Нестурх М.Ф. Происхождение человека
Подождите немного. Документ загружается.


перебирая прядь за прядью, обезьяна время от времени схватывает пальцами и ногтями
насекомое или иной посторонний предмет, находящийся в шерсти или на коже.
Насекомые бывают весьма мелки и назойливы, избавление от их присутствия на теле составляет
благодеяние для тропических животных и для людей. Достаточно вспомнить приводимые
разными путешественниками описания их длительных и нередко мучительных странствований
по Африке, где одним из главнейших препятствий были мириады всевозможных мелких
жалящих и колющих насекомых, в том числе муравьев. Увидеть и поймать мелкое насекомое,
спасающееся быстрым бегством или ловкими прыжками, не так просто. Это требует острого
зрения, большой быстроты и координации движений, хорошо налаженной связи в мозгу между
двигательной зоной передней конечности и зрительной зоной.
Нужно отметить, что термин «искание блох» мало отражает действительное содержание поисков
обезьян в шерсти друг у друга. Возможно предполагать, что здесь. большую роль играет
удаление слабых или уже выпавших волос, от которых тотчас же поедаются обезьяной луковица
и основание волоса (вместе с находящимися здесь мельчайшими кристалликами веществ,
входящих в состав пота).
Бинокулярное зрение прогрессировало у предков человека — обезьян — параллельно с
развитием более тонких хватательных движений передней конечности и с усовершенствованием
способности большого пальца кисти противопоставляться остальным пальцам.
Манипулирование различными съедобными предметами, поворачивание их перед глазами и
рассматривание с разных сторон, откусывание частей плода и рассматривание надкусанных
частей, поимка и рассматривание мелких паразитов — все подобные действия должны были
содействовать в процессе естественного отбора комбинированному прогрессивному развитию
двигательного анализатора, передней конечности и сочетанию движений с контролем зрения
еще у обезьян третичного периода. Трудовые действия древнейших людей преобразовывали
216
этот процесс и усилили его уже в первые сотни тысяч лет четвертичного периода, о чем в
известной мере можно судить по слепкам мозговой полости черепа, или эндокранам.
Приготовление орудий немыслимо без особо развитой хватательной передней конечности —
руки, без острого бинокулярного зрения. А такое сочетание анатомофизиологических
особенностей как раз и имелось у наших предков.
Следовательно, для возникновения человека и всей его дальнейшей эволюции было необходимо
непрерывное прогрессивное развитие общего строения нервно-мышечного аппарата руки,
тонкой архитектоники соответствующих частей центральной нервной системы и зрительного
аппарата. Взаимно влияя друг на друга, рука и мозг развивались в процессе общественного
труда.
Осязательный и зрительный анализаторы в соединении с двигательным развивались у древних
приматов в сравнении с их наземными предками по-новому, в ином качественном состоянии,
позволяя приспособляться не только к смене типа локомоции и способа питания, но и к
условиям межвидовой борьбы, в частности с такими опасными животными, как средние и
крупные кошачьи хищники, разные змеи или налетающие с воздуха хищные птицы.
Немалое значение приобретает слуховой анализатор, специализирующийся в различении
разнообразных лесных звуков, имеющих жизненное значение. Чуткость органов слуха очень
важна для обезьян не только днем, но и ночью: она нередко спасает их от подкрадывающихся
ночных хищников. Обезьяны постоянно прислушиваются к звукам голоса, издаваемым другими
особями стада. Некоторые звуки обезьян имеют особое значение сигналов, причем довольно
безразлично, какая именно из них издаст звук, предупреждающий об опасности.
Большое исследование средств общения у обезьян в стаде осуществила Н. А. Тих на Сухумской
медико-биологической станции Академии медицинских наук СССР (Тих, 1950). Работа была
произведена на павианах гамадрилах, у которых оказалось около пятнадцати разнообразных
звуков голоса, выражающих эмоциональное состояние обезьян. Тих установила, что звуки,
издаваемые обезьянами, тесно связаны с эмоциональной основой и соответствующими
телодвижениями. Их искусственное вызывание даже по методу условных, рефлексов
натолкнулось на большие трудности, в частности вследствие стереотипности звукового
выражения эмоций, связанности, врожденных (безусловных) звуков с определенной
биологической значимостью. Так, звук оборонительного значения гораздо труднее сделать
пищевым сигналом, чем звук на пищу. В опытах М. А. Панкратова при этом наблюдался даже
срыв высшей нервной деятельности у
обезьян (Воронин, 1952).
Сюда присоединяется то
обстоятельство, что периферический и
корковый отделы слухового анализатора
у обезьян, в том числе и
человекообразных, частью выше
дифференцированы, чем у человека.
