Pugnaire F.I. Valladares F. Functional Plant Ecology
Подождите немного. Документ загружается.

Van der Putten, W.H., C. Vandijk, and B.A.M. Peters, 1993. Plant-specific soil-borne diseases contrib-
ute to succession in foredune vegetation. Nature 362: 53–56.
Van der Putten, W.H. and C.D. Van der Stoel, 1998. Plant parasitic nematodes and spatio-temporal
variation in natural vegetation. Applied Soil Ecology 10: 253–262.
van Tol, R., A.T.C. van der Sommen, M.I.C. Boff, J. van Bezooijen, M.W. Sabelis, and P.H. Smits,
2001. Plants protect their roots by alerting the enemies of grubs. Ecology Letters 4: 292–294.
Villenave, C. and P. Cadet, 1998. Interactions of Helicotylenchus dihystera, Pratylenchus pseudopraten-
sis, and Tylenchorhynchus gladiolatus on two plants from the Soudano-Sahelian zone of West
Africa. Nematropica 28: 31–39.
Warrior, P., L. Rehberger, M. Beach, P. Grau, G. Kirfman, and J. Conley, 1999. Commercial devel-
opment and introduction of Ditera TM, a new nematicide. Pesticide Science 55: 343–389.
Weston, L.A. and S.O. Duke, 2003. Weed and crop allelopathy. Critical Reviews in Plant Sciences 22:
367–389.
Whipps, J.M. and K.G. Davies, 2000. Success in biological control of plant pathogens and nematodes
by microorganisms. In: G. Gurr and S. Wratten, eds. Biological Control: Measures of Success,
Kluwer Academic Publishers, pp. 231–269.
Wills, R.T. and J. Kinnear, 1993. Threats to the biota of the Stirling Range. In: C. Thompson, G.P.
Hall, and G.R. Friend, eds. Mountains of Mystery—A Natural History of the Stirling Range.
Department of Conservation and Land Management, Como, Western Australia, pp. 135–141.
Xavier, L.J.C. and J.J. Germida, 2003. Selective interactions between arbuscular mycorrhizal fungi and
Rhizobium leguminosarum bv. viceae enhance pea yield and nutrition. Biology and Fertility of
Soils 37: 261–267.
Yeates, G.W., 1987. Nematode feeding and activity—the importance of development stages. Biology
and Fertility of Soils 3: 143–146.
Yeates, G.W., T. Bongers, R.G.M. Degoede, D.W. Freckman, and S.S. Georgieva, 1993. Feeding habits
in soil nematode families and genera—an outline for soil ecologists. Journal of Nematology 25:
315–331.
Young, I. and K. Ritz, 2005. The habitat of soil microbes. In: R.D. Bardgett, M. Usher, and D. Hopkins,
eds. Biological Diversity and Function in Soils. Cambridge University Press, Cambridge,
pp. 31–43.
Zacheo, G., 1993. Introduction. In: M.W. Khan, ed. Nematode Interactions. Chapman & Hall, London,
pp. 1–25.
Zentmyer, G.A., 1980. Phytophthora cinnamomi and the Diseases It Causes. American Phytopatholo-
gical Society, St Paul, MN.
Francisco Pugnaire/Functional Plant Ecology 7488_C019 Final Proof page 600 16.4.2007 2:38pm Compositor Name: BMani
600 Functional Plant Ecology

20
Resistance to Air Pollutants:
From Cell to Community
Jeremy Barnes, Alan Davison, Luis Balaguer,
and Esteban Manrique-Reol
CONTENTS
Introduction ....................................................................................................................... 601
Cellular Level ..................................................................................................................... 602
Uptake ............................................................................................................................ 603
Metabolism ..................................................................................................................... 607
Gene Expression ............................................................................................................. 609
Plant Level..........................................................................................................................610
Population Level ................................................................................................................ 613
Community Level............................................................................................................... 617
Conclusions ........................................................................................................................ 620
Acknowledgments .............................................................................................................. 620
References .......................................................................................................................... 621
The race is not to the swift, nor the battle to the strong . . . but time and chance happeneth
to them all.
—Ecclesiastes 9:11
INTRODUCTION
The generation of energy by the burning of fossil fuels, all manner of industrial processes, the
biodegradation of wastes, and some farming operations lead to the release of wide range of
contaminants into the air. Most have little or no discernible effect on the environment,
because the resulting concentrations in the atmosphere are well below levels known to be
toxic or because they are not toxic to biological systems. Others attain levels that are known
to threaten human health and to damage both fauna and flora. The situation is not new
because there have been air pollution problems of one kind or another since fire was first used
and metals were first smelted, but the unbridled expansion of industry in many parts of the
world over the past century has resulted in problems on an unprecedented scale, with impacts
extending from the local or regional to global level.
