Сиволоб А.В. Молекулярна біологія. Підручник
Подождите немного. Документ загружается.


Розділ 4. Організація днк у клітинах: геноми та структура хроматину
123
Інший блок – хромодомен – упізнає метильовані Lys. Гістонові хвос-
ти слугують своєрідними платформами для збирання різноманітних
білкових комплексів, склад яких залежить від патерну модифікацій
– розподілу певних модифікованих груп по хвостах. Співвідношен-
ня між патерном модифікацій і набором білків, які впізнають та-
кий патерн, що у свою чергу має певні функціональні наслідки, на-
зивають
гістоновим кодом.
K9
K14
K18
K23 K27
K5
K8
K12
K16
K5
K15
K20
K12
K5
K9
K4
K9
K27
R2
R17
R26
S10
S1 R3
K20
H3
H4
H2B
H2A
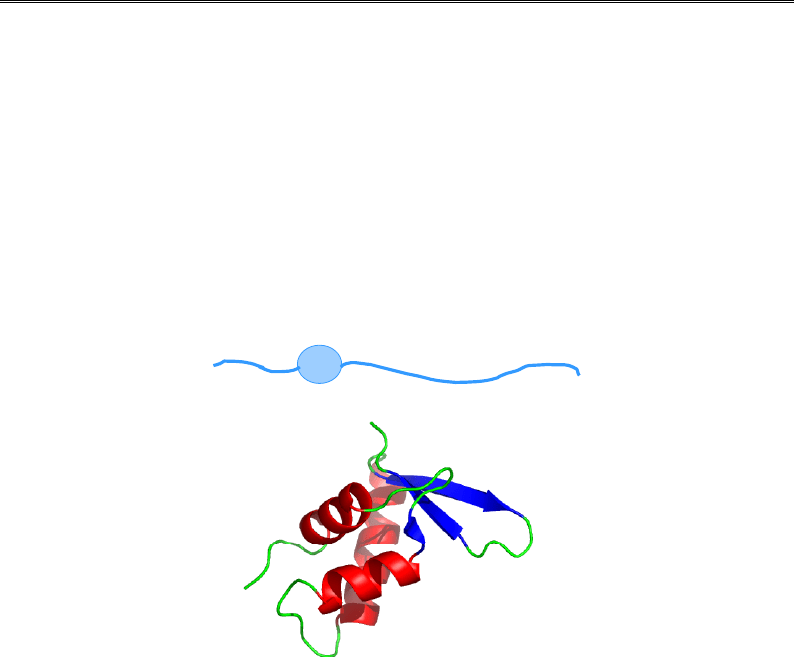
Рис. 4.13. Кінцеві хвости гістонів у структурі нуклеосоми (1KX5)
та їхні модифікації. Кольором виділено по одному N-кінцевому хвосту
кожного гістону, а також С-кінцевий – гістону Н2А (поряд із N-кінцем Н3).
Стрілочками вказано ацетилювання (червоні), метилювання (зелені)
і фосфорилювання (блакитні) залишків певного типу,
які займають певні позиції в поліпептидних ланцюгах
Наприклад, фосфорилювання Н3-Ser10 із одночасним ацетил-
юванням Н3-Lys14 супроводжується активацією транскрипції, деа-
цетилювання Н3-Lys14 із одночасним метилюванням Н3-Lys9 – реп-
ресією. Це лише невеличкий елемент такого коду: зрозуміло, що різ-
номанітних комбінації модифікованих залишків на восьми гістоно-
вих хвостах (рис. 4.13) може бути велика кількість. Остаточне з'ясу-
Сиволоб А.В. Молекулярна біологія
124
вання гістонового коду ще далеко не завершено. Певні його законо-
мірності та інші приклади залучення гістонового коду до регуляції
транскрипції наведені у розділі 6.
Гістон Н1
Одна молекула п'ятого гістону – лінкерного гістону Н1 – взаємодіє з
кожною нуклеосомою у хроматині. Проте ця взаємодія, на відміну від
корових гістонів, є динамічною: спостерігається швидкий обмін лін-
керних гістонів між хроматином та їхнім пулом у ядрі.
