Страйер Л. Биохимия. Том 3
Подождите немного. Документ загружается.

перстянка... Наперстянка влияет на бие-
ние сердца в большей степени, чем
какое-либо из других лекарств, и это дей-
ствие можно с успехом использовать для
исцеления больного».
36.9. Транспорт кальция осуществляется
другой АТРазой
Ионы кальция играют важную роль в регу-
ляции мышечного сокращения (разд. 34.10),
а также во многих других физиологических
процессах. В скелетной мышце содержится
сложная сеть связанных с мембраной трубо-
чек и везикул. Эта мембранная система, на-
зываемая саркоплазматическим ретикулу-
мом, регулирует концентрацию Са
2+
в среде, окружающей сократительные мы-
шечные волокна. В состоянии покоя Са
2+
насасывается в саркоплазматический рети-
кулум, так что непосредственно вокруг мио-
фибрил концентрация Са
2+
бывает очень
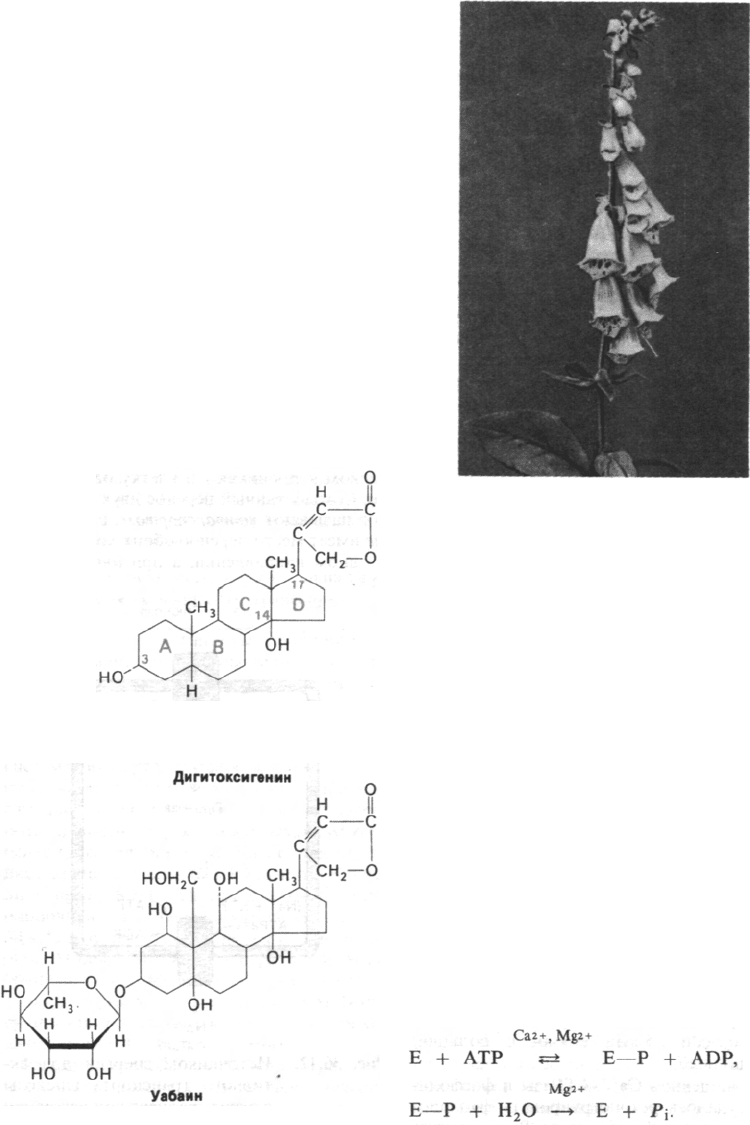
Рис. 36.9. Кардиотонические стероиды,
например дигитоксигенин и уа-
баин, ингибируют (Na
+
+
+ К
+
)-насос.
Рис. 36.10. Наперстянка. (Krochmal A.,
Krochmal С., A Guide to the
Medicinal Plants of the United
States, Quadrangle Books, 1973,
p.
243.)
низкой. Возбуждение мембраны саркоплаз-
матического ретикулума под влиянием не-
рвного импульса ведет к мгновенному выс-
вобождению больших количеств Са
2+
, и это
запускает мышечное сокращение. Другими
словами, Са
2+
служит промежуточным
звеном между нервным импульсом и сокра-
щением мышечного волокна.
