Сиволоб А.В. Молекулярна біологія. Підручник
Подождите немного. Документ загружается.

Розділ 8. Синтез білків
235
AP
E
мРН
К
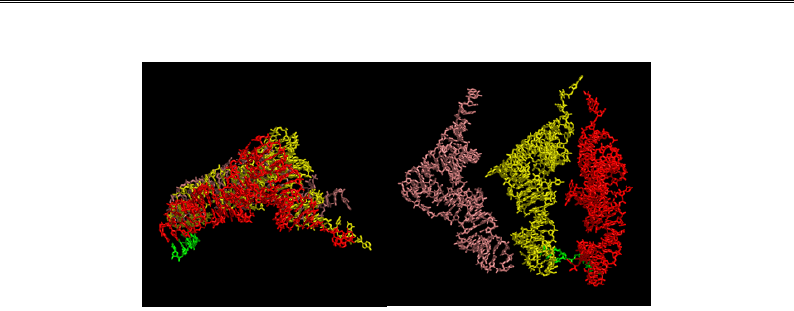
Рис. 8.11. Взаємне розташування трьох молекул тРНК у рибосомі
(дві проекції приблизно збігаються з проекціями на рис. 8.9 і 8.10)
Аа-тРНК потрапляє до рибосоми через щілину між субодиницями
(рис. 8.9), розмір якої може змінюватись унаслідок рухливості струк-
турних елементів рибосоми. Так само й розмір “воріт”, через які тРНК
виходить із Е-сайта, модулюється рухливістю виросту L1.
Функціональну схему рибосоми наведено на рис. 8.12: систему зо-
бражено в момент перед додаванням четвертої амінокислоти до лан-
цюга, що синтезується; показано аа-тРНК, пептидил-тРНК і деаміноа-
цильовану тРНК, що зв'язані відповідно з А-, Р- і Е-сайтами. Централь-
ні сайти, навколо яких відбувається вся робота рибосоми – сайти
зв'язування тРНК – формуються обома субодиницями. Між субодини-
цями існує своєрідне відносне розділення праці: маленька субодини-
ця, яка взаємодіє з мРНК та антикодоновими частинами тРНК, відпо-
відає головним чином за декодуючу функцію рибосоми, а велика, яка
взаємодіє з акцепторними частинами тРНК – за каталітичну функцію.
Рибосомні РНК становлять ~2/3 маси рибосоми й саме вони ви-
значають її структуру та функції. Полінуклеотидний ланцюг рРНК
утворює велику кількість подвійних спіралей, з'єднаних петлями, –
переважно це шпильки, проте відбувається і спарювання між відда-
леними по ланцюгу ділянками. Загалом ланцюг утворює складну про-
сторову структуру, де подвійні спіралі взаємодіють одна з одною та
з одноланцюговими ділянками. Зокрема, аденозини одноланцюгових
ділянок досить часто взаємодіють з маленьким жолобком подвійних
спіралей, стабілізуючи структуру.
У складі 16S рРНК близько половини нуклеотидів залучено до
~60 коротких дволанцюгових ділянок середньою довжиною вісім пар
основ. Але зазвичай у складі 16S РНК виділяють 45 більш-менш
Сиволоб А.В. Молекулярна біологія
236
суцільних (із короткими неспареними ділянками всередині) подвійних
спіралей. За своєю загальною Y-подібною формою 16S рРНК майже
не відрізняється від маленької субодиниці рибосоми (рис. 8.13, по-
рівн. рис. 8.8) – просторова структура чітко розділяється на чотири
структурні домени: 5'-кінцевий формує тіло маленької субодиниці,
центральний – платформу, так званий 3'-кінцевий мажорний домен –
головку, 3'-кінцевий мінорний, який складається з довгої спіралі 44
(порядковий номер) і коротшої останньої спіралі 45, формує інтер-
фейс взаємодії з великою субодиницею. На цей РНК-скелет у складі
маленької субодиниці “нарощуються” білки, однак загальна форма
зберігається. При цьому сайти зв'язування мРНК і антикодонових ча-
стин тРНК формуються переважно 3'-кінцевою зоною 16S рРНК
майже без участі білків ; взаємодія між субодиницями також пере-
важно забезпечується контактами РНК-РНК.