N
C
C
N
GH1
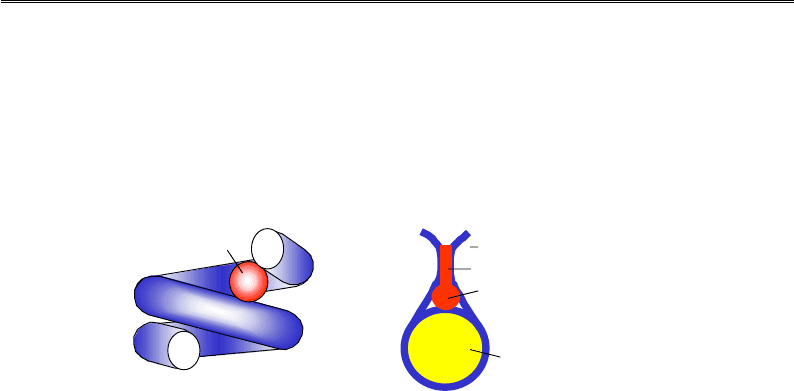
Рис. 4.14. Схема будови гістону Н1
і структура його глобулярного домену GH1
(1HST, аналог гістону Н1 з еритроцитів птахів,
що позначається як Н5)
За своєю структурою гістон Н1 (мономерний білок) суттєво відріз-
няється від корових гістонів (рис. 4.14). Молекула Н1 містить
N-кінцевий невпорядкований хвіст, глобулярний домен GH1 і довгий,
збагачений позитивно зарядженими залишкам (насамперед Lys)
С-кінцевий хвіст довжиною приблизно 100 амінокислотних залишків
(приблизно половина молекули). Глобулярний домен (приблизно
80 амінокислот) належить до так званої родини “спіральних із криль-
цем” (winged helix) ДНК-зв'язувальних білків.
На поверхні домену існують два можливі сайти взаємодії з ДНК,
які формуються кластерами позитивно заряджених амінокислот і ло-
калізовані на протилежних боках глобули. Відповідно, глобулярний
Розділ 4. Організація днк у клітинах: геноми та структура хроматину
125
домен взаємодіє з нуклеосомною ДНК на виході з нуклеосоми, імовір-
но, між двома сусідніми витками (рис. 4.15, а), додатково стабілізую-
чи нуклеосомну суперспіраль. С-кінцевий хвіст Н1 взаємодіє з обома
лінкерами, що виходять з нуклеосоми. У результаті дві лінкерні ділянки
(довжиною по 10–30 пар основ) утворюють
стеблоподібну струк-
туру
на виході з нуклеосоми (рис. 4.15, б).
GH1
GH1
нуклеосома
л
інкер
С-кінцевий хвіст
аб
Рис. 4.15. Схема взаємодії глобулярного домену GH1 (а)
та цілого
гістону Н1 (б) із нуклеосомою
Наднуклеосомна упаковка хроматинової фібрили
У хроматині нуклеосоми з'єднані лінкерами довжиною ~50 пар ос-
нов. Якщо лінкер (у полінуклеосомному ланцюзі без гістону Н1) просто
продовжує хід нуклеосомної ДНК по прямій, нуклеосоми у складі по-
лінуклеосомної нитки мають бути розташовані зиґзаґом (рис. 4.16, а).
Саме такий вигляд має декомпактизована (якщо відсутній Н1 і за
низької іонної сили) полінуклеосомна фібрила під мікроскопом (елект-
ронним чи атомно-силовим). Товщина такого зиґзаґу дорівнює приблиз-
но 30 нм. Полінуклеосомний зиґзаґ може конденсуватися (рис. 4.16, б).
Умова конденсації (необхідна, але недостатня) – фізіологічна іонна
сила (крім одновалентних мають бути присутніми двовалентні катіо-
ни) для зниження електростатичного розштовхування між нуклеосо-
мами.
Фактор конденсації (її рушійна сила) – невпорядковані хвости
корових гістонів: лабільні позитивно заряджені хвости ефективно
“зшивають” фібрилу, взаємодіючи з ДНК сусідніх нуклеосом.
У присутності гістону Н1 компактна хроматинова фібрила товщи-
ною 30 нм стає значно стабільнішою. Ключова роль у цій стабілізації
належить стеблу, що формується унаслідок взаємодії з гістоном Н1
двох лінкерних ділянок на виході з нуклеосоми: за рахунок стебла
сусідні нуклеосоми значно зближуються, що сприяє конденсації
(рис. 4.17). Отже, присутність Н1 наближає нуклеосоми одна до од-
ної у складі фібрили, чим сприяє “зшиванню” цих нуклеосом хвостами
Сиволоб А.В. Молекулярна біологія
126
корових гістонів. І навпаки, компактизація фібрили за участю хвос-
тів корових гістонів сприяє зв'язуванню Н1 із наближеними у прос-
торі ділянками лінкерів.