Although air pollution can take various forms (i.e., dusts, smoke, fumes, aerosols, or
mist), this chapter focuses on resistance and adaptation to the most common gaseous
pollutants. Stringent control measures have resulted in a steady decline in the emissions of
several pollutants in developed regions (e.g., sulfur dioxide [SO
2
]); however, ground-level
Francisco Pugnaire/Functional Plant Ecology 7488_C020 Final Proof page 601 25.4.2007 5:25pm Compositor Name: DeShanthi
601

concentrations of some of the most potent gases (e.g., ozone [O
3
]) continue to increase
(Penkett 1988, Boubel et al. 1994, Stockwell et al. 1997). Locally, ground-level concentrations
of some pollutants may be high enough to result in severe foliar injury under conditions
favoring accumulation in the atmosphere (i.e., periods of high solar radiation, favorable
temperatures, or temperature inversions), whereas potentially damaging concentrations of
others (e.g., O
3
) maybe generated at a considerable distance from the source (Bell 1984,
Boubel et al. 1994, Krupa 1996). Long distance transport is usually favored by the high levels
of irradiance and stable atmospheric conditions associated with slow-moving high-pressure
systems in the northern hemisphere. Under such conditions, there is poor dispersal of polluted
air masses, and pollutant concentrations, although typically lower than those experienced
near to point sources, may be high enough to result in subtle changes in plant physiology,
growth, and community composition. Such effects are not necessarily associated with the
appearance of typical visible symptoms of injury, but are more common and just as debili-
tating (Wolfenden and Mansfield 1991, Davison and Barnes 1992, 1998).
It would seem that all plants possess the encoded capability for the perception, signaling,
and response to air pollutants; however, differential expression under the influence of genetic
and environmental factors can result in constitutive and inducible differences between the
reaction norms of plants within and between populations to the same air pollution insult. In
this chapter, we discuss aspects related to the ecotoxicology of airborne pollutants. We begin by
reviewing what is known about the mechanisms underlying differential resistance to the most
common gaseous pollutants, and then attempt to scale-up from responses at the cellular level to
those affecting resistance at the plant, population, and community level. A generic model is
used to provide a conceptual framework within which to discuss the mechanisms underlying
differential resistance. Where appropriate, we have elected to focus on plant responses to O
3
.
Not only because the authors are more familiar with the literature relating to this pollutant
than any other, but also because O
3
is now recognized to be one of the most potent and
widespread toxic agents to which vegetation is exposed in the field (Davison and Barnes
1992, Ka
¨
renlampi and Ska
¨
rby 1996, Fuhrer et al. 1997, Davison and Barnes 1998). Moreover,
increasing concentrations of the pollutants pose a growing threat to vegetation in many regions
(Penkett 1988, Stockwell et al. 1997), and exciting advances have recently been made in our
understanding of the mechanisms underlying the genetic basis of resistance to O
3
(Kangasja
¨
rvi
et al. 1994, Schraudner et al. 1994, Alscher et al. 1997, Pell et al. 1997, Schraudner et al. 1997).
CELLULAR LEVEL
The processes controlling differential resistance to pollutants are considered within the frame-
work of a conceptual model (Figure 20.1), discussed first by Ariens et al. (1976) and later by
Tingey et al. (Tingey and Taylor 1981, Hogsett et al. 1988, Tingey and Andersen 1991), where
resistance* is envisaged to be governed by constitutive and inducible differences in a complex
sequence of events that either reduces pollutant penetration to the target (i.e., avoidance
mechanisms) or enhances the ability of plant tissues to withstand the pollutant and its products
once it has penetrated to the target (i.e., tolerance mechanisms). However, various feedbacks
can also influence plant response, especially in relation to pollutant detoxification and the
repair of injury. These processes are initially dependent on the constitutive resources available,
whereas subsequent responses may be governed by the regulation of gene expression, post-
translational modification of enzymes (e.g., phosphorylation=dephosphorylation), and the
synthesis of secondary defense-related metabolites. If these responses are not sufficient
* Herein defined after Roose et al. (1982) as the ‘‘relative ability of a genotype to maintain normal growth and remain
free from injury in a polluted environment. A trait that is quantitative, rather than qualitative, as resistance need not
be complete.’’
