Сидельникова В.М. Привычная потеря беременности
Подождите немного. Документ загружается.


ГЛАВА
1.
Физиология репродуктивной системы
I
I
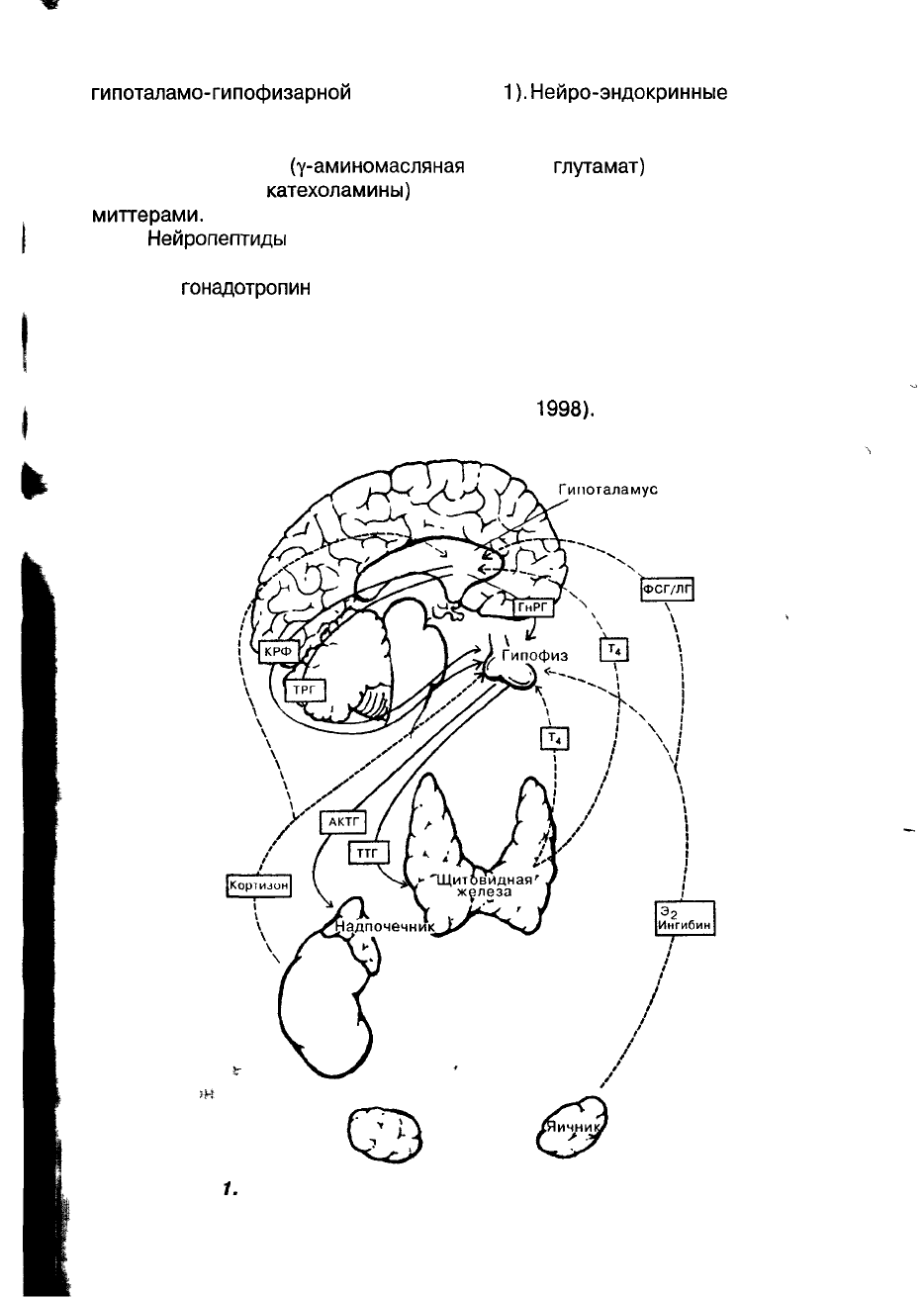
Нейро-эндокринная регуляция репродуктивной системы осуществляется
гипоталамо-гипофизарной
системой (рис.
1).
Нейро-эндокринные
связи осуще-
ствляются химическими медиаторами: моноаминами, аминокислотами и нейро-
пептидами.
Аминокислоты
(у-аминомасляная
кислота,
глутамат)
и моноамины (ацетил-
холин, серотонин,
катехоламины)
считаются быстродействующими нейротранс-
миттерами.
Нейропептиды
действуют медленнее, но более длительно. К нейропептидам
относят группу гормонов, имеющих регулирующее значение для репродуктивной
системы:
гонадотропин
— релизинг-гормон, тиреотропин — релизинг-гормон, кор-
тикотропин — релизинг-гормон. Пептиды мозга были выделены, расшифрова-
ны и синтезированы, что позволяет более точно определить их роль в нейро-эн-
докринном контроле репродуктивных процессов у человека. Не менее важным
было получение специфических антител к нейропептидам, что позволило опре-
делить их локализацию и концентрацию (Yen,
1998).
Рис.
1.
Нейро-эндокринная регуляция репродуктивной системы.