Например, обезьяны слышат такие
высокие тона (около 30 тыс. колебаний в
секунду), какие недо-
217
ступны человеческому невооруженному
уху и обнаруживаются только с
помощью специальных акустических
аппаратов. Обезьяны обладают
развитыми ушными мышцами и
свободно двигают сводами ушными
раковинами, в то время как наружное
ухо человека при его больших размерах
лишено или почти лишено
подвижности, как у гориллы и
орангутана, обладающих маленькими
ушами.
Точно так же в некоторых отношениях у
обезьян сильнее развиты такие
анализаторы, как, например, глазной,
уже не говоря об обонятельном.
Другими словами, в области первой
сигнальной системы обезьяны даже
обладают известным преимуществом.
Зато человек несравненно превосходит
их ближайших сородичей в
деятельности головного мозга,
свойственной его анализаторам в связи с
работой второй сигнальной системы и
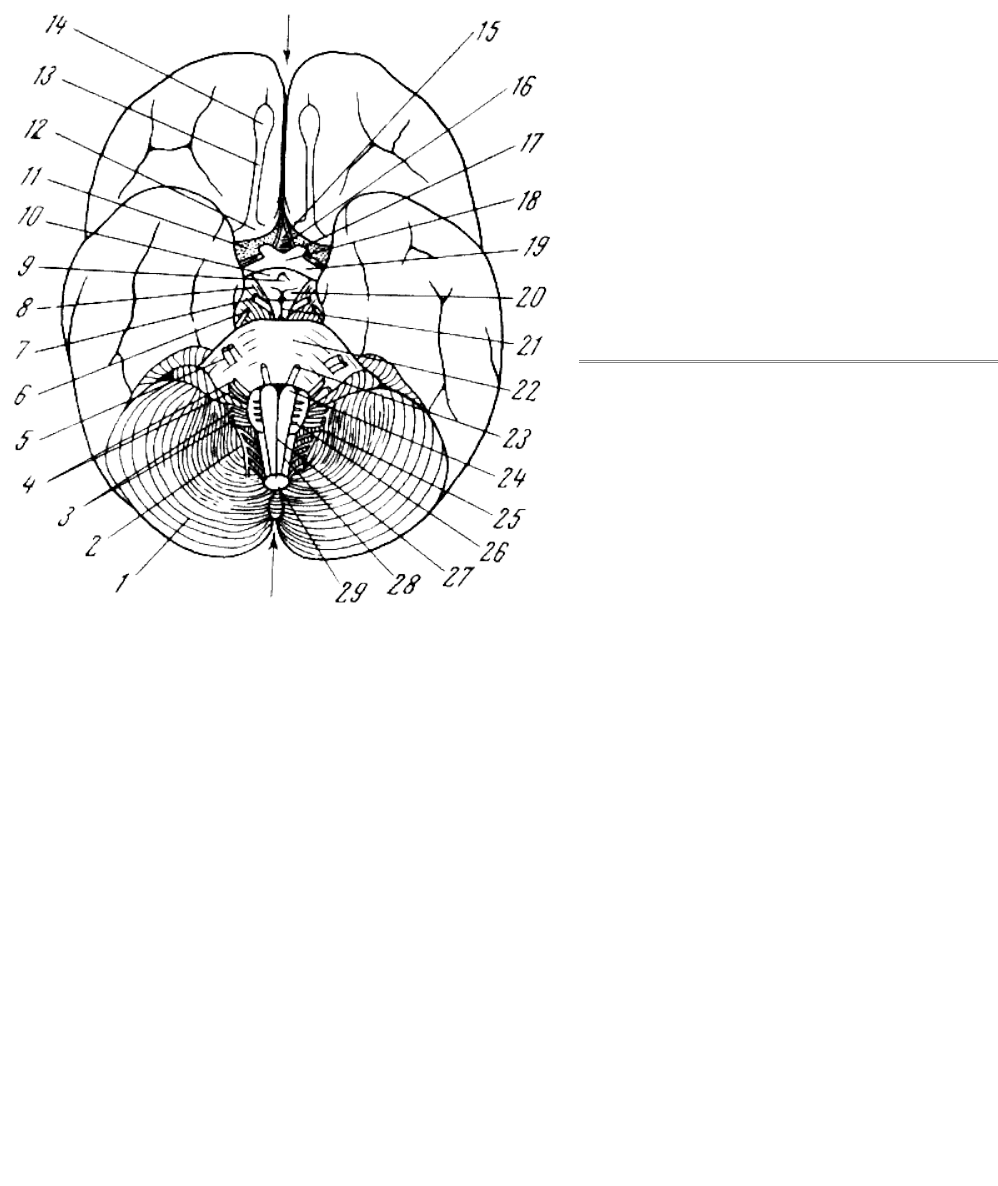
Рис. 67. Головной мозг человека (вид снизу)
1 — мозжечок; 2 — добавочный нерв; 3 —
языкоглоточный и блуждающий нервы; 4 — лицевой
и слуховой нервы; 5 — тройничный нерв; 6 —
блоковый нерв; 7 — глазодвигательный нерв; 8 —
ножки мозга; 9 — воронка; 10 — серый бугор; 11 —
серое переднее продырявленное поле; 12 —
обонятельный треугольник; 13 — обонятельный
тракт; 14 — обонятельная луковица (стрелка между
14 и 15 указывает на продольную фиссуру головного
мозга); 15 — конечная пластинка; 16 — диагональная
связка; 17 — глазной нерв; 18 — зрительный
перекрест; 19 — зрительный тракт; 20 — мозговые
шарики; 21 — ямка Тарини между ножками моста;
22 — мост и основная борозда моста; 23 —
отводящий нерв; 24 — пирамида продолговатого
мозга; 25 — олива; 26 — подъязычный нерв; 27 —
срединная передняя фиссура продолговатого мозга;
28 — передняя боковая борозда продолговатого
мозга; 29 — продолговатый мозг (в разрезе: стрелка
между 29 и 1 — указывает на заднюю вырезку
мозжечка). По Э. Виллигеру, 1930

обеспечивающей познание признаков вещей, недоступное никакому животному.