Francisco Pugnaire/Functional Plant Ecology 7488_C020 Final Proof page 602 25.4.2007 5:25pm Compositor Name: DeShanthi
602 Functional Plant Ecology
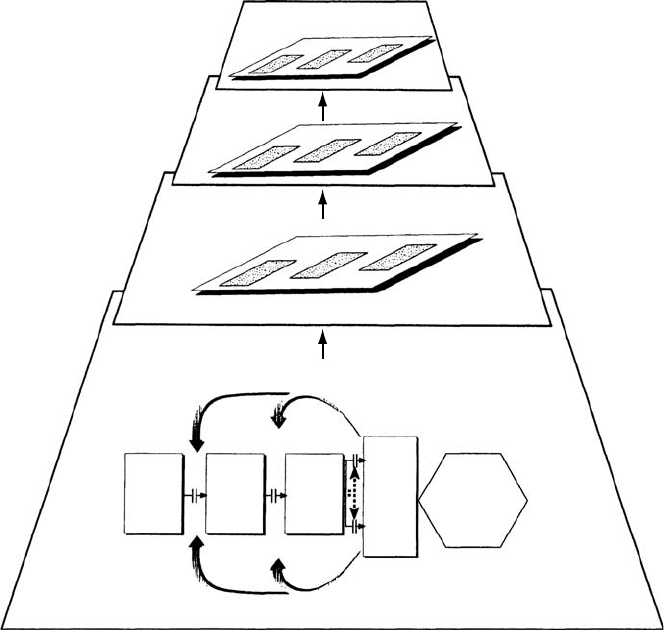
to prevent damage at the cellular level, then there will be destabilization and injury that will be
reflected in downstream consequences at the level of the individual, population, and commu-
nity. The mechanisms conferring resistance may be independent (i.e., pollutant-specific) or
broadly based (i.e., a number of different pollutants trigger the same coordinated defense
reaction); broadly based responses can result in cross-resistance to several pollutants (and
possibly to a number of other environmental stresses), whereas there is growing evidence that
cross-tolerance may be restricted to pollutants (and other stresses) that provoke similar insult
on the same target (Schraudner et al. 1997, Barnes and Wellburn 1998).
UPTAKE
The dose of a pollutant absorbed by plant tissues plays a key role in determining effects on
metabolism and physiology, and in the description and quantification of dose–response
relationships (Taylor et al. 1998, Runeckles 1992). Uptake is predominantly controlled by
rates of foliar gas exchange, with conventional approaches focusing on the importance of the
cuticle and stomata in controlling the rate at which pollutants diffuse into individual
leaves (Mansfield and Freer-Smith 1984). However, it is important to recognize that factors
operating at different scales of resolution influence the rate of uptake, in addition to those
operating at the level of the individual leaf. At a higher scale of resolution (i.e., scaling-up),
Community
Population
Plant
Cell
Destabilization
Complete repair
Modification of gene expression
Detoxification
Ambient
exposure
Internal
exposure
Internal
interaction
New
metabolic
state
Target
exposure/
Reserve
capacity
Uptake Transport/
metabolism
FIGURE 20.1 Conceptual model showing the processes that govern the sensitivity of plants to gaseous
air pollutants. Pollutant levels that exceed the capacity of avoidance=tolerance mechanisms will result in
cellular destabilization—an effect that underpins changes in the performance of the individual and shifts
in the genetic composition of both populations and communities.
Francisco Pugnaire/Functional Plant Ecology 7488_C020 Final Proof page 603 25.4.2007 5:25pm Compositor Name: DeShanthi
Resistance to Air Pollutants: From Cell to Community 603
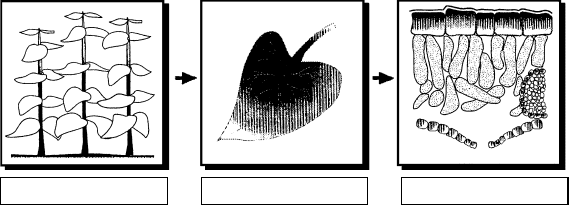
aerodynamic factors influence uptake at the canopy level, whereas at a finer scale of reso-
lution (i.e., scaling-down) physicochemical factors determine uptake at the interface between
the plant and its external surroundings (Figure 20.2).