13
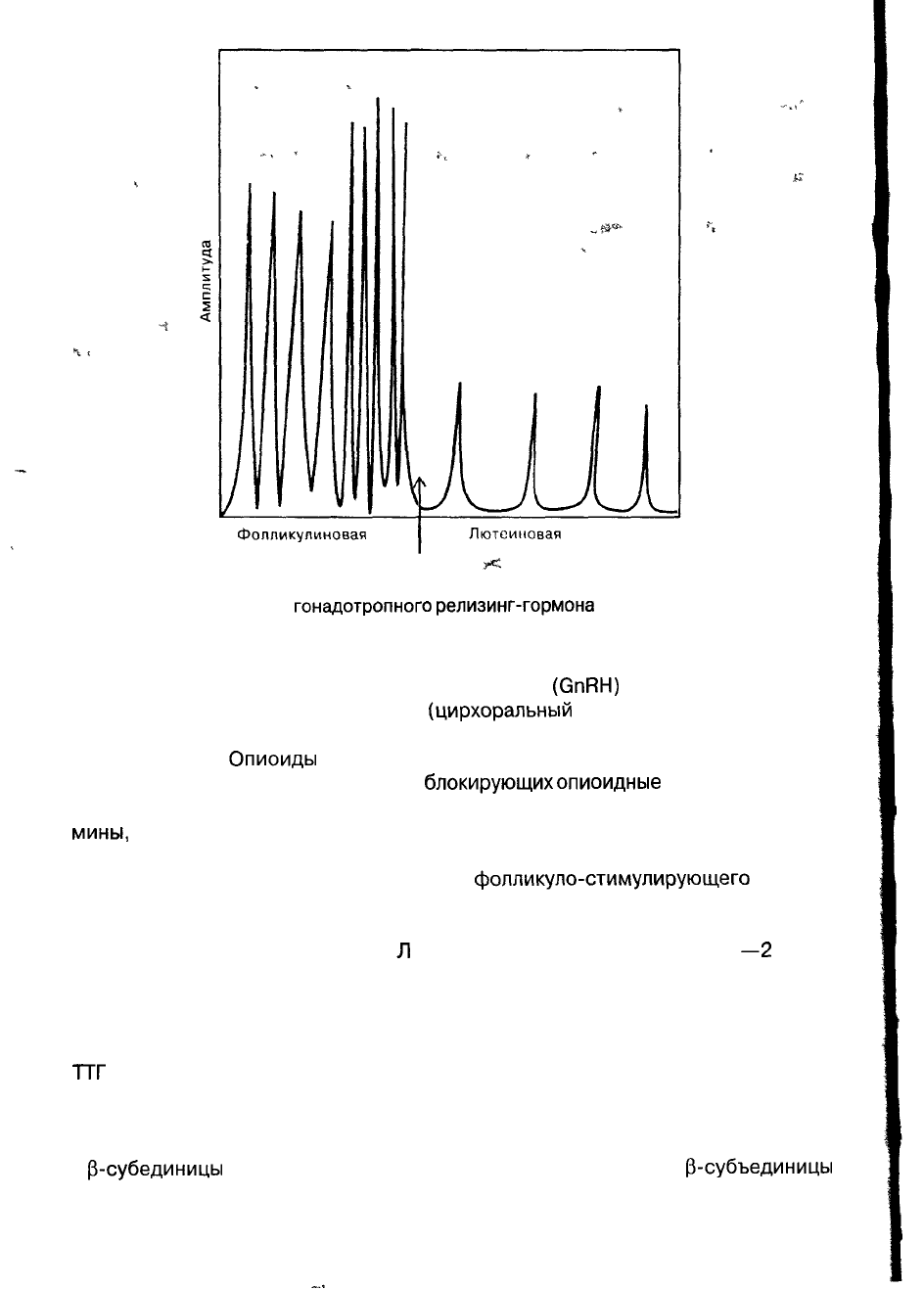
Фолликулиновая
фаза цикла
Лютсимовая
фаза цикла
Овуляция
Рис. 2. Секреция
гонадотропного
релизинг-гормона
в фолликулиновую
и лютеиновую фазы цикла
Секреция гонадотропного релизинг-гормона
(GnRH)
носит импульсный ха-
рактер — примерно 1 импульс в час
(цирхоральный
ритм) (рис. 2). Выделение
GnRH регулируется по механизму обратной связи с эстрадиолом и прогестеро-
ном (Yen, 1982).
Опиоиды
оказывают тормозящее влияние на выделение GnRH.
При введении препаратов (налоксан),
блокирующихопиоидные
рецепторы, резко
увеличивается выброс GnRH. Важную роль в контроле GnRH играют и катехола-
мины,
которые также оказывают тормозящий эффект.
GnRH связывается со специфическими рецепторами на гонадотрофах ги-
пофиза и стимулируют синтез и секрецию
фолликуло-стимулирующего
(ФСГ) и
лютеинизирующего (ЛГ) гормонов. Секреция происходит прерывисто и синхро-
низирована с пульсирующим выделением GnRH из гипоталамуса. Импульсы ФСГ
значительно меньше, чем импульс
Л
Г. Интервалы между импульсами 1
—2
часа в
фолликулиновую фазу цикла и около 4 часов в лютеиновую. В средине цикла уве-
личивается и частота и амплитуда импульсов.
По химической структуре ЛГ и ФСГ сходны, они оба являются гликопроте-
идами, состоят из ос и р — субъединиц: а-субъединица — общая (в том числе для
ТТГ
и ХГ) и р-субъединица варьирует, и этим определяется специфичность дейст-
вия каждого гормона.
Обе субъединицы синтезируются в одних и тех же клетках, но с участием
разных генов. В настоящее время определена структура генов. Установлено, что
у
р-субединицы
ЛГ ген одинаков у человека, крысы и мыши. Для
р-субъединицы
14

ХГЧ существует 7 генов или псевдогенов в
19
хромосоме, собранных в единый
тандем. Особенностью этого гена является возможность поддерживать его экс-
прессию в условиях повышенной эстрогенизации, чтобы обеспечить постоянст-
во продукции ХГ для поддержания желтого тела (Talmadge и
соавт.,
1984).