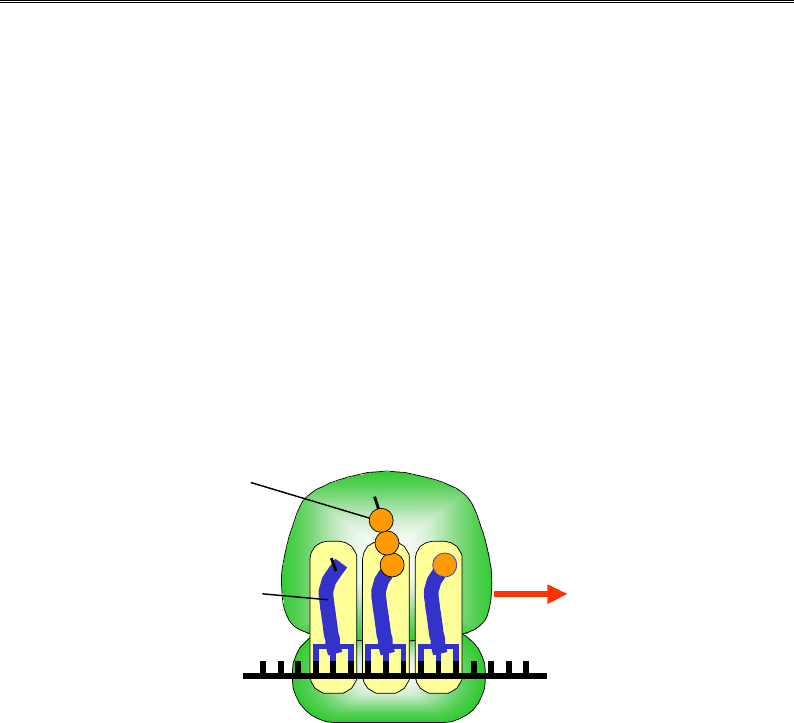
Амінокислоти
т
РН
К
мРНК
Велика
субодиниця
Маленька
субодиниця
5'
A
43
2
1
H
2
N
OH
PE
3'
Рис. 8.12. Функціональна схема рибосоми: червоною стрілкою
позначено напрямок руху при трансляції, амінокислоти пронумеровано
відповідно до порядку включення їх до ланцюга
Еукаріотична 18S рРНК є гомологічною 16S, відрізняючись кілько-
ма вставками.
У складі 23S рРНК розрізняють шість структурних доменів, але во-
ни тісно взаємодіють один з одним. Порівняно із 16S рРНК, ця струк-
тура є значно монолітнішою (рис. 8.13). Така монолітність зумовлена
головною каталітичною функцією великої субодиниці – каталіз не пе-
редбачає конформаційної рухливості, а навпаки, вимагає жорсткості
просторової організації. Каталітична активність великої субодиниці
Розділ 8. Синтез білків
237
пов'язана саме з певною зоною у складі 23S рРНК, яка також взаємо-
діє з акцепторними частинами тРНК. Порівняно невелика 5S рРНК
у комплексі з певними рибосомними білками взаємодіє з 23S, фор-
муючи центральний протуберанець.
