Радченко В.Г. Биология пчел
Подождите немного. Документ загружается.

220
Глава 12. Происхождение социального образа жизни
так как мутуалистическое объединение особей само по себе не требует отказа от
их размножения, то отбор будет благоприятствовать самкам, которые, используя
преимущества совместной жизни, как можно полнее реализуют свой репродук-
тивный потенциал.
Гипотеза мутуализма, как и ее усовершенствованный вариант – гипотеза
полигинной семьи (см. ниже), базируется на ошибочных сведениях о существо-
вании в естественных условиях видов, живущих квазисоциальными и семисоци-
альными колониями (раздел 9.1). Некоторые авторы (в частности: Starr, 1979)
вообще не рассматривают гипотезу мутуализма как отдельную теорию, из-за ее
частного характера и неясности. Мы полностью разделяем мнение Гамильтона
(Hamilton, 1964b) о невероятности не только возникновения каст в группе нерод-
ственных особей, но и какого-либо альтруизма между ними. Вместе с тем некото-
рые взгляды, изложенные Миченером в гипотезе мутуализма, способствовали
лучшему пониманию предпосылок, ведущих к возникновению эусоциальной
жизни (раздел 13.1).
Следует указать, что после появления теории отбора родичей (см. ниже) Миченер (Lin, Michener,
1972; Holldobler, Michener, 1980) в значительной степени пересмотрел свой взгляд на возможность
возникновения стерильной касты в группе неродственных особей мутуалистическим путем. Он при-
знал, что эусоциальность, по-видимому, появилась в результате взаимодействия между двумя типами
отборов: индивидуального и отбора родичей. При этом отбор родичей, по его мнению, был, наряду с
мутуализмом, задействован в становлении стерильных каст.
Отбор родичей. Ключевая идея теории, разработанной Гамильтоном (Hamil-
ton, 1963, 1964а), заключается в возможности особи передавать свои гены в
следующую генерацию не только непосредственно при размножении, но и при
выращивании близкородственных особей, в той или иной мере несущих копии ее
генов. Названная автором «теорией родства» (kinship theory) и получившая сразу
более удачное и широко распространившееся в литературе название «теория
отбора родичей» (kin selection; Maynard Smith, 1964), она, как справедливо
подчеркивают многие авторы (Starr, 1979; Andersson, 1984, и др.), привела к
революции в представлениях о происхождении социального поведения.
Предшественниками Гамильтона можно по праву считать Фишера (Fisher, 1930), который пока-
зал, что общее количество генов у особей, состоящих в близком родстве с альтруистом, может превы-
шать количество генов в геноме альтруиста (особи, которая действует на пользу других, но в ущерб
себе), а также Холдейна (Haldane, 1932, 1955) и Уиллиамсов (G.Williams, Williams, 1957), видевших
решение проблемы возникновения касты рабочих в генетическом родстве последних с репродуктив-
ными членами семьи, чье потомство рабочие особи выращивают. Все эти ценные мысли, однако, не
получили достаточного распространения, пока не появились работы Гамильтона.
Гамильтон вводит понятие совокупной приспособленности (inclusive fitness)
особи, обозначенное им как сумма прямого генетического вклада особи через
собственное потомство (т.е. индивидуальной приспособленности) и косвенного
вклада этой особи в приспособленность ее родственников. Такая формулировка
не позволяет однозначно трактовать совокупную приспособленность, что препят-
ствует выработке методов ее измерения (Oster et al., 1977; Grafen, 1982).
Более конкретное условие положено Гамильтоном (Hamilton, 1964а) в фор-
мулу, определяющую возможность отбора на альтруизм:
К*>1/r, (12.1)
где К* – отношение выигрыша особи, пользующейся услугами альтруиста, к
проигрышу альтруиста; r – генетическое сходство между этими особями.
Отметим, что в большинстве публикаций разных авторов для обозначения доли идентичных гамет
в геномах сравниваемых особей применяются понятия «родственность» (relatedness), или «родство»
(relationship), в действительности имеющие более широкий смысл (включая степень генеалогического
родства). В связи с этим для данной цели мы используем термин «генетическое сходство», который
лучше отражает суть дела.
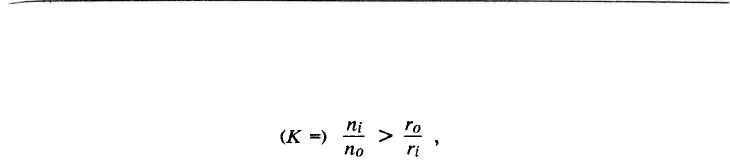
12.1. Гипотезы о механизме возникновения каст 221
(12.2)
где n – число репродуктивных потомков (п
0
– одиночной самки; n
i
– в колонии
в расчете на одного члена, участвовавшего в выращивании расплода), r – генети-
ческое сходство (r
0
– самки с собственным потомством, r
i
– члена колонии с
репродуктивным потомством колонии). При сопоставлении неравенства (12.2) с
реальными данными по насекомым (например, осам семейства Vespidae), кото-
рые тратят много времени и сил на выкармливание потомства, удобно предло-
жение Квеллера (Queller, 1989) вместо n использовать произведение числа
откладываемых самкой яиц на вероятность вырастить потомство до имаго.