The size, density, and shape of the canopy all have a pronounced effect on the concen-
tration of a pollutant to which individual leaves are exposed. Relatively few studies have
addressed this issue, but it has been shown that the concentration of O
3
(and other pollutants)
declines as one passes down through the canopy to soil level (Bennett and Hill 1973). As a
result, leaves within a dense canopy tend to be exposed to lower concentrations than those on
the surface or at the edges. However, because the air movement within a dense canopy is
reduced, concentrations within it tend to be less prone to short-term fluctuations (Runeckles
1992). Other features of the leaves (e.g., leaf thickness, Rubisco content, ratio of mesophyll
cell surface area to projected surface area, etc.) and of the environment (e.g., reduced levels of
irradiance, increased temperature, higher humidity, etc.) also differ within the canopy in
relation to those on the outside. This may strongly influence the uptake and effects of
pollutants one leaves at different positions within the canopy and is an important consider-
ation when attempting to extrapolate to the field from laboratory-based studies, identify the
magnitude of the response of different species in a mixture, and establish the impact of
pollutant at different developmental stages, since certain species or particular growth stages
(e.g., seedlings) may be protected from exposure to potentially damaging pollutant concen-
trations by other elements of the canopy.
At the leaf-level, the flux of the pollutant to the leaf interior (J ) is a function of the
concentration gradient between the atmosphere (i.e., the concentration of the pollutant in
the surrounding air, C
a
) and the leaf interior (i.e., the concentration of the pollutant in the
intercellular air spaces, C
i
), and the sum of the physical, chemical, and biological resistances
(SR) to diffusion from source to sink. Mathematically, this is generally expressed in a form
analogous with Ohm’s law, where:
J ¼ (C
a
C
i
)SR:
Plant canopy
Sites of deposition
• Vegetation
• Soil
Sites of deposition
• Substomatal chamber
• Mesophyll tissue
Sites of deposition
• Leaf surface
• Leaf interior
Resistances
• Aerodynamic
• Surface
Resistances
• Cuticular
• Stomatal
Resistances
• Gas-liquid interface
• Residual
Leaf interiorIndividual leaf
FIGURE 20.2 The different scales of resolution that need to be considered in determining rates of
pollutant uptake. (Redrawn from Tanaka, K., Furusawa, I., Kondo, N., and Tanaka, K., J. Plant Cell
Physiol., 29, 743, 1988. With permission.)
Francisco Pugnaire/Functional Plant Ecology 7488_C020 Final Proof page 604 25.4.2007 5:25pm Compositor Name: DeShanthi
604 Functional Plant Ecology
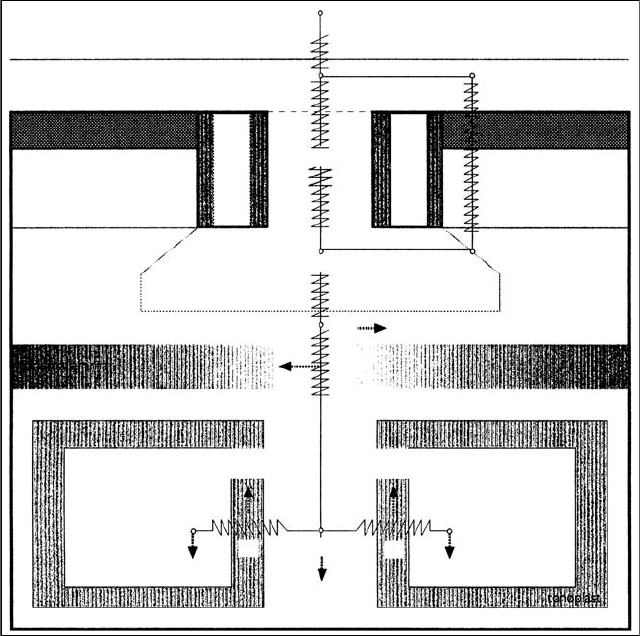
The various impedances (SR) are generally visualized as a network of resistances to gas
flow (Figure 20.3), using Gaastra’s (1959) fundamental principles governing the tortuous
route taken by effluxing water vapor molecules. The most important resistances at the leaf
level are recognized to be those governing the movement of the pollutant into the leaf interior
(i.e., stomatal resistance, r
1
, and cuticular resistance, r
2
), the rate of deposition on the
hydrated surface of mesophyll cells, and the extent of sorption and reaction on the surface
of the cuticle and in the substomatal cavities. These resistances are under genetic and
environmental control, as well as being influenced by the physicochemical characteristics of
the gas in question.
The rate of diffusion of gaseous pollutants through the cuticular membrane is commonly
several orders of magnitude lower than that through the stomata (Lendzian 1984) and may
be considered negligible for some reactive gases, such as O
3
(Kersteins and Lendzian
1989). Hence, the dose of the pollutant absorbed at the leaf level is predominantly con-
trolled by factors determining stomatal conductance (i.e., stomatal aperture and frequency).