а б
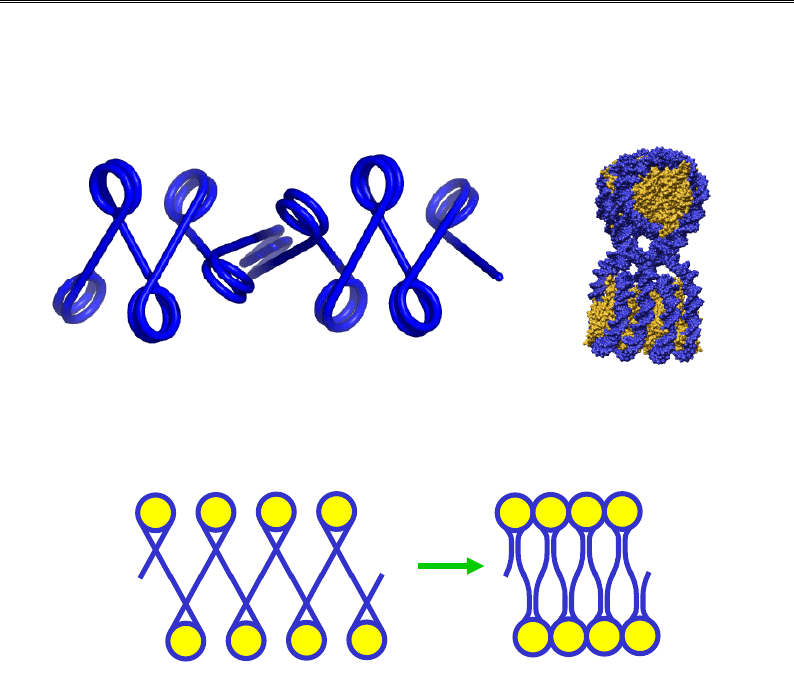
Рис. 4.16. Зиґзаґоподібна конфігурація полінуклеосомної нитки (а)
і структура компактизованого елемента такої нитки
у кристалах тетрануслеосом (б, 1ZBB)
+ Н1
Рис. 4.17. Роль Н1-залежного стебла на виході з нуклеосоми
у стабілізації компактного стану хроматинової фібрили.
Отже, суперструктура конденсованої фібрили товщиною 30 нм яв-
ляє собою тривимірний зиґзаґ нуклеосом, з'єднаних практично пря-
мими – без вигинів – лінкерами, які спрямовані всередину фібрили.
Усередині містяться також і гістони Н1; сумісна дія Н1 і хвостів коро-
вих гістонів підтримує компактний стан фібрили.
Фібрила товщиною 30 нм – основна форма існування інтерфаз-
ного хроматину. Але у хроматині існує значна гетерогенність за
ступенем конденсації. З одного боку, передумовою активації окре-
мих ділянок хроматину є деконденсація фібрили. Факторами такої
деконденсації є зниження спорідненості до ДНК хвостів корових
гістонів унаслідок їхнього ацетилювання (зниження позитивного
заряду) і тимчасова дисоціація Н1. Дисоціації Н1 сприяє ацетилю-

Розділ 4. Організація днк у клітинах: геноми та структура хроматину
127
вання хвостів корових гістонів, а також посттрансляційні модифі-
кації самого Н1, зокрема фосфорилювання. Деконденсація фібрили
та дисоціація Н1 створюють можливість взаємодії з ДНК різномані-
тних негістонових білків, що врешті-решт і викликає активацію
транскрипції (див. розділ 6).
З іншого боку, у репресованих ділянках хроматинова фібрила може
бути як додатково стабілізованою в компактному стані, так і піддава-
тися компактизації вищого порядку. Частина хроматину, що зберігає
стан підвищеної компактизації протягом інтерфази, називається
гетерохроматином (решта хроматину, де в принципі може відбува-
тися активація транскрипції, позначається як еухроматин). Утво-
рення гетерохроматину здійснюватися, головним чином, у ділянках,
що містять повтори – у центромерах, теломерах, зонах концентрації
мобільних елементів. Деталі структури гетерохроматину залишаються
нез'ясованими, механізми встановлення та підтримання неактивного
конденсованого стану гетерохроматину розглядаються в розділі 6.