Транспорт Са
2+
через мембрану сарко-
плазматического ретикулума происходит за
счет энергии АТР. В саркоплазматическом
ретикулуме имеется АТРаза, которую акти-
вирует Са
2+
. Эта Са
2+
-АТРаза является со-
ставной частью Са
2+
-насоса подобно тому,
как (Na
+
+ К
+
)-АТРаза - часть (Na
+
+
+ К
+
)-насоса. Са
2+
-АТРаза также подвер-
гается фосфорилированию в ходе гидролиза
АТР:
36. Мембранный транспорт
311
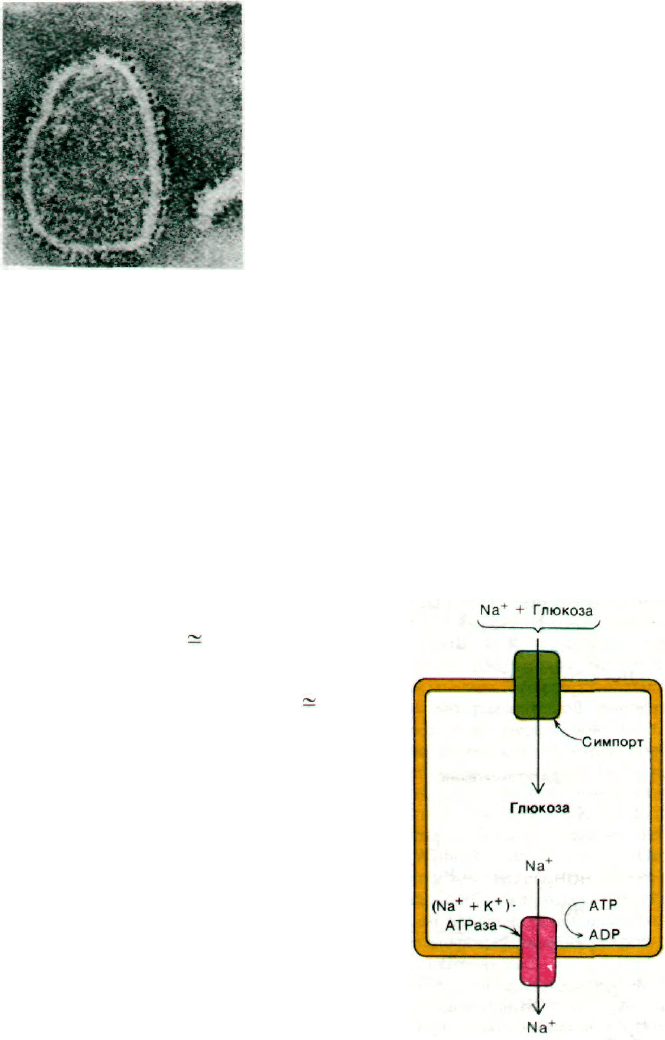
Рис. 36.11. Мембранные везикулы, обра-
зованные из очищенной
Са
2+
-АТРазы. Глобулярные
частицы на поверхности мем-
браны - участки молекулы
АТРазы, пронизывающей мем-
брану. [Stewart P. S., Мас-
Lennan D. H., J. Bioi.Chem., 249,
987 (1974).]
Цикл конформационных изменений, обус-
ловленных фосфорилированием и дефосфо-
рилированием, обеспечивает перенос двух
ионов Са
2+
при расщеплении одной моле-
кулы АТР. Благодаря очень высокому срод-
ству этой АТРазы к Са
2+
(К 10
-7
М) фер-
мент эффективно транспортирует Са
2+
из
цитозоля
(где
[Са
2+
]
<
10
-5
М) в
сарко-
плазматический рстикулум (где
[Са
2+
]
10
-2
М).
Плотность молекул Са
2+
-насоса в мем-
бране саркоплазматического ретикулума
очень велика, а именно около 20 000 в расче-
те на 1 мкм
2
. В сущности, Са
2+
-АТРаза со-
ставляет более 80% общего количества ин-
тегральных белков мембраны и занимает
треть ее поверхности. Большая субъединица
(100 кДа) Са-насоса пронизывет мембрану
и содержит участок фосфорилирования; как
и в (Na
+
+ K
+
)-насосе, таким участком
является специфическая боковая цепь, пред-
ставленная остатком аспартата. Еще одна
общая черта обоих насосов - это наличие
гликопротеина: в Са
2+
-насосе гликопро-
теин массой 55 кДа связан с большой
субъединицей.
Из очищенной Са
2+
-АТРазы и фосфоли-
пидов удалось реконструировать функцио-
нально активный Са
2+
-насос. В этом экспе-
312
Часть V.
Молекулярная физиология
рименте Са
2+
-АТРазу выделяли из мем-
бран саркоплазматического ретикулума по-
сле их солюбилизации детергентом хола-
том. Очищенный солюбилизированный
фермент добавляли к фосфолипидам из со-
евых бобов. После удаления детергента пу-
тем диализа образовывались мембранные
везикулы. Эти реконструированные вези-
кулы с высокой скоростью накапливали Са
2+
в присутствии АТР и Мg
2+
.
36.10. Поток Na
+
обеспечивает энергией
активный транспорт сахаров и аминокислот
в животных клетках
Многие транспортные процессы не зависят
непосредственно от гидролиза АТР, а со-
пряжены с потоком ионов по электрохими-
ческому градиенту. Так, во многих жи-
вотных клетках насасывание глюкозы обес-
печивается одновременным входом Na
+
.
При этом ионы натрия и глюкоза связы-
ваются со специфическим транспортным
белком и проникают в клетку одновремен-
но. Согласованный перенос двух компонен-
тов называют котранспортом; при симпор-
те имеет место перенос обоих компонентов
в одном направлении, а при антипорте - в
Рис. 36.12. Источником энергии для ак-
тивного транспорта глюкозы
служит градиент концентрации
Na
+
. Эта система симпорта
свойственна плазматическим
мембранам клеток кишечника
и почек.
противоположных направлениях. Ионы на-
трия, которые входят в клетку вместе с мо-
лекулами глюкозы посредством симпорта,
выводятся из клетки (Na
+
+ К
+
)-АТРазой
(рис. 36.12). Количество транспортируемой
глюкозы и скорость ее транспорта зависят
от трансмембранного градиента концентра-
ции
Na
+
.