Some plants are known to exhibit intrinsically higher stomatal conductance than others
(Ko
¨
rner 1994), and, in general, these tend to be more susceptible to damage (Reich 1987,
Becker et al. 1989, Darrall 1989). However, the response of stomata to external stimuli
(e.g., irradiance, vapor pressure deficit (VPD), soil moisture content, the presence of
C
0
R
0
C
1
C
1
C
2
C
2
R
3
R
2
C
3
C
6
C
4
C
5
R
6
R
5
R
4
R
1
Stoma
Substomatal cavity
Reaction with extracellular antioxidants
Laminar boundary
Air
Epidermis
CuticleCuticle
Epidermis
Cell wall/apoplast
Cytosol
Chloroplast Vacuole
Reaction with
membrane lipids
Reaction with
antioxidants
Reaction with
antioxidants
Reaction with
cytosolic
antioxidants
Reaction with
membrane lipids
reaction with
membrane lipids
reaction with
membrane lipids
Guard cell
Guard cell
FIGURE 20.3 Resistance analog model indicating the network of physical, chemical, and biological
impedances influencing the rate of diffusion of gaseous pollutants from the external atmosphere to
the target.
Francisco Pugnaire/Functional Plant Ecology 7488_C020 Final Proof page 605 25.4.2007 5:25pm Compositor Name: DeShanthi
Resistance to Air Pollutants: From Cell to Community 605
pollutants in the atmosphere, atmospheric CO
2
concentrations, etc.) can strongly influence
the rate of pollutant uptake through effects on stomatal aperture (Mansfield and Freer-Smith
1984, Darrall 1989, Wolfenden and Mansfield 1991, Wolfenden et al. 1992, Heath and Taylor
1997). Differences in stomatal conductance between sensitive and resistant individuals are
rarely sufficient to result in complete exclusion of the pollutant from the leaf interior;
therefore, it is generally concluded that mechanisms resulting in the avoidance of pollutant
uptake are not the only factors determining resistance to airborne pollutants.
Physical leaf characteristics such as leaf thickness, mesophyll cell surface area, internal air
space volume, cell wall thickness, and the volume of the aqueous matrix of the cell wall
influence the eventual concentration of the pollutant and its dissolution products in the
apoplast. Plants show considerable variation in all of these attributes. Mesophyll cell surface
area: projected leaf area, for example, ranges from typical values of between 10 and 40 for
mesophytes, but may be as high as 70 for some xerophytes (Nobel and Walker 1985, Pfanz
1987). There may also be systematic differences in anatomy along altitudinal gradients, which
can contribute to differential resistance (Ko
¨
rner et al. 1989). In addition, pollutant molecules
in the intercellular space or substomatal cavity are partitioned across the gas-to-liquid
interface at a rate determined by the solubility of the gas in the extracellular fluid (Nobel
1974) and its chemical reactivity in the liquid phase (Heath 1988, Heath and Taylor 1997),
factors influenced by temperature and possibly radiation (Barnes et al. 1996, Cape 1997).
Differences in physicochemical properties between the most common gaseous pollutants
result in substantial differences in their solubility in water and rates of diffusion in air
(Nobel 1974), factors that, independent of other considerations, result in substantial differ-
ences in the rate at which individual gaseous pollutants are taken up (Runeckles 1992, Taylor
et al. 1998). Leaf surface characteristics (such as surface wetness, was composition, micro-
morphology, etc.) can also influence the extent of sorption onto foliar surfaces (Wellburn et al.
1997), whereas reactions with other gases in the boundary layer or in the substomatal cavities
may constitute a significant sink for some pollutants, for example, O
3
(Hewitt and Terry 1992,
Salter and Hewitt 1992).
In most instances, difficult and time-consuming measurements of physical leaf character-
istics are not undertaken, so it has become common practice to express rates of pollutant
uptake on the basis of the flux to the leaf interior (i.e., that impinging on the mesophyll cell
surface). This represents what is often termed the absorbed or effective dose of the pollutant,
and can be readily estimated using Fickian diffusion principles from knowledge of boundary
layer, stomatal and cuticular conductances, correcting for differences in the diffusivities
between water vapor and the pollutant of interest. Hence, the flux of O
3
to the leaf interior
(J
O
3
) may be described as
J
O
3
¼ g
b
þ 0:612 g
H
2
O
(O
a
O
i
),
where g
b
is the turbulent boundary layer conductance, 0.612 is the difference in the binary
diffusivities of water vapor and ozone in air (Nobel 1983), g
H
2
O
represents the stomatal
conductance to water vapor, and O
a
and O
i
represent the concentrations of O
3
in the external
atmosphere and in the intercellular spaces, respectively. No correction needs to be made for
uptake through the cuticle in this case, since the cuticle is considered to represent a virtually
impermeable barrier to O
3
(Kersteins and Lendzian 1989), whereas measurements of the
intercellular O
3
concentration (O
i
) suggest it is close to zero (Laisk et al. 1989). On a
cautionary note, it is important to emphasize two points. First, such calculations represent
the gross flux to the leaf interior—for some gases where the plant may act as a source as well
as a sink (e.g., H
2
S, NH
3
NO), correction for effluxing gas molecules is required to enable
estimates of the net flux of the pollutant. Second, fluxes determined in the above manner take
no account of differences in physical leaf characteristics or internal resistances influencing the
Francisco Pugnaire/Functional Plant Ecology 7488_C020 Final Proof page 606 25.4.2007 5:25pm Compositor Name: DeShanthi
606 Functional Plant Ecology
rate at which the pollutant (or its products) is delivered to the eventual target. Potentially
more informative models are available that enable estimates of the extent of penetration to
the plasmalemma and beyond (Chameides 1989, Ramge et al. 1992, Plo
¨
chl et al. 1993), but
these have rarely been used due to the uncertainties surrounding the complex solution
chemistry of certain gases (e.g., O
3
) and the relative importance of scavenging=transformation
mation in the cell wall region.