Из формулы (12.2) следует, что отбор родичей тем успешнее противостоит
индивидуальному (эгоистичному) отбору, чем выше r
i
– генетическое сходство
альтруиста с потомством особи, которой он помогает, и чем больше K – выигрыш
в числе потомков, выведенных благодаря помощи альтруиста (n
i
). Если взять
случай r0 = ri (это происходит при r0 = r
i
= 1/2, когда дочь помогает выращивать
потомство обоих полов своей матери от того же отца, чью гамету носит самка),
т.е. когда особь одинаково сходна генетически как со своим потомством, так и с
потомством близкородственной особи, то механизм отбора родичей, «толкающий»
рассматриваемую особь к альтруизму, в частности к статусу рабочей, может
сработать только при K > 1, т. е. при n
i
> п
0
. В случае, когда альтруист выращивает
потомство особи, находящейся с ним в менее близком родстве, действие меха-
низма отбора родичей соответственно требует значительно большего превышения
n
i
– числа потомков в семье в расчете на одну особь, над п
0
– числом потомков,
которых она могла бы вырастить самостоятельно.
Например, чтобы самка у организмов с обычным (диплодиплоидным) типом определением пола
отказалась от собственного размножения в пользу своей сестры (т.е. для выращивания племянниц и
племянников, генетическое сходство с которыми у нее составляет r
i
= 1 /4) должно быть K > 2, а чтобы
она стала выращивать потомство своей кузины (r
i
= 1/8) = K > 4. Альтруизм в пользу неродственной
особи предполагает вообще астрономические значения K.
Величина K = n
i
/п
0
дает интегральную оценку преимуществ семейной жизни,
которые возникают у объединившихся особей благодаря организации постоянной
охраны гнезда, улучшению ухода за потомством, разделению труда и т.п. Ясно,
что вычисление реального значения K у какого либо вида насекомых для подста-
новки в формулу (12.2) условия отбора на альтруизм очень сложно, так как
сопряжено с определением общего биологического успеха особей, зависящего от
многих факторов.
Это, судя по всему, хорошо понимал и Гамильтон. Во всяком случае, развивая
свою теорию применительно к эусоциальным насекомым (Hamilton, 1964b), он
для разрешения критических вопросов даже не пытался привлекать количествен-
ные данные по репродуктивному успеху особей, чтобы оценить их совокупную
приспособленность. Не делали подобного и последующие авторы. В многочислен-
ных публикациях, посвященных обсуждению или приложению отбора родичей к
тем или иным ситуациям, реальные значения (точнее теоретически ожидаемые)
подставляются лишь в правую часть неравенства (12.2), т.е. для величин, отра-
жающих генетическое сходство особи со своим потомством (r
0
) и с репродуктив-
ным потомством колонии (r
i
). Такой прием, который широко используется в
дальнейшем тексте и нами, позволяет определить, какую величину должно пре-
Применительно к сопоставлению одиночных и эусоциальных насекомых фор-
мула Гамильтона уточнена Крэйгом (Craig, 1975) и Вест-Эберхард (West-Eber-
hard, 1975). Отказ особи от собственного размножения в пользу выращивания
чужого потомства в соответствии с теорией отбора родичей возможен при условии:
222
Глава 12. Происхождение социального образа жизни
вышать K, чтобы отбор родичей мог благоприятствовать появлению альтруисти-
ческого поведения у особи с определенной (заданной) жизненной стратегией,
характеризующейся введенными в формулу (12.2) значениями r
0
и r
i
.
Таким образом, в рамках теории отбора родичей на величину K = n
i
/n
0
не
вводится никаких ограничений, а эмпирическая оценка K для реальных видов
очень сложна и требует многолетних широкомасштабных экспериментов. Поэто-
му условие отбора родичей, показанное неравенством (12.2), скорее выражает
собой общетеоретическое представление о возможности возникновения эусоци-
альности, чем эмпирически проверяемое предсказание.
В этом смысле условие отбора родичей сходно по логической структуре с
правилом Спенсера о «выживании наиболее приспособленных», которое поло-
жено Дарвиным в основу теории естественного отбора. Действительно, условие
протекания классического индивидуального отбора можно выразить как благо-
приятствование особи x в том случае, если она оставит больше потомков, которые
достигли репродуктивного возраста, чем особь у, т.е. как
n
x
> n
y
. (12.3)
Тогда теория отбора родичей, согласно (12.2), определяет аналогичное усло-
вие, но в отношении числа копий гамет, передаваемых особями в следующую
генерацию:
n
i
r
i
>n
0
r
0
.
(12.4)
По формальной записи условие (12.4) протекания отбора родичей шире усло-
вия (12.3) для классического индивидуального отбора, так как последнее является
частным случаем (12.4) при r
i
= r
0
.