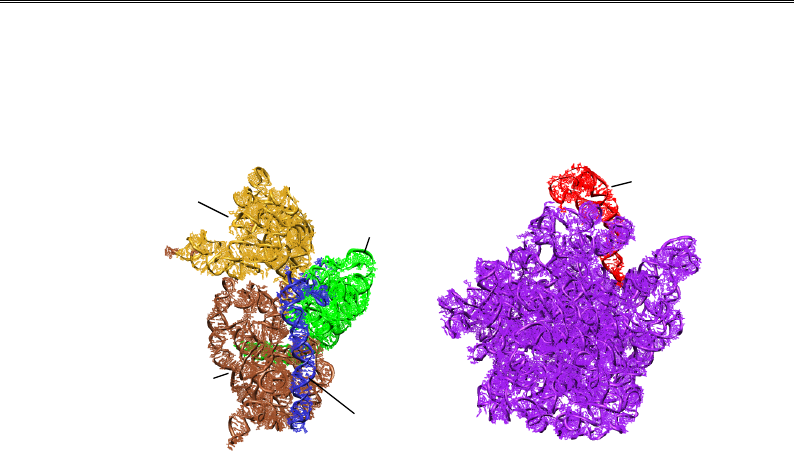
5'-кінцевий
3'-кінцевий
мажорний
3'-кінцевий
мінорний
центральни
й
5S
16S 23S
Рис. 8.13. Рибосомні РНК у складі маленької (2B9O) і великої (2B9P)
субодиниць рибосоми Thermus thermophilus з боку інтерфейсу взаємодії
(як на рис. 8.8). Указано структурні домени рРНК 16S, довга спіраль
у складі 3'-кінцевого мінорного домену – спіраль 44
Еукаріотична 5,8S рРНК є гомологічною 5'-кінцевій зоні 23S. Уза-
галі еукаріотичні 28S і 5,8S рРНК, які утворюють міцний комплекс
між собою, – це ніби трохи подовжена за рахунок вставок 23S РНК,
розділена на дві нерівні частини.
Рибосомні білки розміщуються на поверхні рРНК (відповідно, і на
поверхні рибосоми). У складі маленької субодиниці платформа, щілина
між платформою та головкою, а також інтерфейс взаємодії з великою
субодиницею майже не містять білків, котрі розміщені головним чином
на зовнішньому боці субодиниці (рис. 8.8). По поверхні великої субоди-
ниці білки розподілені рівномірніше, хоча збідненим на них є інтер-
фейс взаємодії з маленькою субодиницею, і навпаки, палець L7/12
утворений кількома копіями відповідних білків без участі РНК.
Більшість рибосомних білків мають дві частини у своїй структурі:
глобулу, розміщену на поверхні рибосоми, і позитивно заряджений
хвіст (нерегулярна ділянка, петля або одна–дві окремі
α-спіралі), який
занурюється в сітку рРНК і стабілізує структуру останньої (рис. 8.14).
У деяких білків глобула відсутня. Отже, усі рибосомні білки (за винят-
ком L7/12) взаємодіють з рРНК, хоча не всі мають високу спорідне-

Сиволоб А.В. Молекулярна біологія
238
ність до вільної рРНК. Збирання субодиниць рибосоми з рРНК та біл-
ків (може бути здійснено in vitro без участі будь-яких факторів) відбу-
вається поступово, через чотири–пять стадій: зв'язування частини біл-
ків на кожній стадії індукує конформаційну зміну рРНК у складі ком-
плексу, що викликає спорідненість до нової порції білків.
L4
L15
S9
S13
Рис. 8.14. Приклади рибосомних білків
у структурі рибосоми (2B9O, 2B9P).
Унизу: взаємодія білка L15 з рРНК 23S
Головна роль рибосомних білків – підтримувати функціонально ак-
тивну структуру рРНК. Хоча рРНК відповідає за майже всі активно-
сті рибосоми, активної конформації вона набуває лише в комплексі
з рибосомними білками. Крім того, деякі білки беруть участь у взає-
модіях з елементами системи трансляції. Наприклад, білок S1, що
міститься поблизу від сайта зв'язування мРНК, сприяє розплітанню
дволанцюгових шпильок у складі матриці; комплекс S1–S18–S21
взаємодіє з мРНК, а також ініціаторною тРНК при ініціації трансля-
ції; білок L10 сумісно з певною ділянкою 23S рРНК і білком L11 ор-
ганізує сайт зв'язування G-білків у основі стебла L7/12.