Биологическое действие ФСГ направлено на фолликулогенез. Рецепторы
f
ФСГ имеются в гранулезных клетках фолликулов. Посредством этих рецепторов
£
ФСГ стимулирует рост фолликулов и их созревание. В противоположность это-
I му рецепторы к Л Г имеются в
тека-клетках,
интерстициальных
и
лютеиновых
клет-
t
ках, а также в гранулезных, в зависимости от стадии зрелости фолликула.
Менструальный цикл
Длительность менструального цикла подвержена индивидуальным коле-
банием, но в активном репродуктивном возрасте составляет в среднем 28 дней.
Укорочение и, в большей степени, удлинение менструального цикла чаще всего
обусловлено нарушениями в гормональной продукции в связи с чем меняется
характер циклических изменений других компонентов репродуктивной системы.
«Менструальный цикл — это повторяющееся выражение деятельности систе-
мы гипоталамус-гипофиз-яичники, вызванными ею структурными и функциональ-
ными изменениями репродуктивного тракта: матки,
маточных
труб, эндометрия,
влагалища. Кульминация каждого цикла — менструальное кровотечение, пер-
вый день которого считается началом цикла» (S.Yen, R.Jaffe,
1998).
При анализе физиологических
процессов,
которые происходят у женщин
репродуктивного возраста целесообразно разделить менструальный цикл на
яичниковый, по сложным процессам
фолликулогенеза,
особенностям стероидо-
генеза с учетом степени зрелости
фолликула,
процесса овуляции, расцвета и
угасания желтого тела, и маточный, с анализом
пролиферативных
и секретор-
ных превращений, которые претерпевают органы мишени в процессе яичнико-
вого менструального цикла. Яичниковый менструальный цикл делят на 3 фазы:
фолликулярную, фазу овуляции и лютеиновую фазу цикла.
Фолликулярная или пролиферативная фаза цикла более вариабельна, чем
лютеиновая, и ее длительность определяется временем выбора доминантного
фолликула. Как полагают Peters и
соавт.,
1975;
Gougeon,1986;
Erickson,
1986 и
др. фолликулогенез начинается в позднюю лютеиновую фазу предшествующего
цикла и завершается созреванием фолликула и овуляцией. Длительность фазы
от 10 до
14
дней. В этот период идет процесс роста нескольких фолликулов, за-
тем селекция доминантного фолликула, которая определяется, в основном, ак-
тивностью биосинтеза эстрогенов в нем; затем этап созревания фолликула. Ос-
тальные фолликулы подвергаются атрезии.
Онтогенез яичников
Процесс роста и атрезии фолликулов начинается с 20 недель беременнос-
ти, и к моменту родов в яичниках девочки остается до 2 млн ооцитов. К моменту
менархе их количество уменьшается до 300 тыс. За весь период репродуктивной
15
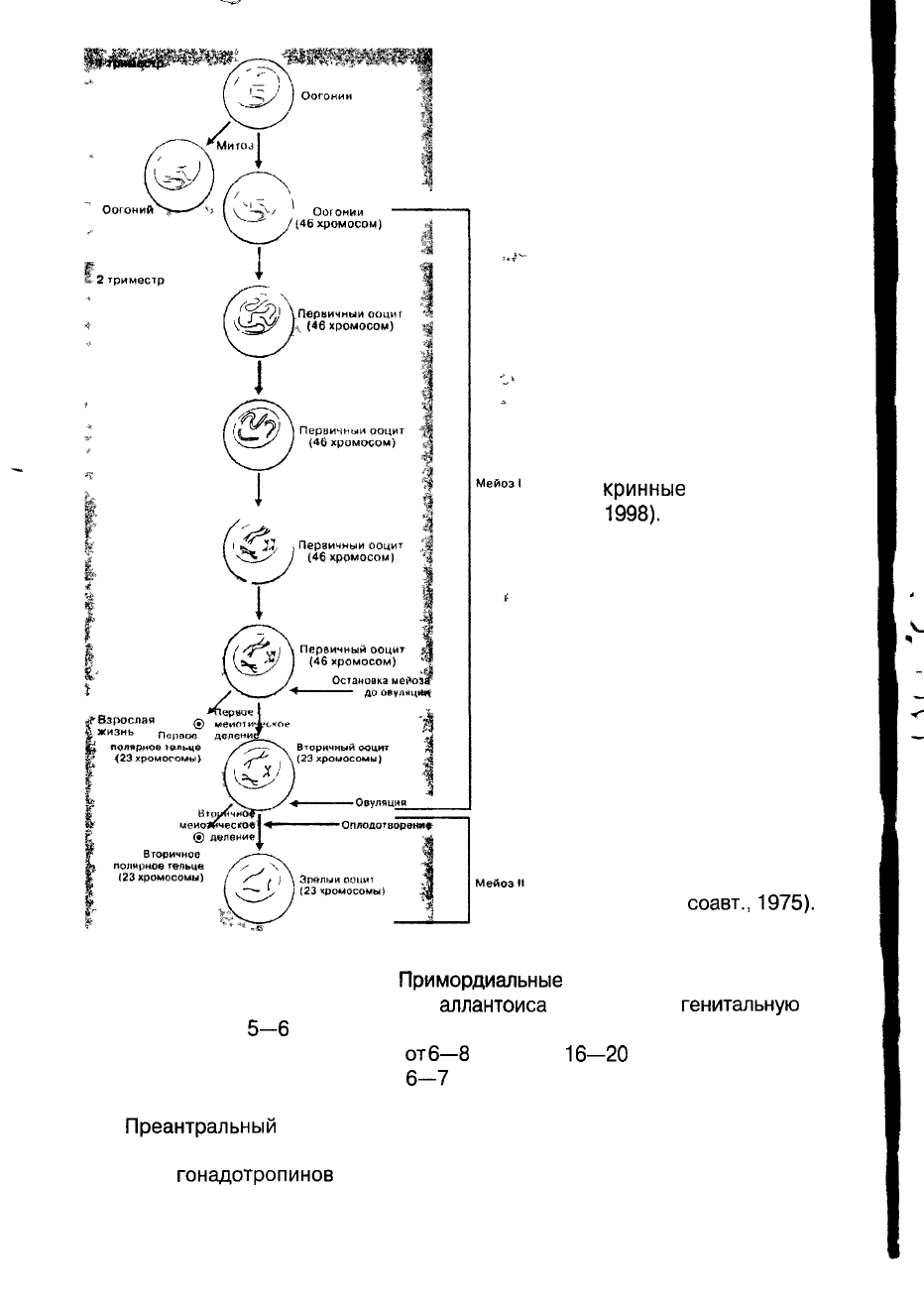
Рис. 3. Стадии развития фолликула.