Отметим, что некоторые авторы (например: Craig, 1979; Starr, 1979) ошибочно выделяют значе-
ние K = 1 /2 в качестве теоретического минимума K, при котором отбор родичей не может идти ни при
каких обстоятельствах. Данный вывод основан на представлении, что минимум в значении правой
части неравенства 12.2 достигается у матки, в колонии которой после выводка рабочих особей выра-
щивается репродуктивное потомство, состоящее только из ее детей (генетическое сходство матки с
ними составляет 1/2; подробности вычисления этого и других показателей, приводимых в настоящем
разделе, см. в разделе 12.3, а также на рис. 143 и 144 и в табл. 6). При этом матка сопоставляется с
одиночной самкой, вместо рабочего выводка выращивающей детей, которые затем дают потомство
следующей генерации, т.е. внуков рассматриваемой одиночной самки (с ними у нее r = 1/4). Подста-
вив эти значения r в формулу 12.2 получим K > (1/4 : 1/2 =) 1/2. В действительности отбор родичей
может теоретически идти при любых значениях K > 0, если учесть, что матка может производить не
один, а сколько угодно рабочих выводков перед репродуктивным. Например, если матка выращивает
два выводка рабочих особей, то ее генетическое сходство с репродуктивным потомством (по-прежнему
r = 1/2) должно сопоставляться со сходством одиночной самки со своими правнуками (r = 1/8), откуда
K
>
(1/8
:
1/2=) 1/4, и
т.д.
Несмотря на указанную выше неэмпиричность общей теории отбора родичей,
сопоставление условия (12.2), при котором отбор благоприятствует появлению
альтруизма, с данными о встречаемости эусоциальности среди насекомых позво-
ляет сделать, по крайней мере, один очень важный вывод, отмеченный уже
Гамильтоном и многими последующими авторами. Он заключается в том, что в
естественных условиях величина K не может стабильно превышать единицу.
Иными словами, помощь альтруистов не может приводить к тому, чтобы репро-
дуктивное потомство, выведенное в образовавшейся колонии, по количеству
превышало сумму потомков, которых все члены колонии могут вырастить само-
стоятельно. Такое превышение для поддержки отбором аллели альтруизма долж-
но, согласно результатам популяционно-генетического моделирования (Craig,
1979), составлять всего 6% (т.е. K > 1.06). Судя по косвенным данным, величина
K не может достигать стабильных значений выше единицы, по крайней мере, в
течение периода времени, достаточного для эволюционных сдвигов к эусоциаль-
ности.

12.1. Гипотезы о механизме возникновения каст 223
Такой вывод однозначно вытекает из факта отсутствия эусоциальности среди
всех насекомых с обычным диплодиплоидным механизмом определения пола, в
результате чего братья и сестры в среднем одинаково генетически сходны как
между собой, так и с сибсом противоположного пола. У таких организмов отно-
шение генетического сходства с собственным потомством (r
0
) к генетическому
сходству альтруиста с потомством родственной особи, которой он помогает (r
i
), не
может быть меньше единицы, причем r
0
/r
i
= 1 достигается лишь когда особь
отказывается от собственной репродукции в пользу матери для воспитания своих
братьев и сестер от того же отца (r
0
= r
i
= 1/2). Напротив, все группы, у которых
известна эусоциальность, характеризуются особенным механизмом определения
пола, приводящим к асимметрии в генетическом сходстве между братьями и
сестрами, – у перепончатокрылых это гаплодиплоидия (раздел 1.3), а у термитов
– система транслокаций половых хромосом (раздел 12.2). Действие отбора роди-
чей у этих насекомых существенно облегчено, так как генетическое сходство
самки с сестрами (а у термитов также и самца с братьями) выше, чем с собствен-
ными детьми. В результате отбор благоприятствует распространению аллели
альтруизма у перепончатокрылых и термитов в случае, когда особь отказывается
от собственного размножения в пользу матери уже при K > 0.8 (раздел 12.2).
Приложение теории отбора родичей к перепончатокрылым в дальнейшем
тексте нам удобнее рассматривать не только как ее конкретизацию и усиленный
вариант, но и как самостоятельную гипотезу под названием «гипотезы гаплодип-
лоидии», противопоставляя ее общей теории отбора родичей Гамильтона, которая
не дает тестируемых предсказаний в отношении эусоциальности. Гипотеза гап-
лодиплоидии в качестве единственного реального механизма возникновения
эусоциальности, принимаемого нами, подробно излагается в разделах 12.2 и 12.3.
Она доказывает возможность существенного повышения совокупной приспособ-
ленности самок, когда они становятся рабочими у своей матери, и тем самым
служит теоретической основой субсоциального пути становления эусоциально-
сти, ведущего к возникновению материнско-дочерних колоний (раздел 13.1).
Гипотеза полигинной семьи. Многие исследователи не признают гипотезу
гаплодиплоидии, а вместе с ней путь возникновения эусоциальности через мате-
ринско-дочерние колонии, поскольку не могут в рамках этой гипотезы объяснить
известные случаи полигинного основания эусоциальных колоний. Они продолжа-
ют поддерживать идею Миченера, высказанную им в ранних работах (Michener,
1958, 1969а, и др.), о парасоциальном пути, согласно которой эусоциальные
колонии возникали первично как объединение самок одной генерации. Развитие
гипотезы мутуализма Миченера за счет подключения к ней теории отбора роди-
чей привело к представлению о том, что на начальном этапе становления эусоци-
альности колонии представляли собой группы совместно живущих сестер, одна
из которых становилась яйцекладущей, а остальные полностью или частично
отказывались от собственной репродукции.