Зависящий от Na
+
симпорт широко ис-
пользуется в животных клетках для нако-
пления аминокислот. В некоторых клетках,
например в микроворсинках щеточной
каемки кишечника (рис. 36.13), посредством
симпорта осуществляется активный транс-
порт сахаров. Кроме того, в тонком кишеч-
нике существует специализированный
Na
+
-зависимый симпорт, обеспечивающий
перенос ионов Cl
-
против градиента кон-
центрации. Во многих клетках ионы натрия
служат также движущей силой в процес-
сах антипорта, направленных на выведение
ионов кальция. Таким образом, градиент
концентрации ионов натрия, создаваемый
(Na
+
+
K
+
)-АТРазой,
обеспечивает
энер-
гией большинство симпортов и антипортов
в животных клетках.
36.11. Поток протонов служит движущей
силой во многих процессах транспорта
у бактерий
Симпорты и антипорты - эволюционно
очень древние механизмы молекулярного
транспорта. Так, движущей силой многих
транспортных систем у бактерий служит по-
ток протонов через плазматическую мем-
брану. Наиболее изученная бактериальная
система симпорта - перенос лактозы у E.coli
(рис. 36.14). У этой обитательницы нижних
отделов кишечника млекопитающих выра-
ботался высокоэффективный механизм кон-
центрирования лактозы. Насос для этого
дисахарида, выделенный Юджином Кенне-
ди (Eugene Kennedy), представляет собой
одиночную полипептидную цепь массой
30 кДа и называется пермеазой для лактозы
(или М-белком). Это интегральный мем-
бранный белок, который кодируется геном
у, входящим в lac-оперон (разд. 28.3). В ин-
дуцированных клетках на его долю прихо-
дится около 4% белков мембраны.
Раскрытию механиза функционирования
лактозного насоса способствовало изучение
мутантов по гену у, а также исследования
везикул (пузырьков), полученных из бакте-
риальных мембран. Везикулы очень удобны
для изучения процессов транспорта, по-
скольку они значительно проще устроены,
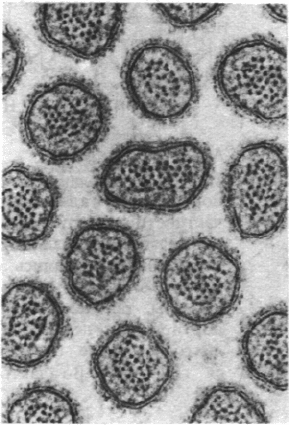
Рис. 36.13. Электронная микрофотогра-
фия поперечного среза микро-
ворсинок кишечника. Наличие
микроворсинок во много раз
увеличивает площадь поверх-
ности, через которую происхо-
дит транспорт питательных ве-
ществ. (Печатается с любезно-
го разрешения д-ра G. Pallade.)
чем целая бактерия. В везикулах есть систе-
ма окислительного фосфорилирования и
другие связанные с мембраной белки, но от-
сутствуют цитоплазматические компоненты
интактной клетки. Сами по себе везикулы не
накапливают лактозы, но если добавить
субстрат окисления, обеспечивающий поток
обладающих высоким потенциалом элек-
тронов по дыхательной цепи, то накопление
лактозы имеет место. Этот же эффект мож-
но получить и иным способом, а именно со-
зданием градиента рН с помощью образуе-
мой вне клеток кислоты. Создание мем-