Петльові домени хроматину та ядерний матрикс
На наступному рівні структурної організації у клітинному ядрі
хроматинова фібрила формує петлі, кінці яких є жорстко закріплени-
ми на скелетних структурах клітинного ядра – ядерному матриксі
(рис. 4.18). Оскільки основи петлі фіксуються на матриксі, ДНК
у складі петлі має топологічні обмеження, тобто є еквівалентною цирку-
лярній (див. розділ 3). Одна петля, що містить від 20 до 200 тис. пар
основ ДНК (один або кілька генів), часто розглядається як важливий
елемент регуляції процесів транскрипції та реплікації.
Із білками матриксу взаємодіють ділянки ДНК довжиною від 300 до
1 тис. пар основ – ділянки, асоційовані з матриксом (MAR, Matrix
Associated Regions). Вони не мають між собою гомології щодо послідо-
вності, часто є збагаченими на АТ-пари. Вважається, що більшість
функціональних процесів, які відбуваються на ДНК, розпочинається
саме на матриксі, тобто на MAR.
Ядерний матрикс – це система білкових філаментів, яка формує
структурний каркас ядра. На периферії ядра розташована особлива
частина матриксу, асоційована із внутрішньою ядерною мембраною –
ядерна ламіна. Філаменти ламіни протягнуті від однієї ядерної пори до
іншої і позиціонують порові комплекси у площині мембрани. Подвійна
ядерна мембрана, ядерні пори й ламіна утворюють єдину систему –
ядерний конверт. Від ламіни всередину ядра протягнуті філаменти
Сиволоб А.В. Молекулярна біологія
128
внутрішнього ядерного матриксу. З ламіною взаємодіє значна частина
гетерохроматину, зокрема центромери та теломери хромосом. Еухро-
матинова частина хромосоми “звисає” всередину ядра, де хроматинові
петлі закріплюються на внутрішній частині матриксу. У результаті
хромосома займає певну зону в об'ємі ядра – хромосомну територію.
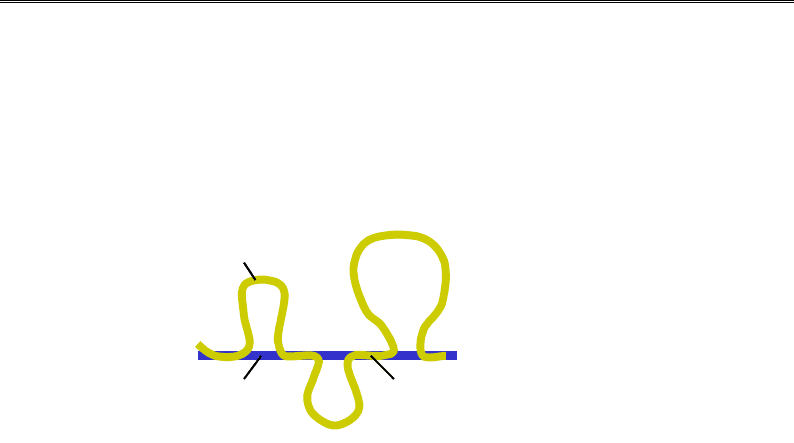
Петля
х
роматинової
фібрили
Філамент
ядерного
матриксу
Д
ілянка,
асоційована з
матриксом (MAR)
Рис. 4.18. Схема петльової організації хроматину
Усе, про що йшлося вище, стосується організації хроматину та хро-
мосом в інтерфазному ядрі. Коли клітина вступає в процес мітозу, піс-
ля реплікації ДНК починається утворення надкомпактної мітотичної
хромосоми, деталі структурної організації якої залишаються недостат-
ньо зрозумілими. Одночасно з компактизацією хроматинової фібрили
частина ламіни “розчиняється” разом із ядерною мембраною , інша ча-
стина, разом із внутрішнім матриксом, перебудовується з утворенням
білкового каркасу мітотичної хромосоми – хромосомного скефолду
(scaffold), з яким залишаються зв'язаними основи хроматинових петель.
КОНТРОЛЬНІ ЗАПИТАННЯ
1. Дайте визначення кодона і відкритої рамки зчитування. Скільки
існує кодонів? Які позиції нуклеотидів у складі кодона є найбільш
і найменш визначальними?
2. Що таке ген?
3. Що таке геном? У чому полягає найсуттєвіша відмінність між ге-
номами про- та еукаріотів?
4. У чому полягає перекриття генів у вірусів та еукаріотів?
5. Дайте визначення інтрона і екзона.

Розділ 4. Організація днк у клітинах: геноми та структура хроматину
129
6. Яка різниця між кластером генів і опероном?
7. Назвіть основні типи послідовностей ДНК, що повторюються.