Подобные взгляды в наиболее четкой форме изложены в гипотезе «полигин-
ной семьи» (polygynous family), разработанной Вест-Эберхард (West-Eberhard,
1978а, 1981). Согласно этой гипотезе, дополнительной предпосылкой к возникно-
вению каст служит присоединение в одном гнезде репродуктивно слабых сестер к
сильной, при этом появление нерепродуктивной касты рабочих не связывается с
обязательной встречей матери со своими дочерьми. Гипотеза полигинной семьи в
последнее 10-летие получила широкое распространение среди социобиологов и
поддерживается как авторами ряда современных обзоров по эволюции эу социаль-
ности у насекомых (Andersson, 1984; Fletcher, Ross, 1985; Gadagkar, 1985a,
224 Глава 12, Происхождение социального образа жизни
1985b), так и отдельными исследователями (Forsyth, 1980; Noonan, 1981; Stras-
smann, 1981; Pollock, 1983; Eickwort, 1986; Packer, 1986c; Queller et al., 1988, и
др.); многие авторы высказывали сходные взгляды и раньше (например: Gibo,
1974, 1978; Litte, 1977; Metcalf, Whitt, 1977).
В гипотезе полигинной семьи Вест-Эберхард делает попытку объяснить меха-
низм возникновения эусоциальности на основе теории отбора родичей путем ее
приложения к конкретной ситуации – к семье, состоящей из сестер, одна из
которых становится яйцекладущей. Эта ситуация может быть в явном виде опи-
сана как параметрами родственных отношений между членами примитивной
эусоциальной колонии (сестры), так и параметрами их генетического сходства с
репродуктивным потомством, выращиваемым в колонии: потомство колонии со-
стоит из детей яйцекладущей самки (с ними у нее r = 1/2), которые являются
племянниками и племянницами для самок, ставших рабочими при своей яйце-
кладущей сестре (у таких самок для гаплодиплоидных организмов среднее гене-
тическое сходство с потомством колонии равно 3/8, см. рис 144 в разделе 12.2).
Поэтому в отличие от общего условия (12.2) отбора родичей гипотеза полигинной
семьи является вполне эмпиричной, и из нее дедуктивным способом можно вы-
вести следствия-предсказания в отношении приведенных выше параметров, про-
верив которые можно оценить непротиворечивость гипотезы эмпирическим
данным.
Следствия, выводимые из гипотезы полигинной семьи, формулируются
следующим образом. Во-первых, отбор родичей должен благоприятствовать рас-
пространению аллели альтруизма у самок гаплодиплоидных организмов, отка-
зывающихся от собственного размножения в пользу своей сестры. Во-вторых,
у видов, недавно перешедших к социальному образу жизни, колонии должны
проходить полный цикл развития как объединение близкородственных самок
одной генерации. Покажем, что оба эти следствия предсказывают явления, кото-
рые не наблюдаются в действительности и, следовательно гипотеза полигинной
семьи не может адекватно описать механизм и путь возникновения эусоциаль-
ности у насекомых.
В отличие от гипотезы гаплодиплоидии (раздел 12.2), также являющейся
конкретизацией теории отбора родичей, гипотеза полигинной семьи не только
смягчает, но даже ужесточает условие, при котором этот отбор может действо-
вать. Так, совокупная приспособленность самки, отказавшейся от собственного
размножения в пользу своей сестры, будет выше ее индивидуальной приспособ-
ленности только при К > 1.33 (см. табл. 6 в разделе 12.2 и обсуждение стратегии
«сестра» в разделе 12.3), т. е. в случае, когда на каждую из сестер в колонии будет
произведено в 1.33 раза больше потомков, чем может вывести одиночная самка.
Выполнение этого условия представляется невероятным, поскольку даже условие
К > 1 (отбор родичей, ведущий к возникновению материнско-дочерних сообществ
у диплодиплоидных организмов, см. выше) не реализуется в природе, на что
указывает отсутствие эусоциальности у обычных диплодиплоидных насекомых.
Другой аргумент против гипотезы полигинной семьи заключается в том, что
она, как и мутуалистическая гипотеза Миченера, предполагает существование
семисоциальных видов, т.е. видов, образующих не материнско-дочерние семьи, а
колонии, состоящие из сестер. Такие виды в природе не обнаружены ни среди пчел
(гл. 9-11), ни среди ос (см. разбор данных работы Вест-Эберхард ниже), но зато
известно много субсоциальных видов, колонии которых справедливо рассматри-
ваются как начальный этап в становлении эусоциальности по субсоциальному
пути, отвергаемому Вест-Эберхард и ее сторонниками.