бранного потенциала градиентом концен-
трации К
+
также приводит к насасыванию
лактозы. Все эти данные показывают, что
активный транспорт лактозы обеспечивает-
ся протонодвижущей силой в плазматиче-
ской мембране. Транспорт молекул лактозы
сопряжен с движением протона в клетку.
При физиологических условиях протонный
36. Мембранный транспорт
313
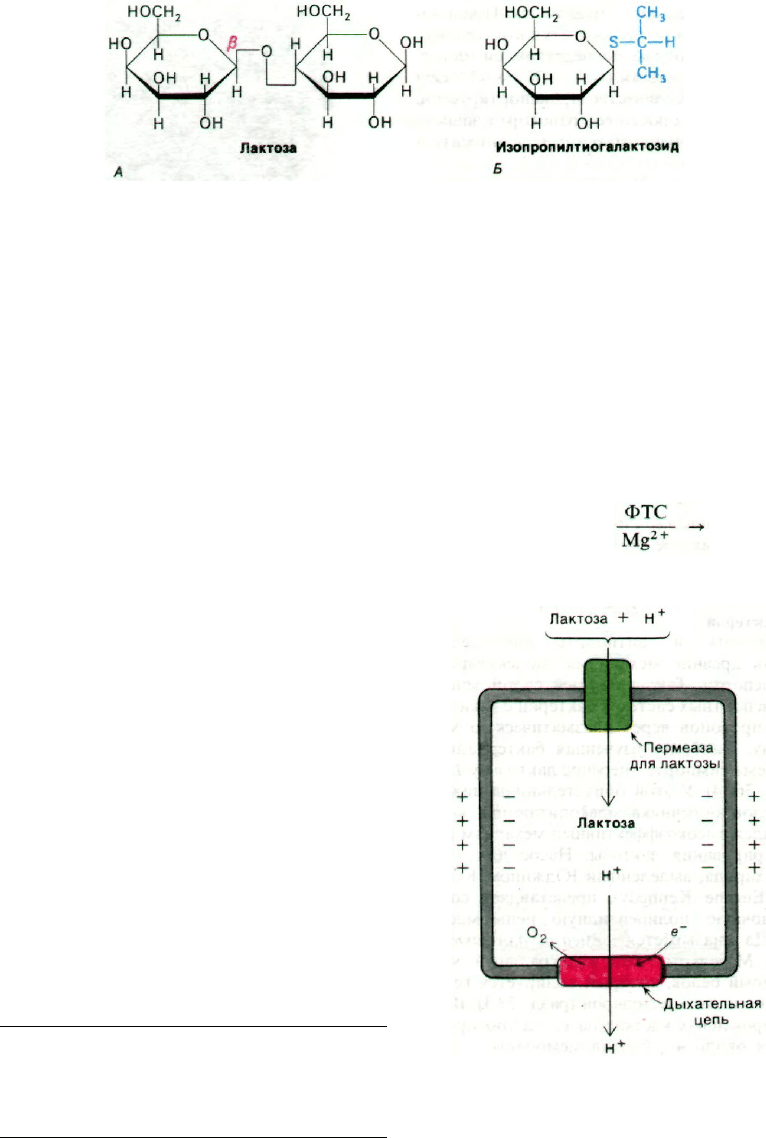
Рис. 36.14. Пермеаза для лактозы транс-
портирует β-галактозиды. в
частности лактозу (А) и изо-
пропилтиогалактозид (Б). Тио-
галактозиды оказались очень
удобными для изучения этой
системы, так как они транс-
портируются пермеазой для
лактозы, но не подвергают-
ся гидролизу β-галактозида-
зой.
градиент, необходимый для этого активно-
го транспорта, возникает за счет потока
электронов от донора, обладающего высо-
ким потенциалом (например, NADH), по
дыхательной цепи. Симпорт протонов и лак-
тозы иллюстрирует обобщающую концеп-
цию Питера Митчелла (Peter Mitchell)
о «преобразовании энергии посредством
протонного градиента» (разд. 14.18).
36,12. Активный транспорт ряда сахаров
сопряжен с их фосфорилированием
Симпорт - не единственный тип насосов,
осуществляющих транспорт сахаров. У не-
которых бактерий накопление углеводов
происходит путем сопряжения их входа
в клетку с фосфорилированием. Например,
у многих бактерий поступающая в клетки
глюкоза превращается в глюкозо-6-фосфат.
Особенность транспорта этого типа, назы-
ваемого транслокацией группы, состоит
в том, что в ходе транспорта происходит
модификация растворенного вещества. Кле-
точная мембрана непроницаема по отноше-
Таблица 36.1. Углеводы, транспортируемые фосфо-
трансферазной системой Е. coli
Глюкоза
Фруктоза
Манноза
N-ацетилглюкозамин
Маннитол
Сорбитол
Галактитол
Лактоза
314
Часть V.
Молекулярная физиология
нию к фосфорилированным сахарам, и по-
тому они накапливаются внутри бакте-
риальной клетки.
Наиболее хорошо изучен процесс транс-
локации групп, осуществляемый фосфо-
трансферазной системой (ФТС) которую
открыл Сол Роузман (Saul Roseman). Осо-
бенность этой системы состоит в том, что
донором фосфорильной группы служит фос-
фоенолпируват, а не АТР или какой-либо
иной нуклеозидтрифосфат. Суммарная ре-
акция, катализируемая фосфотрансфераз-
ной системой, следующая:
Сахар
вне клетки
+
+ Фосфоенолпируват
—>
Фосфорилированный сахар
в
клетке
+
+ Пируват.
Рис. 36.15. Протонный градиент служит
источником энергии для транс-
порта ряда сахаров и амино-
кислот в бактериальные клет-
ки. Протонный градиент гене-
рируется током электронов
в дыхательной цепи.
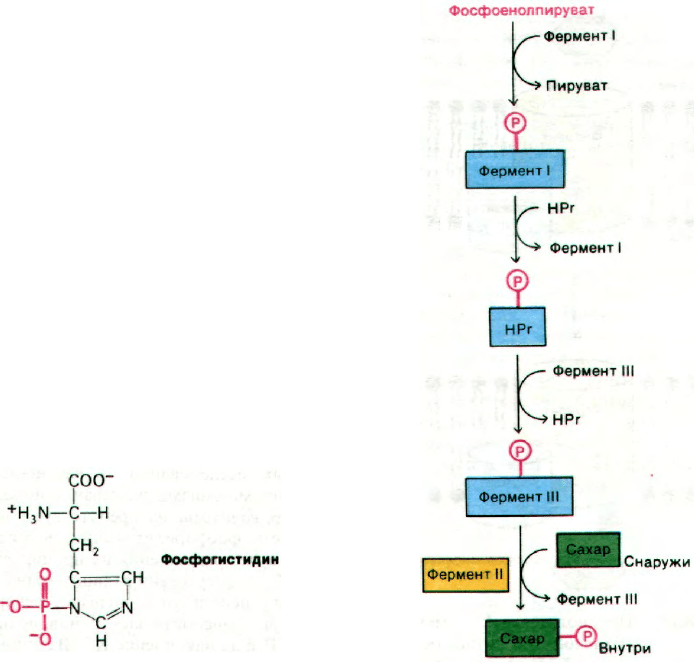
В этой транслокации участвуют 4 белка:
HPr, фермент I, фермент II и фермент III.