Розділ 8. Синтез білків
239
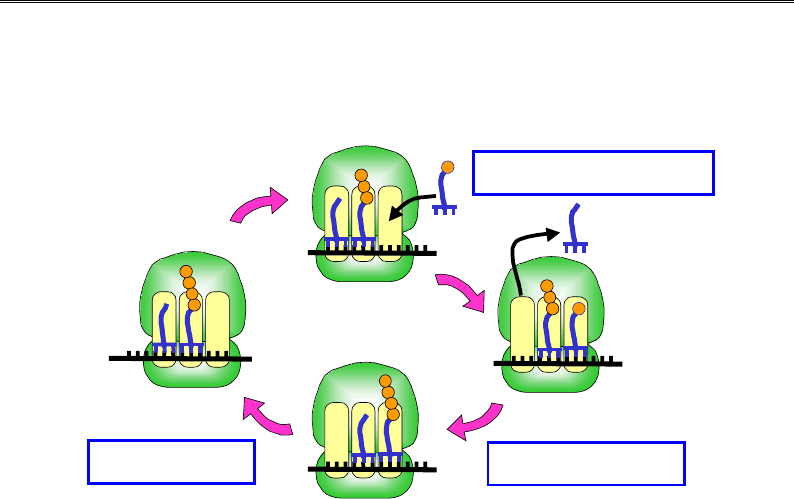
Елонгаційний цикл
Робота рибосоми під час елонгації трансляції полягає в послідовно-
му (потриплетно) зчитуванні інформації з мРНК і відповідному при-
єднанні амінокислот до поліпептидного ланцюга. Кожен такий крок
складається з трьох операцій, що циклічно повторюються (елонгацій-
ний цикл). Цикл розпочинається з такої конфігурації системи, коли
в Р-сайті знаходиться пептидил-тРНК, А-сайт є вільним від тРНК
і в його межах на маленькій субодиниці розташований черговий ко-
дон, який має бути впізнаним (рис. 8.15).
Перша операція циклу –
зв'язування аа-тРНК з А-сайтом. Зв'язування має відбутися з високою
специфічністю щодо взаємодій між кодоном і антикодоном – тільки
споріднена до даного кодона тРНК має бути відібрана системою. Про-
цес розміщення аа-тРНК в А-сайті часто супроводжується дисоціацією
з Е-сайта деаміноацильованої тРНК, яка залишилася там з попередньо-
го циклу. Наслідком зв'язування є належне розташування акцепторних
частин аа-тРНК і пептидил-тРНК відносно одна одної та каталітичного
активного центру. У результаті рибосома здійснює
другу операцію –
транспептидацію
– перенесення пептидилу з пептидил-тРНК на амі-
нокислоту у складі аа-тРНК. Наслідком є перебудова системи: в А-сайті
опиняється пептидил-тРНК із подовженим на одну амінокислоту пеп-
тидилом, у Р-сайті – деаміноацильована тРНК.
Третя операція –
транслокація
– полягає в переміщенні рибосоми на один кодон уз-
довж мРНК (молекули тРНК залишаються зв'язаними зі своїми кодона-
ми), після чого розпочинається наступний елонгаційний цикл.
Відбір серед різних молекул аа-тРНК на першому етапі, коли не-
споріднені молекули мають швидко звільнятися, а також рух рибосо-
ми при транслокації, передбачають реалізацію певної “відкритої”, не
жорстко зафіксованої, рухливої структурної форми рибосоми. Каталіз
транспептидації, навпаки, вимагає жорсткої фіксації субстратів
у “закритій”, жорсткій і нерухливій формі рибосоми. Ефективне ви-
рішення цих суперечливих завдань залежить від полегшення реаліза-
ції відкритих форм рибосоми на першому та третьому етапах елонга-
ційного циклу завдяки двом факторам елонгації (EF – Elongation
Factors): EF1 (еукаріотичний аналог позначається як eEF1 або eEF1А,
eEF – eukariotic Elongation Factor) і EF2 (еукаріотичний аналог – eEF2).