12.1. Гипотезы о механизме возникновения каст
225
С целью подкрепить свою гипотезу максимальным количеством фактов Вест-Эберхард (West-
Eberhard, 1978а) приводит список 39 видов ос (Pompilidae – 2 вида, Sphecidae – 27, Masaridae – 1,
Eumenidae – 3, Vespidae – 8), которые предположительно образуют квази- или семисоциальные
колонии. Правда, термин «квазисоциальные виды» Вест-Эберхард не использовала, а называла их
«примитивно социальными». При этом в список ошибочно попали один несомненно эусоциальный вид
(Microstigmus comes; Matthews, 1968; Matthews, Ross, 1988) и еще, по крайней мере, один вид (Steno-
gaster concina; Spradbery, 1975), для которого можно предположить наличие эусоциальности. Все
остальные виды, как это следует из текста статьи Вест-Эберхард, приведены фактически потому, что
для них отмечены случаи совместного проживания нескольких самок в одном гнезде. Среди них
неперекрывание генераций, предполагающее возможность «примитивной социальности», точно ус-
тановлено лишь для 5 видов, но для 4 из них (Сerсегis simplex, Philanthus gibbosus, Sphecius speciosus,
Tachpyus petiolatus) неясно число самок, откладывающих яйца при совместном поселении в одном
гнезде. Только для Sphex ichneumoneus, насколько можно понять из текста, зарегистрировано совме-
стное заполнение кормом одной ячейки двумя самками, из которых особь, приносившая последнюю
жертву, по-видимому, откладывала яйцо. В своих последующих публикациях Вест-Эберхард также
не приводит каких либо достоверных сведений о существовании «примитивно социальных» видов, и
при этом указывает, что информация о таких видах остается очень скудной (см. например: West-
Eberhard, 1987). В частности, добавляя к числу «примитивно социальных» видов Xenorhynchium
nitidulum (Eumeninae), она признается, что гнезда данного вида располагаются одно возле другого так
близко, что трудно утверждать, помогают ли самки друг другу или нет.
Совместное проживание нескольких особей в одном гнезде (образование составных гнезд), как и
совместное заполнение кормом одной ячейки разными особями, а иногда и просто паразитические
тенденции, когда самка откладывает яйцо в чужую ячейку, изредка наблюдаются и у пчел (разделы
9.1 и 9.2). Все эти явления носят случайный характер и поэтому не могут однозначно интерпретиро-
ваться как наличие у данных видов истинно парасоциального поведения.
В качестве частного замечания отметим также неверный подход Вест-Эбер-
хард (West-Eberhard, 1978b, p. 441) к оценке совокупной приспособленности
самки, которую она делает лишь на период, когда самка принимала участие в
выращивании потомства, т.е. по отрезку времени, искусственно вырванному из
полного цикла развития колонии. Такой подход ведет, в частности, к тому, что те
из самок, выполнявших функции рабочих особей, которые не дожили до выведе-
ния репродуктивного выводка, вообще не имеют совокупной приспособленности.
Основной довод в пользу гипотезы полигинной семьи – объяснение случаев
совместного основания колонии несколькими сестрами – теряет свое значение в
связи с тем, что данное явление теперь находит, на наш взгляд, вполне убедитель-
ное обоснование в рамках гипотезы гаплодиплоидии. Этот вопрос подробно рас-
сматривается в разделе 12.3, где также показано, что полигинное основание
колонии действительно выгодно сестрам, даже выполняющим функции рабочих
особей, но только у эусоциальных видов, имеющих настоящую рабочую касту из
дочерей матки.
Родительская манипуляция. Гипотеза родительской манипуляции (parental
manipulation) выдвинута Алекзандером (R.Alexander, 1974) первоначально как
противопоставление гипотезе гаплодиплоидии, поскольку подразумевала невоз-
можность альтруизма детей по отношению к своей матери. Соавторство этой
гипотезы обычно приписывают также Миченеру и Бразерсу (Michener, Brothers,
1974), хотя изложенный ими подход заметно отличается от гипотезы Алекзанде-
ра.
Миченер и Бразерс (Michener, Brothers, 1974) в действительности лишь поддержали теоретиче-
ские соображения Алекзандера, используя при этом вместо «родительской манипуляции» термин
«контроль матки» (queen control), и привели ряд дополнительных данных, свидетельствующих, по их
мнению, в пользу гипотезы. На самом деле Миченер и Бразерс применили идею Алекзандера не к
материнско-дочерним (т.е. настоящим эусоциальным) колониям, а к «семисоциальным», т.е. состоя-
щим из самок одной генерации, между которыми наблюдается явная борьба за доминирование с
правом откладывать яйца. Эти данные, однако, получены на искусственно созданных полигинных
колониях галиктин (раздел 10.1), которые часто составлялись из неродственных особей. Миченер не
придает этому значения и даже не видит разницы между настоящей маткой и доминирующей самкой
в группе особей одной генерации, называя «матками» или «царицами» обеих, так как последовательно
(см.: Michener, 1958, 1969а, 1974, 1985b, 1990а) отрицает субсоциальный путь возникновения эусо-
циальности (который, кстати, лежит в основе гипотезы родительской манипуляции; см. также раздел
13.1) для галиктин. К сожалению, многие из выводов, полученных на искусственных колониях, были
226 Глава 12. Происхождение социального образа жизни
некритически перенесены на настоящие материнскодочерние семьи и в таком ошибочном виде
попали в современные обзоры (например: Starr, 1979; Andersson, 1984; Fletcher, Ross, 1985; Кипятков,
1986, и др.).