Фермент II, будучи интегральным белком
мембраны, образует трансмембранный ка-
нал и катализирует фосфорилирование саха-
ра. При этом фосфорильная группа фосфое-
нолпирувата переносится на сахар не прямо,
а сначала на фермент I и после на специфи-
ческий остаток гистидина небольшого
термостабильного белка HPr (рис. 36.16).
Образующийся в качестве промежуточного
соединения фосфогистидин имеет высокий
потенциал переноса фосфатной группы.
промежуточный по величине между со-
ответствующими значениями для АТР
и фосфоенолпирувата. Далее происходит
перенос фосфорильной группы от фосфори-
лированного HPr на фермент III - перифе-
рический мембранный белок, который уже
взаимодействует с собственно каналом, т. е.
ферментом II:
Конечный этап - перенос фосфорильной
группы от фермента III на транспорти-
руемый сахар (рис. 36.17). Из указанных
четырех очищенных белков удалось рекон-
струировать функционально активный фер-
ментный комплекс.
Некоторые белки, входящие в состав фос-
фотрансферазной системы, обладают специ-
фичностью, другие - нет. Так, HPr и фер-
мент I, которые являются растворимыми
белками цитозоля, участвуют в транспорте
всех сахаров, переносимых этой системой.
С другой стороны, ферменты II и III про-
являют специфичность в отношении опреде-
ленных сахаров. Например, в транспорте
глюкозы, лактозы и фруктозы участвуют
разные ферменты II и III. Такой же резуль-
тат был получен и при генетических иссле-
дованиях. У мутантов, дефектных по HPr
или ферменту I, не происходит транспорта
большого числа разных сахаров, тогда как
мутанты, дефектные по синтезу ферментов
II и III, не способны транспортировать
только какой-либо определенный сахар.
Фермент III не участвует в транслокации
гекситолов, в частности галактитола; в этом
Рис. 36.16. Поток фосфорильных групп от
фосфоенолпирувата на сахар,
транспортируемый через мем-
брану фосфотрансферазной си-
стемой.
случае фосфорильная группа переносится
непосредственно от HPr на углевод.
Чем объяснить, что фосфотрансферазная
система устроена значительно сложнее дру-
гих переносчиков, например пермеазы для
лактозы? Представляется вероятным, что
фосфотрансферазная система не только осу-
ществляет транспорт сахаров, но и выпол-
няет регуляторные функции. Избыточное
поступление какого-то одного углевода под
действием фосфотрансферазной системы
сильно подавляет активный транспорт саха-
ров другими переносчиками. Это ингибиро-
вание опосредуется, по-видимому, измене-
нием содержания сАМР, а именно повыше-
36. Мембранный транспорт
315
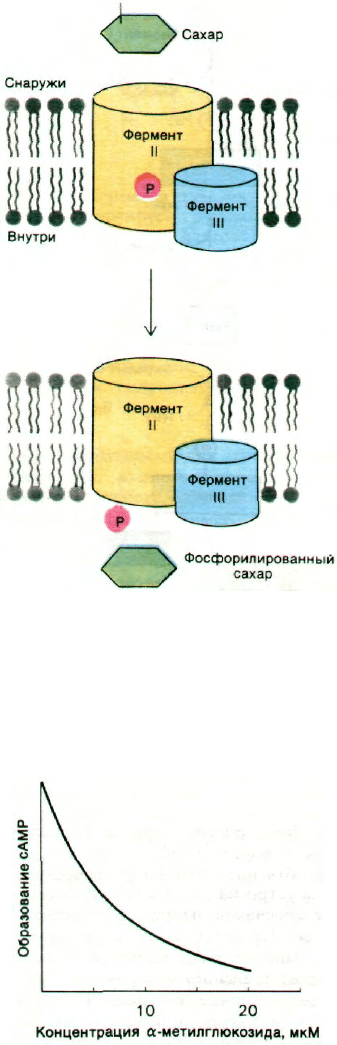
Рис. 36.17. Предполагаемый механизм
транслокации групп, осущест-
вляемой фосфотрансферазной
системой.
Рис. 36.18. Транспортируемый фосфо-
трансферазной системой сахар
α-метилглюкозид ингибирует
образование сАМР.
316
Часть V.
Молекулярная физиология
ние концентрации сахара, накопленного
фосфотрансферазной системой, ведет к сни-
жению выработки сАМР (рис. 36.18). В ре-
зультате прекращается транскрипция ряда
индуцируемых оперонов. Вспомним, что
экспрессия таких индуцируемых оперонов,
как lac и gal, значительно возрастает при
связывании комплекса сАМР с белком БАК
в соответствующих участках промотора
(разд. 28.6). Следовательно, фосфотрансфе-
разная система регулирует использование
источников углерода.
36.13. Транспортные антибиотики повышают
ионную проницаемость мембран
Ряд микроорганизмов синтезирует низко-
молекулярные соединения, в присутствии
которых мембраны становятся прони-
цаемыми для определенных ионов. Эти не-
большие молекулы, называемые транс-
портными антибиотиками, оказались
ценным инструментом для эксперимен-
тальных исследований, в частности для
изучения механизма связывания ионов. На-
пример, валиномицин препятствует окисли-
тельному фосфорилированию в митохон-
дриях путем повышения их проницаемости
для К
+
: в присутствии валиномицина мито-
хондрии используют энергию, генерируе-
мую при транспорте электронов, не на син-
тез АТР, а на накопление К
+
. Валиномицин
имеет циклическую структуру, образован-
ную из повторяющейся три раза последова-
тельности четырех разных остатков (А, Б,
В и Г) (рис. 36.19). Эти остатки четырех ти-
пов соединены чередующимися эфирными
и пептидными связями.
