жутся прочь от капилляра. Такое направлен-
ное движение бактерий в сторону одних ве-
ществ и прочь от других называется
хемотаксисом. В 60-х годах Джулиус Адлер
(Julius Adler) начал изучать молекулярную
основу хемотаксиса бактерий. Выполненные
им, а также большим числом других ученых
биохимические, генетические и структурные
исследования раскрыли многие стороны
этого процесса. Хемотаксис начинается с об-
наружения химических соединений специфи-
ческими хеморецепторами на поверхности
клетки. Информация от этих сенсоров пере-
дается в систему преобразования, где проис-
ходит анализ и интеграция большого числа
стимулов. Далее сенсорная преобразующая
система посылает сигналы к моторам, при-
водящим в движение жгутики. В зависимо-
сти от этих сигналов бактерия либо плавно
передвигается по прямой, либо резко меняет
направление движения.
У Е. coli обнаружено около 20 различных
хеморецепторов. Каждый из этих белков ло-
кализован или в плазматической мембране,
или в периплазматическом пространстве.
Хеморецептор содержит узнающий и сигна-
37. Возбудимые мембраны
и сенсорные системы
349
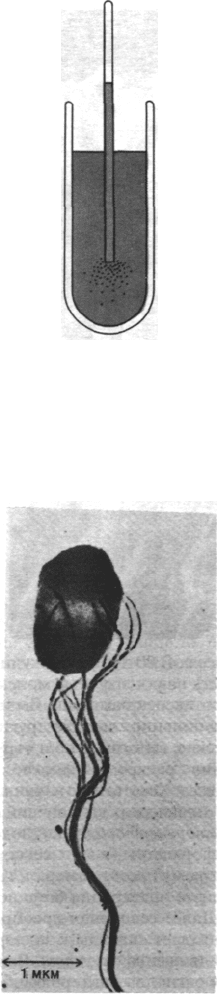
Рис. 37.38. Хемотаксис у бактерий. Бакте-
рии движутся к капилляру, со-
держащему атрактант, напри-
мер глюкозу.
Рис. 37.39. Электронная микрофотогра-
фия S. typhimurium. Хорошо
видно, что жгутики собраны
в пучок. (Печатается с любезно-
го разрешения д-ра Daniel
Koshland.)
350
Часть V.
Молекулярная физиология
лизирующий компоненты. В хеморецепто-
рах, обеспечивающих положительный хемо-
таксис (привлечение), узнающий компонент
оказался связывающим белком, участвую-
щим в транспорте данного соединения
в клетку. Так, например, связывающий га-
лактозу белок (растворимый белок, локали-
зованный в периплазматическом простран-
стве) служит и узнающим компонентом при
положительном хемотаксисе на галактозу,
и частью насоса, осуществляющего ак-
тивный транспорт галактозы в клетку. Хе-
морецептор для глюкозы является также
компонентом фосфотрансферазной си-
стемы, связанной с мембраной и обеспечи-
вающей активное потребление этого сахара
(разд. 36.12). На поверхности Е. coli имеются
хеморецепторы и для таких аттрактантов,
как серин, цистеин, аланин, глицин. Хотя
транспорт и хемотаксис тесно связаны ме-
жду собой, однако хемотаксис не зависит от
процессов транспорта. Так, некоторые му-
танты, неспособные транспортировать
определенные сахара или аминокислоты, со-
храняют способность направленно двигать-
ся к ним. Имеются также различные хеморе-
цепторы, связанные с отрицательным хемо-
таксисом. Жирные кислоты, спирты, гидро-
фобные аминокислоты, индол, Н
+
(рН <
< 6,5), ОН
-
(рН > 7,5) и сульфиды отталки-
вают бактерии путем взаимодействия со
специфическими хемосенсорами.
37.21. В основании бактериального жгутика
находится вращающий его реверсивный
«мотор»
Бактерии плывут благодаря вращению жгу-
тиков, отходящих от поверхности клетки.
Эти тонкие спиральные нити состоят из
субъединиц массой 53 кДа, называемых
флагеллином. У бактерии Е. coli имеется
около 6 жгутиков длиной 10 мкм и диаме-
тром 150 А. Жгутики бактерий по сравне-
нию со жгутиками и ресничками эукариот
(разд. 34.18) имеют значительно меньшие
размеры и проще устроены. Бактериальный
жгутик сам по себе не может совершать ак-
тивных волнообразных движений, так как
в нем нет сократительного аппарата. Его
вращает «мотор», расположенный в участке
соединения жгутика с клеточной оболочкой.
Выделение жгутиков, сохраняющих прикре-
пленную к ним базальную структуру, позво-
лило изучить эти образования; оказалось,
что они состоят из нити, крючка и стерж-
ня. У E.coli на стержень насажены 4 коль-
ца. Наружное кольцо прикреплено к наруж-