Гипотеза родительской манипуляции объясняет возникновение эусоциально
сти у насекомых тем, что мать с помощью различных приемов доминирования и
контроля за своими дочерьми подавляет их репродуктивные способности, при
нуждая становиться рабочими. Самка, имеющая дочерейпомощниц, естествен
но получает селективное преимущество по сравнению с одиночными самками.
Возникающий при этом конфликт генетических интересов, связанный с пониже
нием индивидуальной приспособленности рабочих особей, насильно решается в
свою пользу маткой, стремящейся максимально повысить собственную приспо
собленность. При этом отбор должен содействовать самкам, которые производят
послушное рабочее потомство, помогающее ей в воспитании нормального репро
дуктивного потомства.
Предположение о возможности появления социальной жизни у насекомых в результате вынуж
денного подчинения дочерей своей матери первым в достаточно отчетливой форме выдвинул Легеви
(Legewie, 1925а, 1925b). Такую возможность он обосновывал «гипогенитализмом» (hypogenitalism)
части дочерей, имеющих недоразвитую репродуктивную систему и поэтому склонных помогать своей
матери. Недоразвитость самок Легеви объяснял их недокормленностью. В развитии этой гипотезы еще
одну интересную мысль высказал Ричардс (O.Richards, 1953), считавший, что матки эусоциальных
перепончатокрылых благодаря способности регулировать пол своего потомства могут в первых вывод
ках производить одних дочерей, которым матки препятствуют копулировать, чем добавочно стимули
руют дочерей становиться рабочими особями.
Сразу после своего появления гипотеза подверглась серьезной критике по
целому ряду пунктов.
Вопервых, она совершенно игнорирует генетические интересы детей, стано
вящихся рабочими особями. Созданные различные генетические модели развития
эусоциальности путем родительской манипуляции показывают, что отбор роди
телей, сильнее подавляющих репродуктивные способности своих детей, действи
тельно может благоприятствовать развитию социальности (Charlesworth, 1978),
однако его действие блокируется отбором между потомками, направленным на
сохранение их репродуктивных возможностей (Trivers, 1974; Craig, 1979). В
результате мать будет только тратить ресурсы, производя субфертильных, но
всегда полностью «эгоистичных» дочерей (Craig, 1979; Crozier, 1979).
Вовторых, родительская манипуляция у гаплодиплоидных организмов дол
жна превращать самцов в рабочих особей так же легко, как и у диплоидных
(Hamilton, 1975; Craig, 1982), чего никогда не происходит. Правда, Крэйг (Craig,
1982) отмечает, что главной причиной отсутствия касты рабочих самцов является
не особый коэффициент генетического сходства, делающий для них невыгодным
принимать какоелибо участие в выращивании своих братьев и сестер в ущерб
собственному размножению, а неприспособленность самцов к выполнению этих
функций. Гипотеза родительской манипуляции также не в состоянии ответить на
вопрос почему эусоциальность так часто возникала среди перепончатокрылых и
всего один раз (у термитов) возникла среди других насекомых, хотя механизм,
постулируемый этой гипотезой, должен срабатывать в самых разнообразных
группах.
Втретьих, с помощью гипотезы родительской манипуляции очень трудно
объяснить причины разделения сестер на касты. Попытка Алекзандера (R.Alexan
der, 1974) объяснить этот случай дифференцированным программированием
своих дочерей, решительно отвергнута последующими исследователями (West
Eberhard, 1975; Crozier, 1977; Craig, 1979; Starr, 1979: Hölldobler, Michener, 1980;
Andersson, 1984; Кипятков, 1986, и др.) изза отсутствия механизмов подобного
воздействия. Некоторые из приведенных авторов (Starr, 1979, 1985; Кипятков,
12.1. Гипотезы о механизме возникновения каст 227
1986, 1991), тем не менее, поддерживают гипотезу родительской манипуляции,
уклоняясь при этом от ответа на вопрос о механизме разделения сестер на касты
при полигинном основании колонии.
Критика гипотезы родительской манипуляции по указанным пунктам, хотя
и справедлива, но, на наш взгляд, недостаточна, так как она основана прежде
всего на том, что гипотеза не дает удовлетворительного объяснения некоторым
явлениям. В принципе такая критика может быть снята добавлением к гипотезе
дополнительных утверждений. Гипотеза родительской манипуляции по своему
содержанию и логической структуре является вполне эмпирической, поэтому
лучший способ ее проверки на истинность – это выведение из гипотезы дедуктив-
ным способом следствий-предсказаний и их тестирование фактическими данны-
ми. С позиции гипотезы родительской манипуляции можно сделать, по крайней
мере, три предсказания, которые ранее в литературе специально не рассматрива-
лись.
Во-первых, если эусоциальность возникает в результате родительской мани-
пуляции, то подавление маткой своих дочерей должно быть наиболее сильно (и
обязательно!) выражено у примитивно-эусоциальных видов.
Во-вторых, одним из главных и наиболее очевидных способов родительской
манипуляции является препятствование спариванию дочерей путем исключения
самцов из всех выводков, кроме репродуктивного; следовательно, по крайней
мере, у наиболее примитивных в социальном отношении пчел и ос матки для
рабочих выводков должны откладывать только диплоидные («самковые») яйца.