Еше один хорошо изученный транспорт-
ный антибиотик - грамицидин А (рис. 36.20).
Это полипептид с открытой цепью, сос-
тоящий из 15 аминокислотных остатков.
Примечательна структура грамицидина А:
в нем чередуются D- и L-аминокисло-
ты. Кроме того, N- и С-концы поли-
пептида модифицированы. Как будет пока-
зано несколько ниже, транспорт ионов
грамицидином А и валиномицином осу-
ществляется совершенно по-разному.
Механизм переноса ионов этими анти-
биотиками удобно исследовать на таких хо-
рошо охарактеризованных модельных си-
стемах, как двуслойные липидные пузырьки
(разд. 10.6) и плоские двуслойные мем-
браны (разд. 10.6). В опытах используют
пузырьки, содержащие радиоактивный ион,
например
42
К
+
; их получают путем обра-
ботки мембран ультразвуком в присутствии

Рис. 36.19. Валиномицин имеет периоди-
ческую циклическую структу-
ру, состоящую из остатков
L-лактата (А), L-валина (Б),
D-гидроксиизовалерианата (В)
и D-валина (Г).
этого
иона
и
последующего удаления
42
К
+
,
не попавшего внутрь пузырьков, методом
гель-фильтрации. Далее сравнивают ско-
рость выхода радиоактивного иона из пузы-
рьков в присутствии и в отсутствие антибио-
тика. Другой способ получения информации
относительно ионной проницаемости дву-
слойных мембран состоит в измерении та-
ких электрических параметров, как сопроти-
вление мембраны и мембранный потенциал.
Например, сопротивление двуслойной мем-
браны по отношению к К
+
в присутствии
10
-7
М валиномицина или 10
-9
М грами-
цидина падает более чем в 10000 раз при
градиенте концентраций КСl на мембране
0,02
М.
Рис. 36.20. Структура грамицидина А.
36.14. Транспортные антибиотики
функционируют либо как подвижные
переносчики, либо как каналообразователи
Существует два совершенно разных меха-
низма действия транспортных антибиоти-
ков на проницаемость мембран для ионов
(рис. 36.21). Некоторые антибиотики (на-
пример, грамицидин А) формируют канал,
пронизывающий мембрану. Ионы входят
в такой канал на одной стороне мембраны,
диффундируют по нему и выходят на дру-
гой стороне мембраны. Стимуляция транс-
порта ионов по этому механизму не сопря-
жена с движением самого антибиотика-ка-
налообразователя. Антибиотики другой
группы (например, валиномицин) функцио-
нируют как переносчики ионов через углево-
дородную область мембраны. Активность
этих транспортных антибиотиков сопряже-
на с их собственной диффузией.
В эксперименте подвижные переносчики
и каналообразователи можно различить сле-
дующим образом. Измеряют температур-
ную зависимость ионной проводимости ис-
кусственной липидной двуслойной мем-
Рис. 36.21. Схематическое изображение
различий между транспортны-
ми антибиотиками-канало-
образователями и подвижны-
ми переносчиками. Все из-
вестные транспортные белки
принадлежат к категории кана-
лообразователей.
36. Мембранный транспорт
317
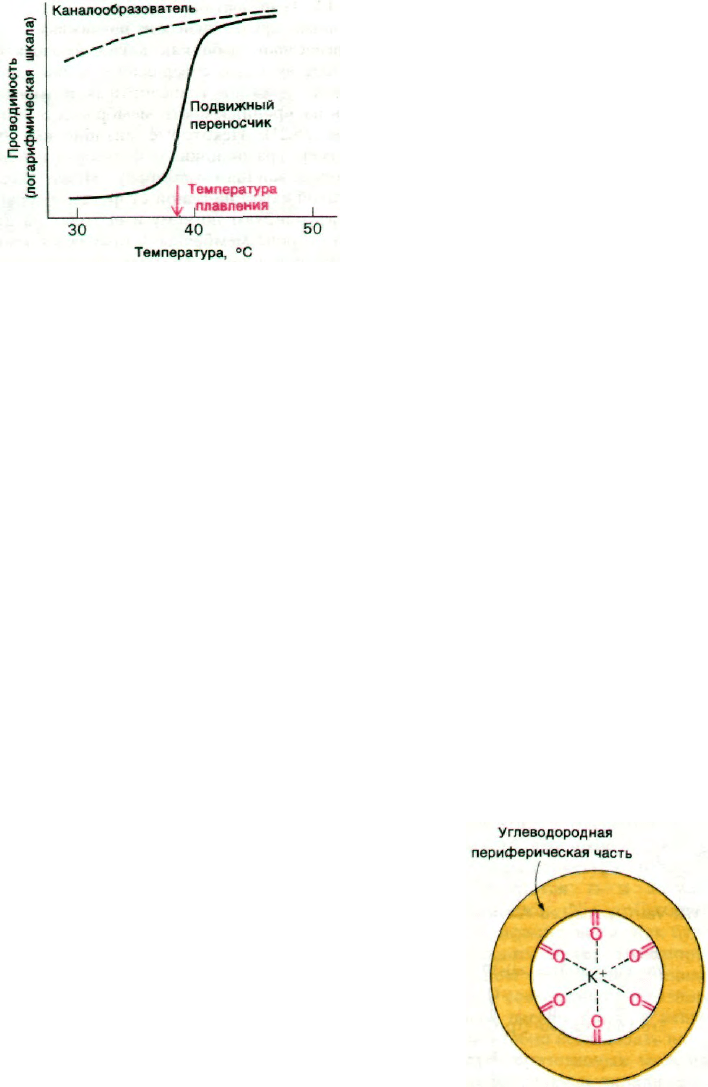
Рис. 36.22. Влияние температуры на про-
водимость липидных дву-
слойных мембран, одна из ко-
торых содержит транспортный
антибиотик-каналообразова-
тель, другая - транспортный
антибиотик, являющийся под-
вижным переносчиком.