Обидва фактори належать до родини G-білків, або GTP-зв'язу-
Сиволоб А.В. Молекулярна біологія
240
вальних білків. Інші білки цієї родини залучені до багатьох різномані-
тних процесів, зокрема, є елементами асоційованих із мембранами
клітинних сигнальних систем.
5'
A
4
PE
3'
2
1
3
5'
A
4
PE
3'
2
1
3
5'
A
PE
3'
4
2
1
3
5'
A
PE
3'
4
2
1
3
Зв'язування аа-тРНК
Транспептидація
Транслокація
Рис. 8.15. Схема елонгаційного циклу
Елонгаційний фактор EF1
Фактор EF1 (часто також позначається як EF-Tu) – мономерний бі-
лок, що має три структурні домени, – може існувати у двох структур-
них станах (рис. 8.16). Співвідношення між вільними енергіями цих
станів, а відповідно й імовірність переважної реалізації одного з них,
залежить від типу ліганду (GTP чи GDP), що зв'язаний з білком (у по-
вній відповідності до схеми на рис. 2.21). У комплексі з GDP (гуанозин-
дифосфат) реалізується відкрита конформація EF1 з порушеними вза-
ємодіями між структурними доменами. Заміна GDP на GTP (гуанозин-
трифосфат) приводить до локальної конформаційної перебудови
в
межах GTP-зв'язувального домену, унаслідок якої певні амінокислотні
групи виводяться до інтерфейсу взаємодії з іншими доменами – така
взаємодія відбувається і структура “замикається”. Гідроліз GTP (який
здійснюється за певних умов самим білком, див. нижче) проводить до
заміни ліганду і, відповідно, до зворотного перемикання конформації.
Структурна перебудова EF1 є важливою не сама по собі. GDP-
і GTP-асоційовані структурні форми мають відповідно низьку та
високу спорідненість до аа-тРНК і рибосом: саме у вигляді потрійного

Розділ 8. Синтез білків
241
комплексу EF1·GTP–аа-тРНК (рис. 8.17) і відбувається зв'язування
аа-тРНК з рибосомою на першому етапі елонгаційного циклу. При
цьому вже потрійний комплекс відіграє роль ліганду, що перемикає
структурні стани рибосоми.
GDP
GTP
Рис. 8.16. Структура фактора EF1 у комплексі
з GDP (1TUI) і GTP (1TFT). Зелена кулька – іон Mg
2+
Рис. 8.17. Комплекс EF1·GTP з аа-тРНК (1В23)
Зв'язування EF1·GTP із акцепторною частиною аа-тРНК (рис. 8.17)
відбувається відразу після аміноацилювання тРНК. При завершенні
процесу взаємодії з рибосомою (див. нижче) EF1 здійснює гідроліз

Сиволоб А.В. Молекулярна біологія
242
GTP, що веде до втрати спорідненості та дисоціації EF1·GDP. Після
дисоціації молекула GDP витісняється білковим кофактором EF-Ts
(еукаріотичний аналог – eEF1В), який, у свою чергу, замінюється на
молекулу GTP, і знову відбувається зв'язування EF1·GТP з
новою
молекулою аа-тРНК.
Зв'язування аа-тРНК з А-сайтом рибосоми
Роль фактора EF1 і гідролізу GTP. Щоб аа-тРНК могла потрапи-
ти до рибосоми, рибосома має опинитися в певному “відкритому”
структурному стані з розширеним каналом між субодиницями з боку
стебла L7/12 (див. рис. 8.9). Крім того, така відкрита форма рибосо-
ми не допускає дуже міцної взаємодії з аа-тРНК, тобто сприяє її лег-
кій дисоціації в разі невідповідності між кодоном і антикодоном.