8. З яких елементів складається нуклеосома? Яке місце в її струк-
турі належить білкам? Як організована у просторі ДНК у нуклеосомі?
9. Які взаємодії стабілізують нуклеосому? За яким принципом від-
бувається збирання нуклеосоми?
10. У чому полягає механізм позиціювання нуклеосом відносно по-
слідовності ДНК?
11. Назвіть основні типи посттрансляційних модифікацій гістонів.
Які амінокислотні залишки піддаються модифікаціям? В яких части-
нах молекул гістонів ці залишки розташовані?
12. Як організована хроматинова фібрила у просторі? За яким основ-
ним механізмом фібрила піддається компактизації? У чому полягає роль
гістону Н1?
13. Що таке ядерний матрикс і яку участь він бере в організації
хроматину в клітинному ядрі?
РЕКОМЕНДОВАНА ЛІТЕРАТУРА
Akey, C.W., Luger K. Histone chaperones and nucleosome assembly
// Curr. Op. Struct. Biol. – 2003. – Vol. 13. – Р. 6–14.
Bednar, J., Horowitz, R.A., Grigoryev, S.A. et al. Nucleosomes, linker
DNA, and linker histone form a unique structural motif that directs the
higher-order folding and compaction of chromatin // Proc. Natl. Acad.
Sci. USA. – 1998. – Vol. 95. – Р. 14173–14178.
Brown T.A. Genomes. – New York ; London: Garland Science, 2002.
Chromatin structure and gene expression: frontiers in molecular biology
/ eds. S. Elgin, J.L. Workman. – Oxford : Oxford University Press, 2002.
Chromatin structure and dynamics: state-of-the-art. New Comprehen-
sive Biochemistry / eds. J. Zlatanova, S.H. Leuba. Amsterdam : Elsevier,
2004. – Vol. 39.
The ENCODE Project Consortium. Identification and analysis of
functional elements in 1 % of the human genome by the ENCODE pilot
project // Nature. – 2007. – Vol. 447. – 799–816.
Gerstein, M.B., Bruce, C., Rozowsky, J.S. et al. What is a gene, post-
ENCODE? History and updated definition // Genome Res. – 2007.
– Vol. 17. – Р. 669–681.
Gruenbaum, Y., Margalit, A., Goldman, R.D. et al. The nuclear lamina
comes of age // Nat. Rev. – 2005. – Vol. 6. – Р. 21–31.

Сиволоб А.В. Молекулярна біологія
130
International Human Genome Sequencing Consortium. Initial sequ-
encing and analysis of the human genome // Nature. – 2001. – Vol. 409.
– Р. 860–921.
International Human Genome Sequencing Consortium. Finishing the
euchromatic sequence of the human genome // Nature. – 2004.
– Vol. 431. –Р. 931–945.
Koonin, E.V. How many genes can make a cell: the minimal-gene-set
concept // Annu. Rev. Genomics Hum. Genet. – 2000. – Vol. 1.– Р. 99–116.
Lesk, A.M. Introduction to bioinformatics. – New York : Oxford
University Press, 2002.
Luger, K., Richmond, T. J. DNA binding within the nucleosome core
// Curr. Op. Struct. Biol. – 1998. – Vol. 8. – Р. 33–40.
Luger, K., Richmond, T.J. The histone tails of the nucleosome
// Curr. Opin. Genet. Dev. – 1998. – Vol. 8 – Р. 140–146.
Pearson, H. Genetics: what is a gene? // Nature. – 2006. – Vol. 441.
– Р. 398–401.
Ramakrishnan, V. Histone H1 and chromatin higher-order structure
// Crit. Rev. Eukaryot. Gene Expr. – 1997. – Vol. 7. – Р. 215–230.
Simpson, R.T. Nucleosome positioning: occurrence, mechanisms, and
functional consequences // Prog. Nucl. Acid Res. Mol. Biol. – 1991.
– Vol. 40. – Р. 143–184.
Sivolob, A., Prunell, A. Nucleosome conformational flexibility and
implications for chromatin dynamics // Phil. Trans. Roy. Soc. Lond.
A.– 2004. – Vol. 362. – Р. 1519–1547.
Strahl, D.D., Allis, C.D. The language of covalent histone modifications
// Nature. – 2000. – Vol. 403. – Р. 41–45.
Tremethick, D.J. Higher-order structures of chromatin: the elusive
30 nm fiber // Cell. – 2007. – Vol. 128. – Р. 651–654.