браны, содержащей транспортный антибио-
тик. При этом берут температурный интер-
вал, включающий температуру фазового
перехода липида; в этом интервале углево-
дородная середина мембраны переходит от
практически затвердевшего состояния до
совершенно жидкого. Каналообразователь,
опосредуя транспорт ионов через мембрану,
сам по себе не диффундирует. Следователь-
но, застывание углеводородного слоя не
окажет существенного влияния на его спо-
собность транспортировать ионы. Иная си-
туация складывается с подвижным перено-
счиком ионов, который для проявления
активности должен диффундировать сквозь
углеводородный слой мембраны: его эффек-
тивность при затвердении углеводородного
слоя должна значительно уменьшиться.
Данные экспериментов с валиномицином
и грамицидином А действительно выяв-
ляют различие между двумя рассматри-
ваемыми механизмами (рис. 36.22). Прони-
цаемость содержащей валииомицин дву-
слойной мембраны возрастает при ее
разжижении более чем в 1000 раз. В то же
время на транспортную активность грами-
цидина А переход мембраны в жидкое со-
стояние почти не оказывает влияния.
Важно подчеркнуть, что все известные
в настоящее время природные системы
транспорта представляют собой каналы.
318
Часть V.
Молекулярная физиология
Как уже обсуждалось выше (разд. 10.14),
перемещение интегральных мембранных
белков с одной поверхности мембраны на
другую (поперечная диффузия) идет крайне
медленно либо вовсе отсутствует. Следова-
тельно, они не могут служить подвижными
переносчиками.
36.15. Антибиотики-переносчики имеют
форму скорлупы ореха и связывают ионы
в своей центральной полости
По данным рентгеноструктурного анализа
и спектроскопических исследований, анти-
биотики - подвижные переносчики ионов -
характеризуются определенной общностью
структуры. Все исследованные к настояще-
му времени переносчики по форме напо-
минают скорлупу ореха. Единственный
связываемый ион металла образует коор-
динационные связи с несколькими атомами
кислорода, окружающими центральную по-
лость. Число таких атомов кислорода
(связывающих ион металла) обычно равно
6 или 8. Периферическая часть структуры
подвижного переносчика состоит из углево-
дородных групп (рис. 36.23).
Функция центрально расположенных ато-
мов кислорода, окруженных углеводо-
родным наружным слоем, вполне очевидна.
В самом деле, в водной среде ион металла,
например К
+
, связывает через кислород не-
сколько молекул воды. Антибиотик-перено-
счик конкурирует с молекулой воды за
связывание иона, образуя с ним хелатное со-
единение через несколько соответствующим
образом расположенных атомов кислорода
в центральной полости. Благодаря углево-
дородной периферической части переносчи-
Рис. 36.23. Схематическое изображение
хелатирования К
+
атомами
кислорода в транспортном
антибиотике - подвижном пе-
реносчике.

Рис. 36.24. Модели валиномицина (А)
и его комплекса с К
+
(Б). При
связывании К
+
происходит из-
менение конформации анти-
биотика. [Изображено в со-
ответствии с атомными
координатами, любезно пред-
оставленными д-ром W. Duax
(для валиномицина) и д-ром
L. Steinrauf (для комплекса
валиномицин—К
+
).]
ка его комплекс с ионом растворим в липи-
дах мембран. По существу, каталитическое
действие рассматриваемых антибиотиков
на транспорт ионов через мембраны и со-
стоит в том, что они переводят ионы в рас-
творимую в липидах форму.
На рис. 36.24 показаны структуры вали-
номицина и его комплекса с К
+
. Ион К
+
координирован с шестью атомами кислоро-
да, описывающими октаэдр вокруг цен-
тральной полости молекулы. Эти атомы
кислорода принадлежат карбонильным
группам шести остатков валина в антибио-
тике. Метильные и изопропильные боковые
цепи формируют углеводородную наруж-
ную часть молекулы валиномицина.
Интересна также структура комплекса с
К
+
другого переносящего ионы антибиоти-
ка, а именно нонактина (рис. 36.25). В этом
комплексе К
+
связан с восемью атомами
кислорода в центре молекулы. Четыре ато-
ма кислорода из восьми принадлежат кар-
боксильным группам, а остальные четы-
ре - эфирным связям.