В-третьих, если матке удается благодаря манипуляции заставить дочерей
отказаться от откладки яиц для выведения самок в репродуктивном выводке, то с
таким же успехом она должна препятствовать выведению ими сыновей.
Анализ данных по биологии эусоциальных насекомых показывает, что ни
одно из приведенных следствий гипотезы родительской манипуляции не соответ-
ствует реальным фактам. Поведенческое и феромонное доминирование матки
над дочерьми как раз наиболее слабо выражено или вообще не выявлено у наибо-
лее примитивных в социальной организации колоний пчел (данные по цератинам
см. в разделе 9.4, по галиктинам – в разделе 10.3) и ос (данные по Polistes см.,
например: Гречка, 1986), хотя у этих видов при полигинном основании колонии
сестрами между ними наблюдается явная агрессивность, которая исчезает лишь
после установления иерархии доминирования. У этих же видов в рабочем выводке
значительная доля принадлежит самцам. Лишь только в наиболее организован-
ных колониях среди примитивно-эусоциальных видов (например, у шмелей –
раздел 11.5 и некоторых галиктин – раздел 10.3) и в колониях продвинуто-эусо-
циальных видов мелипонин (раздел 9.5) и апин (раздел 9.6), так же как у муравь-
ев и многих ос-веспин, родительская манипуляция достигает высокого развития
и в рабочих выводках полностью отсутствуют самцы. Вместе с тем даже у таких
видов самцы в репродуктивном потомстве, как правило, выращиваются из не-
оплодотворенных яиц, отложенных рабочими особями (см. «конфликт при выве-
дении самцов» в разделе 12.2).
Как показали многие авторы (Craig, 1979; Crozier, 1979; Maynard Smith, 1982;
Michod, 1982; Andersson, 1984, и др.), гипотеза родительской манипуляции, хотя
сама по себе и недостаточна для объяснения появления нерепродуктивных каст,
но не является альтернативой к теории отбора родичей, поскольку подразумевает
близкое родство особей (родители – дети). Родительская манипуляция (или кон-
троль матки) сама по себе не может приводить к возникновению стерильной
касты, так как индивидуальный отбор всегда успешно противостоит попыткам
228
Глава 12. Происхождение социального образа жизни
родительской манипуляции, благоприятствуя дочерям, которые либо не отказы-
ваются от собственной репродукции при создании семейной колонии, либо вообще
уклоняются от воздействия матери и поселяются одиночно.
Этот вывод, однако, не означает, что родительской манипуляции вообще не
существует. Просто такая манипуляция, отвечающая интересам матки, может
появляться только после возникновения эусоциальности (раздел 13.2). Более
того, родительская манипуляция, осуществляемая с помощью феромонов, пове-
денческих акций или через ограничение корма (разделы 9.5, 9.6, 10.3, 11.5) и
направленная на подавление матерью репродуктивных способностей ее дочерей
и их принуждение к выполнению функций рабочих особей, очень широко распро-
странена в эусоциальных колониях и является важным фактором в эволюции
эусоциальности.
Недавно Кипятков (1986) предпринял объединение гипотез отбора родичей, родительской мани-
пуляции и мутуализма, которое назвал «новой синтетической теорией» эволюции эусоциальности у
насекомых. Как отмечалось выше, мутуализм и родительская манипуляция в качестве факторов
способствующих возникновению эусоциальности, связаны с совершенно разными представлениями о
путях становления – соответственно с парасоциальным и субсоциальным, который в основном прини-
мается Кипятковым.
Объединение гипотезы мутуализма и теории отбора родичей, проведенное ранее Вест-Эберхард,
хотя и основано на отвергаемом нами парасоциальном пути, привело к созданию вполне логичной и
внутренне непротиворечивой гипотезы полигинной семьи. Присоединение же мутуализма к родитель-
ской манипуляции, сделанное Кипятковым, не только искусственно, но неверно по сути: мутуалисти-
ческие (т.е. взаимополезные) отношения между матерью и ее дочерьми, которых она принуждает
стать рабочими особями, просто не могут возникнуть. В результате «синтетическая теория» Кипяткова
по существу не отличается от собственно гипотезы родительской манипуляции, обсуждение которой
почти все авторы с самого начала вели с позиции теории отбора родичей (например: Charlesworth,
1978; Charnov, 1978; Craig, 1979).
12.2. Гипотеза гаплодиплоидии
Открытие Гамильтона. Как отмечалось в разделе 1.3, у гаплодиплоидных
организмов за счет арренотокичного партеногенеза самцы получаются из неопло-
дотворенных гаплоидных яиц, тогда как самки – из оплодотворенных диплоидных
(рис 143). Это явление для перепончатокрылых насекомых, в частности у медо-
носной пчелы, было описано еще в середине прошлого века (Dzierzon, 1845).