жизни достигает зрелос-
ти и овулирует не более
500 фолликулов. Началь-
ный рост фолликулов не
зависит от стимуляции
ФСГ (Halpin и соавт.
1986), ограничен, и бы-
стро наступает атрезия.
Полагают, что вместо сте-
роидных гормонов основ-
ным регулятором роста и
атрезии первичных фол-
ликулов являются мест-
ные аутокринно/пара-
кринные
пептиды (Yen,
1998).
Полагают, что про-
цесс роста и атрезии
фолликулов не прерыва-
ется никакими физиоло-
гическими процессами.
Этот процесс продолжа-
ется во всех возрастах,
включая внутриутробный
период и менопаузу, не
прерывается беремен-
ностью, овуляцией и ано-
вуляцией. Механизм, ко-
торый запускает рост
фолликулов и их количе-
ство в каждом конкрет-
ном цикле, пока не ясен
(Peters и
соавт.,
1975).
В своем развитии
фолликул претерпевает
несколько стадий развития (рис. 3).
Примордиальные
зародышевые клетки проис-
ходят из эндодермы желточного мешка,
аллантоиса
и мигрируют в
генитальную
об-
ласть эмбриона на
5—6
неделе беременности. В результате быстрого митотичес-
кого деления, которое продолжается
отб—8
недель до
16—20
недель беременности,
в яичниках эмбриона образуется до
6—7
млн ооцитов, окруженных тонким слоем
гранулезных клеток.
Преантральный
фолликул — ооцит окружен мембраной (Zona pellucida).
Гранулезные клетки, окружающие ооцит, начинают пролиферировать, их рост
зависит от
гонадотропинов
и коррелирует с уровнем эстрогенов. Гранулезные
16
I
I
клетки являются мишенью
для
ФСГ. На стадии
преантрального
фолликула
грану-
лезные клетки способны синтезировать три класса стероидов: преимущественно
эстрогены, а также андрогены и прогестерон. Повышение уровня ФСГ индуцирует
увеличение числа его рецепторов, за счет роста числа гранулезных клеток ФСГ
индуцирует активность
ароматазы,
основного фермента, превращающего андро-
гены в эстрадиол. Полагают, что эстрадиол способен увеличивать число собст-
венных рецепторов, оказывая прямой
митогенный
эффект на гранулезные клетки
независимый от ФСГ. Его рассматривают как
паракринный
фактор, усиливаю-
щий эффекты ФСГ, включая активизацию процессов ароматизации (Adashi E и
соавт.,
1982;
Hsueh и
соавт.,
1983).
Рецепторы ФСГ появляются на мембранах гранулезных клеток сразу же
как начинается рост фолликула. Снижение или увеличение ФСГ ведет к измене-
нию числа его
рецепторов.
Это действие ФСГ модулируется ростовыми фактора-
ми
(Tilly
J.,
1992).
ФСГ
действует
через
G-протеин.аденилат-циклазную
систему
хотя стероидогенез в фолликуле в основном регулируется ФСГ, в этот процесс
вовлечены многие факторы: ионные каналы,
рецепторы
тирозин-киназы,
фосфо-
липазная система вторичных мессенжеров.
Роль андрогенов в раннем развитии фолликула сложна. На гранулезных клет-
ках есть рецепторы андрогенов
(Hild-Petito
и соавт.
1991).
Они являются не толь-
ко субстратом для
ФСГ-вызванной
ароматизации в эстрогены, но могут в низких
концентрациях усиливать процесс ароматизации.
Когда уровень андрогенов увеличивается,
преантральные
гранулезные
клетки
преимущественно
выбирают не путь ароматизации в эстрогены а более
простои путь превращения в андрогены через
5а-редуктазу,
превращаясь в анд-
роген, который не может быть превращен в эстроген, и таким путем ингибирует-
ся ароматазная активность. Этот процесс также ингибирует ФСГ и образование
рецепторов ЛГ, таким образом останавливая развитие фолликула.
Таким образом, низкая концентрация андрогенов усиливает их ароматиза-
цию и превращение их в эстрогены. Высокая концентрация ограничивает про-
цесс ароматизации, фолликул с высоким уровнем андрогенов
подвергается
процессам атрезии. Рост и развитие фолликула зависит от его способности пре-
вращать андрогены в эстрогены (Chabab А. и
соавт.,
1986).