METABOLISM
Once the pollutant has penetrated as far as the mesophyll cell surface, it may be metabolized,
sequestered, or excreted. Recent attention has focused on the rate at which pollutants (and
their reactive products, including a variety of free radical species) are immobilized and
detoxified at either the first barrier encountered after entry into the leaf (i.e., in the apoplast)
or subsequently, after penetration into the cell proper. In any consideration of the processes
underlying pollutant detoxification, it is important to recognize that dissolution in the
apoplast can result in different types of stress on cellular constituents. Many gases (e.g.,
HF, NO
x
, NH
3
, and SO
2
) induce acidification of subcellular compartments, whereas others
(e.g., O
3
, PAN, NO
x
) result in oxidative stress, and some (e.g., SO
2
) produce both (Malhotra
and Khan 1984, Hippeli and Elstner 1996, Mudd 1996). The extent of damage resulting
from the former is related to the ability of cells to buffer the increase in acidity or to excrete
protons to the external media (Slovik 1996, Burkhardt and Drechsel 1997), whereas the
influence of the latter is dependent on the efficiency of endogenous antioxidant systems
that scavenge free radical and active oxygen species before they can react with cellular
constituents (Kangasja
¨
rvi et al. 1994, Luwe and Heber 1995, Alscher et al. 1997).
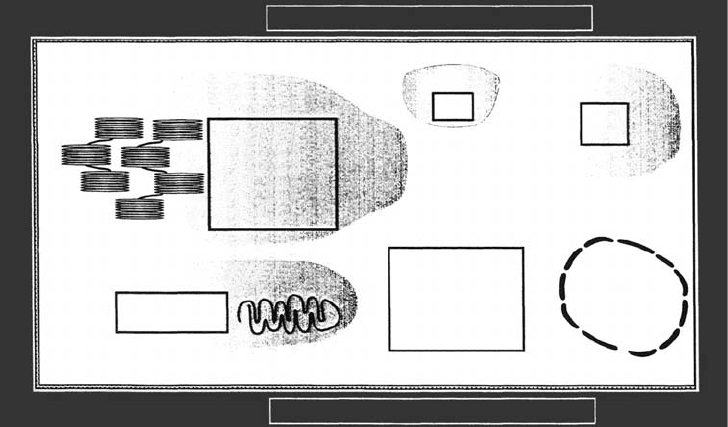
Detoxification systems capable of protecting sensitive targets from the oxidative stress
imposed by pollutants and their derivatives are common in plants, as in animals, and are
subject to strict genetic control. Most attention has focused on those systems located in the
various intracellular compartments (in chloroplasts, mitochondria, cytosol, and peroxi-
somes); however, similar systems are intimately associated with the plasmalemma and the
cell wall (Figure 20.4). In recent years, the latter has attracted increased attention since there
is growing evidence that some pollutants (e.g., O
3
, SO
2
, NO
2
) and their dissolution products
may be scavenged and detoxified=transformed at the mesophyll cell surface (i.e., in the
apoplast). The aqueous matrix of the cell wall is now recognized to contain significant
quantities of ascorbic acid (vitamin C) and polyamines as well as isoforms of Cu=Zn super-
oxide dismutase (SOD), ascorbate peroxidase (APX), and nonspecific peroxidases (GPODs)
(Polle and Rennenberg 1993, Luwe and Heber 1995, Ogawa et al. 1996, Dietz 1996), which
are known to function as antioxidants (Polle and Rennenberg 1993, Kangasja
¨
rvi et al. 1994,
Alscher et al. 1997, Polle 1997). Research is still at an early stage and many questions remain
to be answered, but preliminary model calculations based on the scavenging of O
3
(rather
than its dissolution products) by apoplastic ascorbic acid indicate that detoxification pro-
cesses operating in the apoplast may be sufficient to provide at least limited protection against
O
3
(Chameides 1989, Polle and Rennenberg 1993, Lyons et al. 1999), a finding supported by
several independent lines of evidence (Lyons et al. 1998). Although the relative importance of
scavenging in the cell wall region remains to be established, there is a growing opinion that
factors such as the extracellular concentration of ascorbic acid may play a central role in
determining resistance to O
3
(Conklin et al. 1996, Lyons et al. 1998), as well as influencing the
impacts of SO
2
(Dietz 1996) and NO
2
(Ramge et al. 1992).