Wu, H.-L., Bagby, S., van den Elsen J.M.H. Evolution of the genetic
triplet code via two types of doublet codons // J. Mol. Evol. – 2005.
– Vol. 61. – Р. 54–64.

Розділ 5
ТРАНСКРИПЦІЯ: ПРОКАРІОТИ
Down, down, down. There was
nothing else to do.
Lewis Carroll.
Alice's adventures in Wonderland
Транскрипція (transcription) – процес синтезу РНК з використанням
одного з ланцюгів ДНК як матриці, тобто “переписування” послідовно-
сті нуклеотидів ДНК у послідовність нуклеотидів РНК. Зростання лан-
цюга РНК відбувається в напрямку від 5'- до 3'-кінця. Субстратами ре-
акції є 3'-кінцева ОН-група рибози зростаючого транскрипту (ланцю-
га РНК, що синтезується) і рибонуклеозидтрифосфати (rNTP). Фермент,
що каталізує цю реакцію – ДНК-залежна РНК-полімераза (DNA depen-
dent RNA P
olymerase, RNAP). Приступаючи до розгляду прокаріотичної
системи транскрипції, слід зауважити, що молекулярні механізми син-
тезу РНК є в основному спільними для всіх живих організмі в.
РНК-полімераза
Загальний сценарій транскрипції в бактерій
Бактеріальна РНК-полімераза складається з кількох білкових суб-
одиниць та існує, залежно від стадії транскрипції, у двох формах:
1) кор-фермент (core enzyme) у складі субодиниць, що позначаються
як
α
(дві копії), β, β' та ω; 2) голофермент (holoenzyme) – комплекс кор-
ферменту із субодиницею
σ. Кор-фермент має досить високу неспе-
цифічну спорідненість до ДНК, що дозволяє йому працювати на різ-
номанітних послідовностях. Поява субодиниці
σ у складі голофермен-
ту приводить до зниження загальної неспецифічної спорідненості, але
при цьому виникає специфічна спорідненість до особливих ділянок,
з яких транскрипція має розпочинатися – промоторів (promoters).

Сиволоб А.В. Молекулярна біологія
132
За рахунок цього голофермент має змогу ефективно перебирати
(шляхом зв'язування та швидкої дисоціації) різноманітні ділянки
ДНК, здійснюючи пошук промотора, взаємодія з яким є міцнішою.
Отже, субодиниця
σ виконує роль загального фактора ініціації транс-
крипції. Різні промотори розрізняються за спорідненістю до голофер-
менту (силою промотора). Відповідно, упізнання слабких промоторів
залежить від додаткових, специфічних для даного гена чи групи ге-
нів, факторів ініціації (див. нижче).
Робочий цикл РНК-полімерази складається з наступних стадій.
•
Ініціація транскрипції, яка також є багатостадійним проце-
сом (рис. 5.1):
о зв'язування голоферменту з промотором. У результаті фор-
мується закритий комплекс, у складі якого ДНК зберігає
форму подвійної спіралі;
о локальне плавлення подвійної спіралі з утворенням від-
критого комплексу – розходження ланцюгів ДНК, яке до-
зволяє використовувати один з них як матрицю;
о включення перших двох нуклеотидів до молекули РНК (син-
тез першого фосфодіефірного зв'язку в активному центрі
полімерази) – найповільніша стадія процесу;
о зростання первинного короткого транскрипту – приєд-
нання 8–9 нуклеотидів. Після цього є можливою абортивна
ініціація (визволення короткого транскрипту), тобто не-
вдала спроба ініціації;
о в іншому випадку відбувається очищення промотора –
дисоціація
σ-фактора, яка маркує перехід до елонгації
транскрипції.
•
Елонгація транскрипції, у кожному елементарному акті якої
(елонгаційному циклі) відбувається приєднання чергового ну-
клеотиду до 3'-кінця РНК і пересування кор-ферменту на
один нуклеотид уздовж матриці (транслокація).
•
Термінація транскрипції при впізнанні полімеразою спеціа-
льного сигналу термінації (особливого елемента послідовнос-
ті), коли відбувається визволення транскрипту. Далі кор-
фермент знову взаємодіє з
σ-субодиницею і здійснює новий
пошук промотора.
Суттєвою особливістю прокаріотичної системи транскрипції білко-
вих генів є те, що молекула мРНК зв'язується з рибосомами безпосе-
редньо під час транскрипції – транскрипція мРНК і білковий синтез
(розділ 8) є єдиним процесом.