Под
синергичным
действием ФСГ и эстрогенов увеличивается продукция
фолликулярной
жидкости, в межклеточном пространстве гранулезных клеток об-
разуется полость, и фолликул вступает в стадию антрального фолликула
В присутствии ФСГ доминантной субстанцией фолликулярной жидкости
будут эстрогены. При отсутствии ФСГ - андрогены. ЛГ в норме в фолликулярной
жидкости отсутствует до середины цикла. Как только увеличивается уровень
Л
Г
митотическая активность
гранулезных
клеток снижается, появляются дегенера-
тивные изменения и увеличивается уровень андрогенов в фолликуле Уровень
стероидов в фолликулярной жидкости выше, чем в плазме и отражает функцио-
нальную активность клеток яичников: гранулезных
и
тека-клеток.
Если единственной
мишенью для ФСГ являются
гранулезные
клетки,
то
у
ЛГ имеется много мишеней -
это
тека-клетки,
стромальные
и
лютеиновые
клетки и гранулезные клетки (Hsueh А и
соавт.,
1983). Способностью к стероидогенезу обладают и гранулезные и тека-клет-
ки, но ароматазная активность
преобладает^клетках
гранулезы.
В
фолликуле
Л
Г-рецептоЛБГпри
рецег торы ФСГ
Городского
центра
медицинской профилактики
т
аи
.зй-за
17

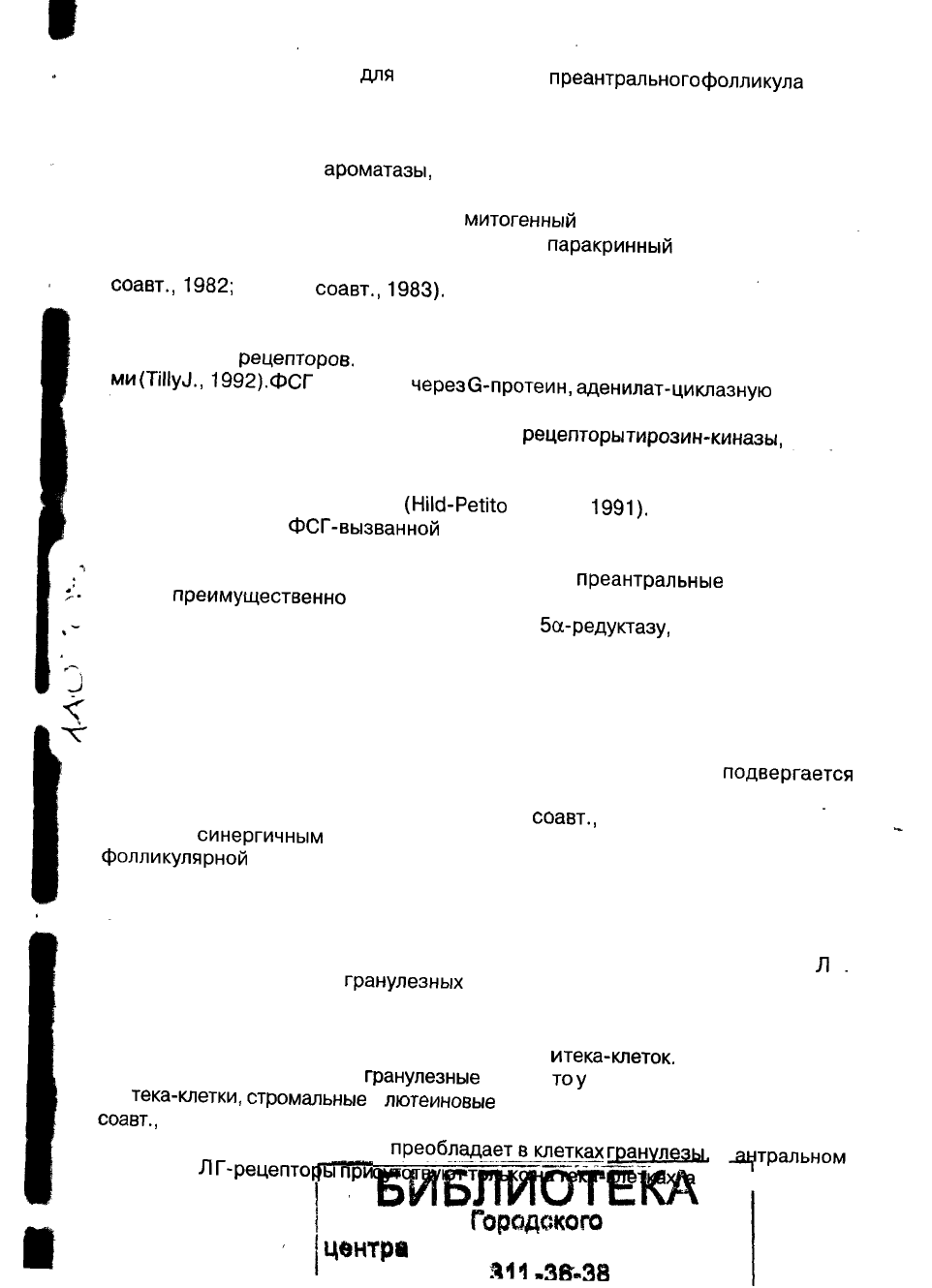
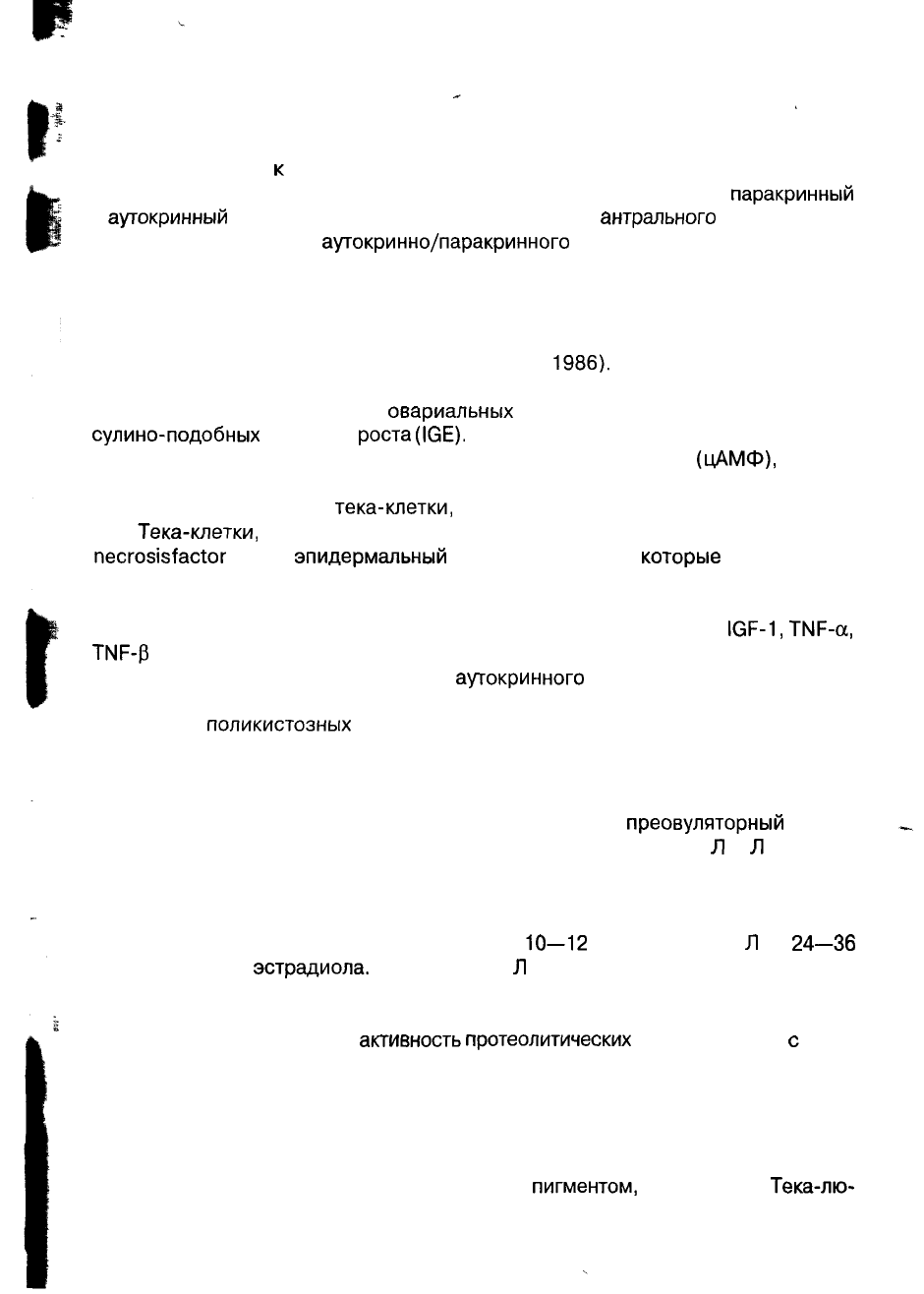
Рис. 4. Регуляция функции яичников через
аутокринные,
паракринные
и эндокринные механизмы.
s
только на гранулезных клетках. В ответ на Л Г тека-клетки продуцируют андроге-
ны,