Pollutants and their reactive products that breach the extracellular defense, or are pro-
duced from the reaction with plasmalemma constituents, must be scavenged by intracellular
detoxification systems if damage is to be averted. There is, for example, strong evidence
linking SO
2
tolerance with the activity of intracellular enzymes such as catalase (CAT),
Francisco Pugnaire/Functional Plant Ecology 7488_C020 Final Proof page 607 25.4.2007 5:25pm Compositor Name: DeShanthi
Resistance to Air Pollutants: From Cell to Community 607
superoxide dismutase (SOD), glutathione synthetase (GS), glutathione reductase (GR),
glutathione peroxidase (GPX), and glutathione transferase (GST) (Tanaka et al. 1988, Ranieri
et al. 1992, Lea et al. 1998), as well as with levels of glutathione (GSH) and ascorbic acid (ASC)
(Madamanchi and Alscher 1994). Furthermore, recent work has drawn attention to the
importance of the subcellular localization of these systems in relation to the protection afforded
against different pollutants. For example, SO
2
tolerance in transgenic plants engineered to
overexpress GR in different cellular compartments indicates that it is the cytoplasmic activity
of this enzyme, rather than that of the plastidic forms, which is important in determining SO
2
tolerance (Aono et al. 1993, Broadbent 1995, Lea et al. 1998). There is also considerable
evidence linking components of the cellular antioxidant system (ASC, GSH, polyamines,
a-tocopherol, carotenoids, SOD, ascorbate peroxidase [APX], GR, and CAT) with O
3
toler-
ance (Heath 1988, Kangasja
¨
rvi et al. 1994, Hippeli and Elstner 1996, Mudd 1996, Alscher et al.
1997, Pell et al. 1997, Heath and Taylor 1997). However, the degree of protection afforded by
these intracellular systems in relation to that achieved by scavenging in the cell wall region
(the primary site of O
3
action) remains poorly understood (Lyons et al. 1998), and it is
interesting to note that transgenic plants overproducing particular antioxidant enzymes in
different intracellular compartments rarely display enhanced O
3
resistance (Pitcher et al. 1991,
Van Camp et al. 1994, Pitcher and Zilinskas 1996, Torsethaugen et al. 1997).
In addition, toxicity thresholds are influenced by the efficiency of metabolic processes
resulting in the utilization, sequestration, and excretion of pollutants. The plasmalemma, for
example, is known to constitute an important obstacle impeding the penetration of pollutants
(and their products) to their intracellular sites of action (Herschbach et al. 1995). Some
pollutants (e.g., O
3
) react readily with membrane constituents (Mudd 1996, Heath and Taylor
1997). The uptake of others (i.e., SO
2
-derived sulfate [SO
2
4
], sulfite [SO
2
3
], and bisulfite
[HSO
3
] is limited by the activity of membrane-bound carriers and transformation processes
Plasma membrane
α-Tocopherol
Chloroplasts
Mitochondria
GSH
SOD
MDHAR
Plasma membrane α-Tocopherol
GR
APX
α-Tocopherol
AA
MDHAR & DHAR
GSH
Carotenoids
SOD
GR
APX
PODs
AA
GSH
MDHAR & DHAR
SOD
GR
Peroxisomes
Cytosol
Vacuole
Nucleus
SOD
CAT
GSH
AA
PODs
Extracellular fluid
Extracellular fluid
AA, APX, PODs, SOD, Polyamines, (GSH?)
AA, APX, PODs, SOD, Polyamines, (GSH?)
FIGURE 20.4 Subcellular localization of the antioxidant systems. AA, ascorbic acid; GSH, glutathione;
APX, ascorbate peroxidase EC. 1.11.1.11; MDHAR, monodehydroascorbate radical reductase EC.