Однак зрозуміло, що відкрита форма рибосоми із частково поруше-
ними контактами (енергетично вигідними взаємодіями) між субоди-
ницями має характеризуватися підвищеною вільною енергією, тобто
є малоймовірною. Інший важливий аспект полягає в тому, що на етапі
первинного перебору аа-тРНК з різними антикодонами необхідно
повністю виключити для акцепторної частини тРНК можливість ви-
падково потрапити до пептидилтрансферазного центру. Узагалі, для
такого великого ліганду, як тРНК, існує кінетичний бар'єр асоціа-
ції / дисоціації: велика кількість контактів з лігандом вимагає утво-
рювати / руйнувати їх одночасно – проміжні стани з лімітованим на-
бором контактів мають забезпечити прискорення обох процесів. Роль
EF1 саме й полягає у вирішенні зазначених проблем:
• Взаємодія EF1 з акцепторною частиною аа-тРНК залишає
тільки антикодонову частину вільною для взаємодій з рибо-
сомою (і зв'язаною з нею мРНК).
• При зв'язуванні потрійного комплексу, EF1 (який при цьому
зв'язаний з акцепторною частиною аа-тРНК) взаємодіє із сай-
том у основі стебла L7/12, що виключає контакт акцепторно-
го стебла тРНК з пептидилтрансферазним центром.
• EF1·GTP має підвищену спорідненість до відкритої форми ри-
босоми, тобто подібно до того, як GTP перемикає конформа-
цію самого EF1, потрійний комплекс EF1·GTP–аа-тРНК фіксує
відкриту форму рибосоми, яка стає енергетично вигідною за
рахунок енергії взаємодій з EF1 (рис. 8.18).

Розділ 8. Синтез білків
243
Отже, за умови присутності EF1·GTP здійснюється швидка й не
дуже міцна взаємодія аа-тРНК з рибосомою (головним чином за-
вдяки взаємодії EF1 з основою стебла L7/12), яка також швидко
змінюється дисоціацією: випробування, під час яких антикодонова
частина тРНК намагається впізнати кодон. Якщо таке впізнання
відбувається (див. нижче), спрацьовує GTPазна активність EF1 –
після гідролізу GTP фактор дисоціює, що дозволяє акцепторній час-
тині аа-тРНК остаточно розміститися в А-сайті.
Зв'язування аа-тРНК є внутрішньою властивістю рибосоми:
in vitro зв'язування може відбуватися без участі будь-яких факто-
рів, оскільки супроводжується зниженням вільної енергії. Проте по-
зафакторне зв'язування є досить повільним через наявність на його
шляху енергетичного бар'єра – високоенергетичного відкритого
стану рибосоми (рис. 8.18). Таким чином, EF1 не зумовлює зв'язу-
вання аа-тРНК, а тільки суттєво прискорює цей процес. Можна ска-
зати, що EF1 здійснює каталіз конформаційних перетворень рибо-
соми за рахунок спорідненості до проміжного інтермедіатного стану .
Щодо гідролізу GTP, то він потрібен лише для заміни ліганду з ме-
тою позбавитися цієї спорідненості й забезпечити дисоціацію EF1
і повернення рибосоми до закритого стану, необхідного для насту-
пної стадії елонгаційного циклу. Але це має відбутися лише за умо-
ви впізнання кодона антикодоном аа-тРНК.
Упізнання кодона. Коли під час первинного зв'язування після кі-
лькох випробувань у межах А-сайта на маленькій субодиниці опиня-
ється нарешті комплементарний антикодон, між ним і кодоном має
утворитися коротка подвійна спіраль. Оскільки вона коротка (три
пари основ), то досить нестабільна: кодон-антикодонова взаємодія
за межами рибосоми є неефективною. При цьому рибосома (переду-
сім маленька субодиниця) не тільки створює особливе середовище,
яке стабілізує кодон-антикодонову спіраль, але й сприяє підвищен-
ню специфічності – ефективно дискримінує споріднені та неспорід-
нені до даного кодону антикодони.