которые затем, через ФСГ — вызванную ароматизацию, превращаются гра-
нулезными клетками в эстрогены (рис. 4).
По мере роста фолликула тека-клетки начинают экспрессировать гены для
ЛГ-рецепторов,
Р450
sec
и Зр-гидроксистероид-дегидрогеназу
(Magoffin
D.A.,
1991),
инсулино-подобный
фактор роста
(IGF-1)
синергично с ЛГ увеличивает
экспрессию гена, однако не стимулируют стероидогенез.
Яичниковый стероидогенез всегда
ЛГ-зависим.
По мере роста фолликула
тека-клетки экспрессируют
Р450с17
— энзим, который образует из холестерола
андроген. Гранулезные клетки не имеют этого энзима и зависимы от тека-клеток
в продукции эстрогенов из андрогенов. В отличие от стероидогенеза — фолли-
кулогенез
зависит от ФСГ. По мере роста фолликула и увеличения уровня эстро-
генов приходит в действие механизм обратной связи — тормозится продукция
ФСГ, что, в свою очередь, ведет к снижению ароматазной активности фолликула
18
р
I
I
I
I
I
I
и, в конечном результате, к атрезии фолликула через апоптоз (программирован-
ную гибель клетки).
Механизм обратной связи эстрогенов и ФСГ ингибирует развитие начавших
рост фолликулов, но не доминантного фолликула. Доминантный фолликул содержит
больше рецепторов
к
ФСГ, которые поддерживают пролиферацию гранулезных кле-
ток и ароматизацию андрогенов в эстрогены. Кроме этого действует
паракринный
и
аутокринный
путь, как важный координатор развития
антрального
фолликула.