1.1.5.4; DHAR, dehydroascorbate radical reductase EC. 1.8.5.1; GR, glutathione reductase
EC 1.6.4.2; SOD, superoxide dismutase EC. 1.15.1.1; CAT, catalase EC. 1.11.1.6; PODs, nonspecific
peroxidase (sometimes referred to as guaiacol peroxidase) EC. 1.11.1.7.
Francisco Pugnaire/Functional Plant Ecology 7488_C020 Final Proof page 608 25.4.2007 5:25pm Compositor Name: DeShanthi
608 Functional Plant Ecology
(Pfanz et al. 1990). Indeed, research on the inheritance of SO
2
resistance in Cucumis sativus L.
suggests that differences in intrinsic membrane properties may contribute to variations in SO
2
resistance (Bressan et al. 1981). Once inside the cell, the products of some (e.g., SO
2
4
, NO
3
,
and NH
þ
4
) may be metabolized via the usual channels, sequestered for later use, stored
indefinitely, or volatilized. For example, SO
2
-derived SO
2
4
can either be metabolized to
yield elevated levels of water-soluble nonprotein thiols (such as cysteine, g-glutamylcysteine,
and glutathione), which can be degraded at a later date to provide reduced S to support
new growth (De Kok 1990), or it can be sequestered on a semipermanent basis in the vacuole
(presuming there is sufficient available energy and H
þ
ions to facilitate its transport across
the tonoplast) (Cram 1990, Kaiser et al. 1989, Slovik 1996). In contrast, SO
3
may be
photoreduced and, after volatilization, be reemitted, mainly as H
2
S. This pathway was
originally considered to form a possible pathway for the detoxification of SO
2
(Rennenberg
1984), but current opinion suggests that the contribution of such emissions to the detoxifi-
cation of environmentally relevant SO
2
concentrations may be negligible (Stuhlen and
De Kok 1990).
GENE EXPRESSION
The photoautotrophic habit adopted by plants has resulted in the evolution of a sophisticated
battery of mechanisms that renders them, with exception of all but a few microbes, the most
adaptable of all multicellular organisms on the planet (Smith 1990). This flexibility includes
the capacity to sense and react to the presence of airborne pollutants, as to other environ-
mental stimuli, in a manner directed at sustaining survival to reproduction, a goal that may or
may not be achieved, depending on the extent of metabolic flexibility and the degree of stress
imposed at the cellular level.
Exposure to pollutants and other oxidative stresses induces changes in the expression
of defense-related genes, posttranslational modification of enzymes (e.g., phosphoryla-
tion=dephosphorylation), and the synthesis of secondary metabolites—resulting in increases
in the threshold for damage, that is, acclimation (Kangasja
¨
rvi et al. 1994, Schraudner et al.
1994). Recent work suggests that the pattern of changes induced by some pollutants (e.g., O
3
)
reflects on orchestrated series of events, triggered by disparate oxidative syndromes, which
resembles the hypersensitive response provoked by pathogen attack (Alscher et al. 1997,
Pell et al. 1997, Schraudner et al. 1997). This raises the question of whether the patterns of
defense-related gene expression triggered by one pollutant are similar to those induced by
another, whether the same signal transduction pathways are involved, and whether the
similarity in response results in enhanced tolerance to a range of pollutants, and possibly
other oxidative stresses. Opinions differ. However, based on the fact that particular geno-
types, specific transformants, and plants treated with specific protectants (e.g., EDU) may be
sensitive to one pollutant but not to another (Barnes and Wellbum 1998 and references
therein), whereas the direction and the extent of the responses of the same species to different
pollutant combinations vary in individual genotypes grown under common conditions
(Bender and Weigel 1992), we take the view that despite the common responses observed at
the level of gene expression, specific mechanisms must underlie tolerance to different air
pollutants (i.e., tolerance to one pollutant is independent of that to another), as the action and
subcellular localization of the stresses imposed by pollutants differ. This conclusion is
supported by observations that differences in stomatal behavior=conductance (i.e., avoidance
mechanisms) commonly govern the similarity in response to different pollutants (Winner et al.
1991). It is also interesting to note that some pollutants (e.g., SO
2
and NO
2
) are much less
effective than others (e.g., O
3
) in eliciting changes in antioxidant gene expression (Schraudner
et al. 1994, Schraudner et al. 1997), whereas other stresses (such as wounding, necrotizing
pathogens, or elevated levels of UV-B radiation), which are known to elicit strong and rapid
Francisco Pugnaire/Functional Plant Ecology 7488_C020 Final Proof page 609 25.4.2007 5:25pm Compositor Name: DeShanthi
Resistance to Air Pollutants: From Cell to Community 609