Открытие Гамильтона (Hamilton, 1964b), впоследствии получившее название
«гипотезы гаплодиплоидии» или «гипотезы 3/4 родства» (например: West-Eber-
hard, 1978а), заключается в следующем. Поскольку самец происходит из гапло-
идного яйца и соответственно имеет в своем геноме гаметы лишь одного типа, то
в геноме всех его дочерей обязательно содержится эта гамета. Второй же гаметой,
получаемой дочерьми от матери, будет одна из ее двух гамет. В результате среднее
генетическое сходство (т.е. доля идентичных гамет, в дальнейшем тексте обозна-
чается буквой r) между сестрами (по матери и по отцу) равно 3/4 (рис. 143 и 144).
Генетическое сходство матери со своими детьми составляет лишь 1/2, так как все
они несут только по одной из ее гамет.
Таким образом, при прочих равных условиях (прежде всего при одинаковом
числе потомков в среднем на одну самку), благодаря асимметрии в распределении
гамет в потомстве, самке генетически выгоднее отказаться от собственного paз-
множения в пользу выращивания своих сестер и стать рабочей особью у своей
матери. Одновременно это выгодно и для матери: становясь маткой, она может в
репродуктивном потомстве колонии взамен внуков (с ними у нее r = 1/4, рис 144),
которые были бы у нее к концу сезона при одиночной жизни, получить своих детей
(имеющих с ней r = 1/2). Именно такой взаимный интерес и послужил основной
причиной возникновения эусоциальности у перепончатокрылых насекомых.
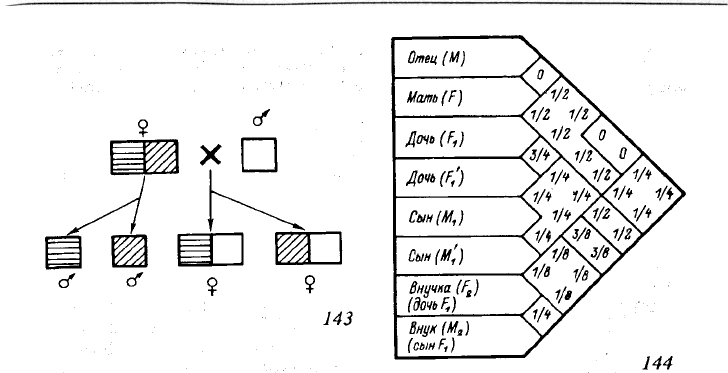
12.2. Гипотеза гаплодиплоидии
229
Рис. 144. Среднее генетическое сходство (доля идентичных гамет) между родителями
и их потомками при арренотокичном партеногенезе
Среднее генетическое сходство указано в местах перекрещивания диагоналей, которые идут от
соответствующих граф с обозначением родственников. Уточнено по сравнению с предыдущими пуб-
ликациями (Hamilton, 1964b; Wilson E., 1971) среднее генетическое сходство между матерью (F) и
внучкой (F
2
). Среднее генетическое сходство между братьями (M
1
и М'
1
) принято r = 1/4, что
соответствует расчету Гамильтона (Hamilton, 1964b) и подтверждается подсчетом числа гамет, пере-
дающихся в следующую генерацию при заданном числе потомков, а не r = 1/2, как это делали
некоторые авторы (например: Crozier, 1979; Кипятков, 1985), исходя из того, что при наличии у
самцов одинаковой (и единственной) гаметы генетическое сходство между ними равно 1 (а не 1/2, как
следует считать).
Отбор родичей у особей с разными стратегиями. В рамках общей теории
отбора родичей (раздел 12.1) указанные особенности генетического сходства меж-
ду гаплодиплоидными организмами существенно смягчают условия, при которых
отбор родичей может вести к возникновению эусоциальности. В таблице 6 приве-
дены значения отношения генетического сходства, ожидаемые при моноандрии
(когда матки копулируют только с одним самцом), которые соответствуют правой
части формулы (12.2). Эти значения показывают ту величину, которую должно
превосходить отношение числа потомков в расчете на одну особь к числу потомков
одиночной самки (т.е. K или левая часть неравенства 12.2), чтобы отбор родичей
благоприятствовал стратегии, «выбранной» данной особью. Помимо данных по
гаплодиплоидным организмам, в таблицу для целей сопоставления введены соот-
ветствующие соотношения, ожидаемые у диплодиплоидных, хотя, по крайней
мере, среди насекомых у таких видов эусоциальность не обнаружена (за исклю-
чением уже отмеченного в разделе 12.1 особого случая с термитами, который
подробнее рассматривается ниже).
r – среднее генетическое сходство (доля идентичных гамет), r (мать – дочери) = (1/2 + 1/2)/2 =
1 /2, r (мать – сыновья) = (1/2 + 1/2)/2 = 1/2, r (мать – дети) = (1/2 + 1/2 + 1/2 + 1/2)/4 = 1/2, r (отец
– дочери) = (1/2 + 1/2)/2 = 1/2, r (отец – сыновья) = 0, r (отец – дети) - (1/2 + 1/2 + 0 + 0)/4 = 1/4,
r (между сестрами) = (1/2 + 1)/2 = 3/4, r (между братьями) = (0 + 1/2)/2 = 1/4, r (сестры – братья) =
(1/2 + 0 + 1/2 + 0)/4 = 1/4, r (между детьми) = 1/2.