При утворенні комплементарної подвійної спіралі, з її маленьким
жолобком взаємодіють два консервативні аденозини рРНК 16S (з по-
рядковими номерами 1492 і 1493 – розташовані у верхній частині
спіралі 44, див рис. 8.13), утворюючи водневі зв'язки з 2'-ОН-групами
рибоз нуклеотидів кодона й антикодона (рис. 8.19). Така взаємодія,
що реалізується лише за умови комплементарності, стабілізує перші
дві (відносно позицій кодона) нуклеотидні пари. Друга нуклеотидна
пара додатково стабілізується, також через маленький жолобок,
Сиволоб А.В. Молекулярна біологія
244
взаємодією з консервативним G530 рРНК 16S. Третя пара кодон-
антикодонової спіралі теж взаємодіє з певним цитидином рРНК 16S,
але ця взаємодія менш специфічна й допускає неоднозначність спа-
рювання (див. підрозділ, де йдеться про тРНК).
AP
2
1
3
AP
2
1
3
4
AP
2
1
3
G
T
P
4
A
P
2
1
3 4
GDP
Вільна
енергія
Відкрита
форма
Закрита
форма
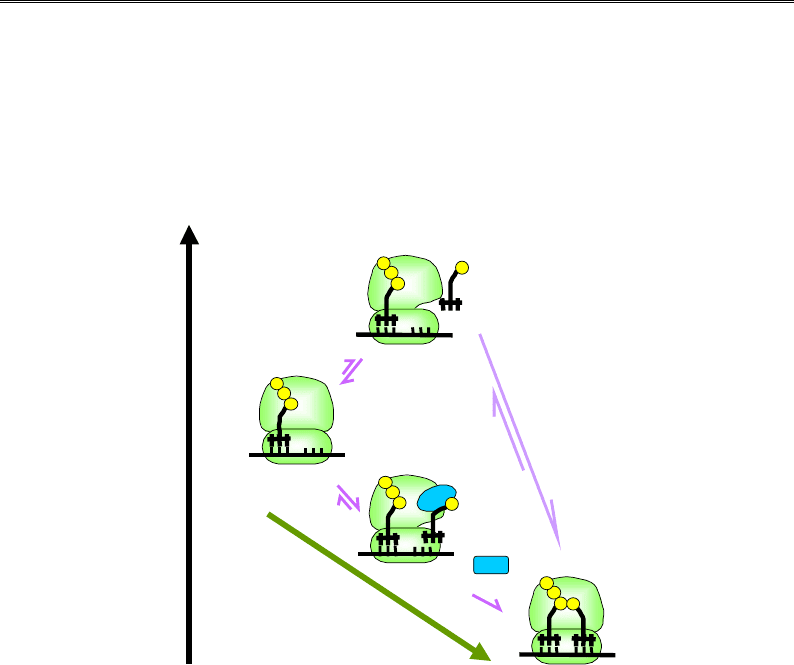
Рис. 8.18. Зв'язування з А-сайтом рибосоми аа-тРНК
у вільному стані та в комплексі з EF1·GTP. Різні стани системи
розміщені на різних рівнях відповідно до їхньої вільної енергії.
Зелена стрілка вказує оптимальний шлях процесу.
Показано тільки два сайти тРНК
Взаємодії нуклеотидів рРНК 16S із кодон-антикодоновою спіраллю
не тільки підвищують специфічність упізнання кодона , але й мають
інший важливий наслідок. Як показано на рис. 8.19, для реалізації
цих взаємодій А1492 і А1493 мають бути переорієнтованими відносно
сусідніх нуклеотидів. Така локальна конформаційна перебудова запу-
скає каскад конформаційних змін в інших частинах рРНК 16S за
“принципом доміно”, що спричиняє глобальну конформаційну пере-
будову всієї маленької субодиниці. Зокрема, головка субодиниці пе-
реміщується в бік великої субодиниці (див. рис. 8.9), міцно замикаючи