Составной частью
аутокринно/паракринного
регулятора являются пептиды
(ингибин, активин, фоллистатин), которые синтезируются гранулезными клет-
ками в ответ на действие ФСГ и поступают в фолликулярную жидкость. Ингибин
снижает ФСГ секрецию; активин стимулирует высвобождение ФСГ из гипофиза
и усиливает действие ФСГ в яичнике; фоллистатин подавляет ФСГ активность,
возможно за счет связывания активина (Bicsak,
1986).
После овуляции и разви-
тия желтого тела ингибин находится под контролем ЛГ.
Рост и дифференциация
овариальных
клеток находится под влиянием ин-
сулино-подобных
факторов
роста
(IGE).
IGF-1 воздействует на гранулезные клет-
ки, вызывая увеличение циклического аденозин-монофосфата
(цАМФ),
прогес-
терона, окситоцина, протеогликана и ингибина.
IGF-1 действует на
тека-клетки,
вызывая увеличение продукции андроге-
нов.
Тека-клетки,
в свою очередь, продуцируют фактор некроза опухоли (tumor
necrosis
factor
TNF) и
эпидермальный
фактор роста (EGF),
которые
также регули-
руются ФСГ. *
EGF стимулирует пролиферацию гранулезных клеток. IGF-2 — основной
фактор роста фолликулярной жидкости, в ней также обнаружены
IGF-1,
TNF-a,
TNF-p
и EGF.
Нарушение паракринного и/или
аутокринного
регулирования овариальной
функции, по-видимому, играет роль в нарушениях процессов овуляции и в фор-
мировании
поликистозных
яичников.
По мере роста антрального фолликула увеличивается содержание эстро-
генов в фолликулярной жидкости. На пике их увеличения на гранулезных клетках
появляются рецепторы к ЛГ, происходит лютеинизация гранулезных клеток и
усиливается продукция прогестерона. Таким образом, в
преовуляторный
период
увеличение продукции эстрогенов вызывает появление рецепторов
Л
Г,
Л
Г, в свою
очередь, вызывает лютеинизацию гранулезных клеток и продукцию прогесте-
рона. Увеличение прогестерона снижает уровень эстрогенов, что, по-видимому,
вызывает второй пик ФСГ в середине цикла (рис. 5).
Полагают, что овуляция наступает через
10—12
часов после пика
Л
Г и
24—36
часов после пика
эстрадиола.
Считается, что
Л
Г стимулирует редукционное деле-
ние ооцита, лютеинизацию гранулезных клеток, синтез прогестерона и проста-
гландина в фолликуле.
Прогестерон усиливает
активность
протеолитических
энзимов, вместе
с
прос-
тагландином участвующих в разрыве стенки фолликула. Прогестероном вызванный
пик ФСГ, позволяет выходу ооцита из фолликула путем превращения плазмино-
гена в протеолитический энзим — плазмин, обеспечивает достаточное количест-
во рецепторов ЛГ для нормального развития лютеиновой фазы.
В течение 3-х дней после овуляции гранулезные клетки увеличиваются, в них
появляются характерные вакуоли, наполненные
пигментом,
— лютеином.
Тека-лю-
19
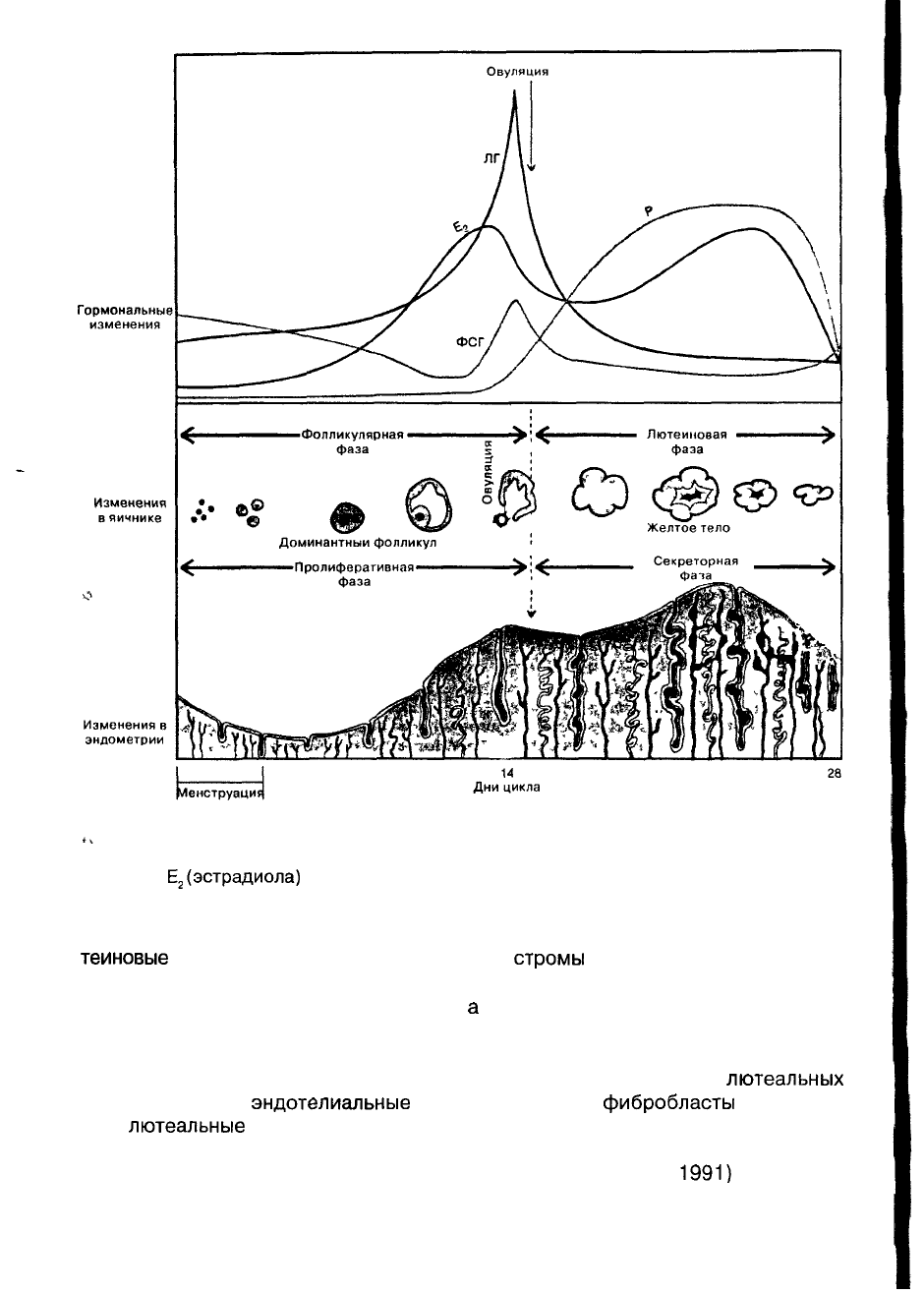
Рис. 5. Менструальный цикл: вверху — циклические изменения ФСГ, ЛГ,
Е
2
(эстрадиола)
и Р (прогестерона) относительно времени овуляции;
в середине — изменения фолликула в яичнике; внизу — изменения в эндометрии.