36.16. Валиномицин связывает К
+
в 1000 раз
прочнее, чем Na
+
Каким образом соединения, транспорти-
рующие ионы, различают такие сходные
ионы, как Na
+
и К
+
? Валиномицин связы-
вает К
+
в 1000 раз прочнее, чем Na
+
, т.е. из-
бирательность этого переносчика выше, чем
у (Na
+
+ К
+
)-АТРазы. Рассмотрим основу
столь высокой избирательности. Как пока-
зали спектроскопические исследования,
комплексы валиномицина с Na
+
и К
+
очень
сходны по структуре. В частности, в обоих
комплексах одинаковы длины координа-
ционных связей между катионом в центре
молекулы и атомами кислорода. Почему же
тогда К
+
связывается намного прочнее, чем
Na
+
? Причина этой избирательности в том,
что К
+
слабее, чем Na
+
, притягивает воду.
Свободная энергия образования гидратной
оболочки для Na
+
на 17 ккал/моль ниже,
чем для К
+
(табл. 36.2). Следовательно, ва-
линомицин связывает К
+
прочнее потому,
что отделение воды от К
+
требует менъ-
Таблица 36.2. Свойства щелочных катионов
Ион
Li
+
Na
+
К
+
Rb
+
Cs
+
Порядковый
номер
элемента
3
11
19
37
55
Радиус иона,
А
0,60
0,95
1,33
1,48
1,69
Свободная
энергия
гидрата-
ции,
ккал/моль
-98
-72
-55
-51
-47
36. Мембранный транспорт
319
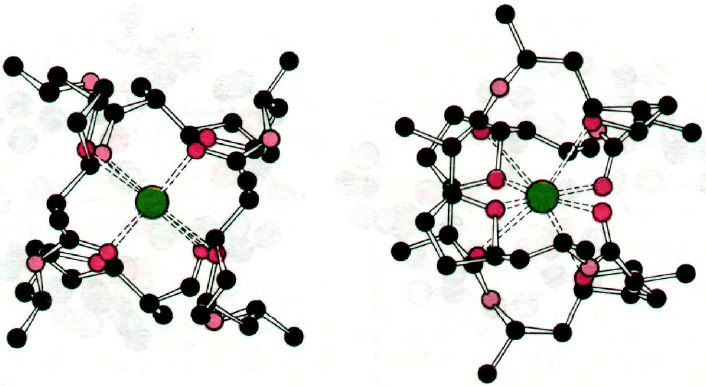
Рис. 36.25. Вид с двух сторон модели ком-
плекса нонактина с К
+
.
шей затраты энергии, чем отделение Na
+
.
Пока еще не проводилось исследования
атомарной структуры какого-либо ион-
транспортирующего соединения, предпоч-
тительно связывающего Na
+
, однако мож-
но представить себе, каким должна быть
ключевая особенность такой структуры. По-
видимому, в ней должно отражаться ис-
пользование того факта, что Na
+
имеет
меньший ионный радиус (r = 0,95 А), чем
К
+
(r = 1,33 А), благодаря чему отрицатель-
но заряженные атомы кислорода могут
в большей степени приблизиться к Na
+
, чем
к К
+
; таким путем может быть преодолен
более высокий энергетический барьер деги-
дратирования Na
+
но сравнению с К
+
. Сле-
довательно, можно предсказать, что участ-
ки специфического связывания Na
+
должны
обладать высокой плотностью отрицатель-
ного заряда и такой геометрией, которая
позволяла бы Na
+
плотно входить в них. В
отличие от этого участки специфического
связывания К
+
должны, по-видимому, обла-
дать отрицательным зарядом меньшей
плотности и их геометрия должна меньше
подходить для образования очень коротких
связей между катионом и координирующи-
ми группами.
Примечательна также гибкость молекулы
валиномицина (рис. 36.24). Хелатирование
320
Часть V.
Молекулярная физиология
К
+
представляет собой ступенчатый про-
цесс, в ходе которого молекулы воды ги-
дратной оболочки иона последовательно
вытесняются кислородными атомами анти-
биотика. Новые связи формируются по мере
разрыва старых, и потому активационный
барьер для связывания иона оказывается
низким. Аналогичным образом и энергия
активации противоположного процес-
са - высвобождения иона - также низка.
В итоге валиномицин присоединяет и выс-
вобождает К
+
множество раз на протяже-
нии секунды. Важная роль гибкости струк-
туры в этом случае так же очевидна, как
и при действии ферментов.
36.17. Можно выявить поток ионов
через единичный канал в мембране
Проводимость планарной двуслойной мем-
браны, содержащей небольшое количество
грамицидина А, не является постоянной.
Как показал Денис Хейдон (Denis Haydon),
проводимость этой мембраны для Na
+
из-
меняется во времени ступенчато (рис. 36.26).
Эти ступени проведения иона определяются
спонтанным открыванием и закрыванием
образованных грамицидином А каналов.
Единичный канал остается открытым в те-
чение примерно секунды. По такому каналу
легко проходят одновалентные катионы, но
не анионы или двухвалентные катионы. Ока-
залось, что по одному каналу в течение се-
кунды мажет, пройти более 10
7
ионов. Такая
скорость транспорта только на один поря-
док величины ниже скорости диффузии в чи-
стой воде. В отличие от этого максимальная
скорость транспорта, осуществляемого под-
вижным переносчиком, составляет менее