теиновые
клетки дифференцируются из теки и
стромы
и становятся частью желто-
го тела. Очень быстро под влиянием факторов ангиогенеза идет развитие ка-
пилляров, пронизывающих желтое тело,
а
с улучшением васкуляризации увели-
чивается продукция прогестерона и эстрогенов. Активность стероидогенеза и
длительность жизни желтого тела определяется уровнем ЛГ. Желтое тело не яв-
ляется гомогенным клеточным образованием. Помимо 2-х типов
лютеальных
клеток в нем есть
эндотелиальные
клетки, макрофаги,
фибробласты
и др Боль-
шие
лютеальные
клетки продуцируют пептиды (релаксин, окситоцин) и более
активны в стероидогенезе с большей ароматазной активностью и большим син-
тезом прогестерона, чем малые клетки (Brannian J.D. и соавт.
1991)
20
Ц
ч
I
I
k
Пик прогестерона наблюдается на 8-й день после пика ЛГ. Отмечено, что
прогестерон и
эстрадиол
в
лютеиновую фазу
секретируются
эпизодически в кор-
реляции с пульсовым выходом
Л
Г (Filicori M. и соавт.,
1984).
С образованием желтого тела контроль над продукцией ингибина перехо-
дит от ФСГ к ЛГ. Ингибин увеличивается вместе с увеличением эстрадиола до
пика
Л
Г и продолжает увеличиваться после
пика
Л
Г, хотя
уровень
эстрогенов сни-
жается. Хотя ингибин и эстрадиол секретируются гранулезными клетками, они
регулируются разными путями (Mac Naughton и
соавт.,
1992). Снижение инги-
бина в конце лютеиновой фазы вносит свой вклад в увеличение ФСГ для следую-
щего цикла.
Желтое тело очень быстро — на
9—11
день после овуляции уменьшается.
Механизм дегенерации не ясен и не связан с лютеолитической ролью эстроге-
нов или с
рецепторно-связанным
механизмом, как это наблюдается в эндомет-
рии (Press M.F. и
соавт.,
1988).
Есть другое объяснение роли эстрогенов, продуцируемых желтым телом.
Известно, что для синтеза рецепторов прогестерона в эндометрии требуются
эстрогены. Эстрогены лютеиновой фазы возможно необходимы для прогесте-
рон-связанных изменений в эндометрии после овуляции. Неадекватное разви-
тие рецепторов
прогестерона,
как следствие неадекватного содержания эстро-
генов,
является, возможно, дополнительным механизмом бесплодия и ранних
потерь беременности, другой формой неполноценности лютеиновой фазы.
Полагают, что длительность жизни желтого тела устанавливается в момент
овуляции.
И оно непременно будет подвергнуто
регрессии,
если не будет поддер-
жано ХГ в связи с беременностью.
Таким образом, регрессия желтого тела ведет к снижению уровней эстра-
диола, прогестерона и ингибина. Снижение ингибина снимает его подавляющее
влияние на ФСГ; снижение эстрадиола и прогестерона позволяет очень быстро
восстановить секрецию ГнРГ и снять механизм обратной связи с гипофиза. Сни-
жение ингибина и эстрадиола, совместно с увеличением ГнРГ дает превалирова-
ние ФСГ над ЛГ. Увеличение ФСГ приводит к росту фолликулов с последующим
выбором доминантного фолликула, и начинается новый цикл, в том случае если
не наступила беременность.
Стероидные гормоны играют ведущую роль в репродуктивной биологии и
в общей физиологии. Они определяют фенотип человека, влияют на сердечно-
сосудистую систему, метаболизм костей, кожи, общее самочувствие организма
и играют ключевую роль при
беременности.
Действие стероидных гормонов отра-
жает внутриклеточные и генетические механизмы, которые необходимы для пере-
дачи внеклеточного сигнала
к
ядру клетки, чтобы вызвать физиологический ответ.
Эстрогены диффузно проникают через клеточную мембрану и связываются с ре-
цепторами,
расположенными в ядре клетки. Комплекс рецептор-стероид затем
связывается с ДНК. В клетках мишенях эти взаимодействия приводят к экспрес-
сии генов, синтезу протеинов, к определенной функции клеток и тканей.