Периферические отделы слухового, осязательного, зрительного и других анализаторов в ходе
эволюции приматов под влиянием внешней среды находились в неразрывной связи с корковыми
отделами больших полушарий мозга и развивались вместе с ними. Перейдем к изложению
некоторых необходимых данных, касающихся соответствующих областей коры.
Тип мозга обезьяны развился из типа мозга более примитивного примата вроде полуобезьяны.
Так, у лемура мозг невелик и весит не более нескольких десятков граммов; его обонятельные
части развиты довольно значительно, а лобные доли слабо; височная, характерная для приматов
вообще, явно очерчена; поверхность полушарий у большинства приматов с небольшим
количеством извилин; на затылочной доле заметна шпорная борозда, имеющаяся и у всех других
приматов; малый мозг, или
218
мозжечок, почти не прикрыт большим мозгом.
В стволовой части головного мозга всех приматов, в том числе и у человека, подмечаются черты
сегментированности, более заметно обнаруживающиеся в спинном мозгу. У последнего это ярко
проглядывает в попарном равномерном отхождении пучков спинных чувствительных и
брюшных двигательных нервов от спинного мозга, а у первого — в симметричном порядке
расположения двенадцати пар черепных нервов. Подобная сегментированность свидетельствует
о том, что не очень далекими предками человека были четвероногие животные и что более
отдаленными звеньями человеческой родословной служили низшие позвоночные. Вместе с
полуобезьянами и обезьянами человек уходит своими корнями в глубину животного мира. Его
головной мозг имеет в основе исходный обезьяний тип строения.
Мозг низшей обезьяны вроде макака или павиана крупнее и тяжелее, чем у лемура, достигая
веса 50—100 г; обонятельные луковицы невелики, а в большом мозгу извилины и другие
участки древнего обонятельного отдела коры мало развиты, хотя и сильнее, чем у человека (рис.
67).
В коре человеческого мозга, по данным исследования И. Н. Филимонова (Институт мозга,
Москва), основная зона, связанная с обонятельной функцией и обозначаемая в качестве древней
коры, или палеокортекса, составляет лишь 0,61% площади поверхности коры (Филимонов,
1949). Если говорить о количественном уменьшении обонятельной области в ряду приматов, то
оказывается, что, по данным того же автора, «величина отношения древней коры ко всей коре
уменьшается у человека в сравнении с игрункой очень резко (в 4,7 раза)».
В противоположность обонятельной области, или древней коре, большого мозга, его новая кора,
или неокортекс, представляет собой в ряду приматов прогрессивную часть мозга и достигает у
человека огромного развития (рис. 68—69). Это происходит, в частности, благодаря разрастанию
некоторых долей коры полушария специфическим для человека образом. В качестве примера
остановим внимание на нижнетеменной области неокортекса обезьян и человека.
Ю. Г. Шевченко, изучив в Институте мозга нижнетеменную (нижнепариетальную) область коры
у игрунки, мартышки, гиббона, шимпанзе, орангутана и человека, установила сходство в чертах
основной структуры и обнаружила у антропоидов поля 39 и 40, характерные для человека
(Brodmann, 1925). Одновременно она обнаружила и заметные количественные и качественные
отличия. С одной стороны, границы между соседними полями и подполями данной области у
антропоидов более резкие, чем между соответствующими формациями у человека, с другой
стороны, у антропоидов они слабее развиты, не захватывая всей нижнепариетальной дольки, а
лишь ее верхние отделы. У игрунки нижнетеменная область равна 0,17% от всей поверхности
коры, у орангутана и шимпанзе составляет лишь 3,3%, в то время как у человека она очень

219
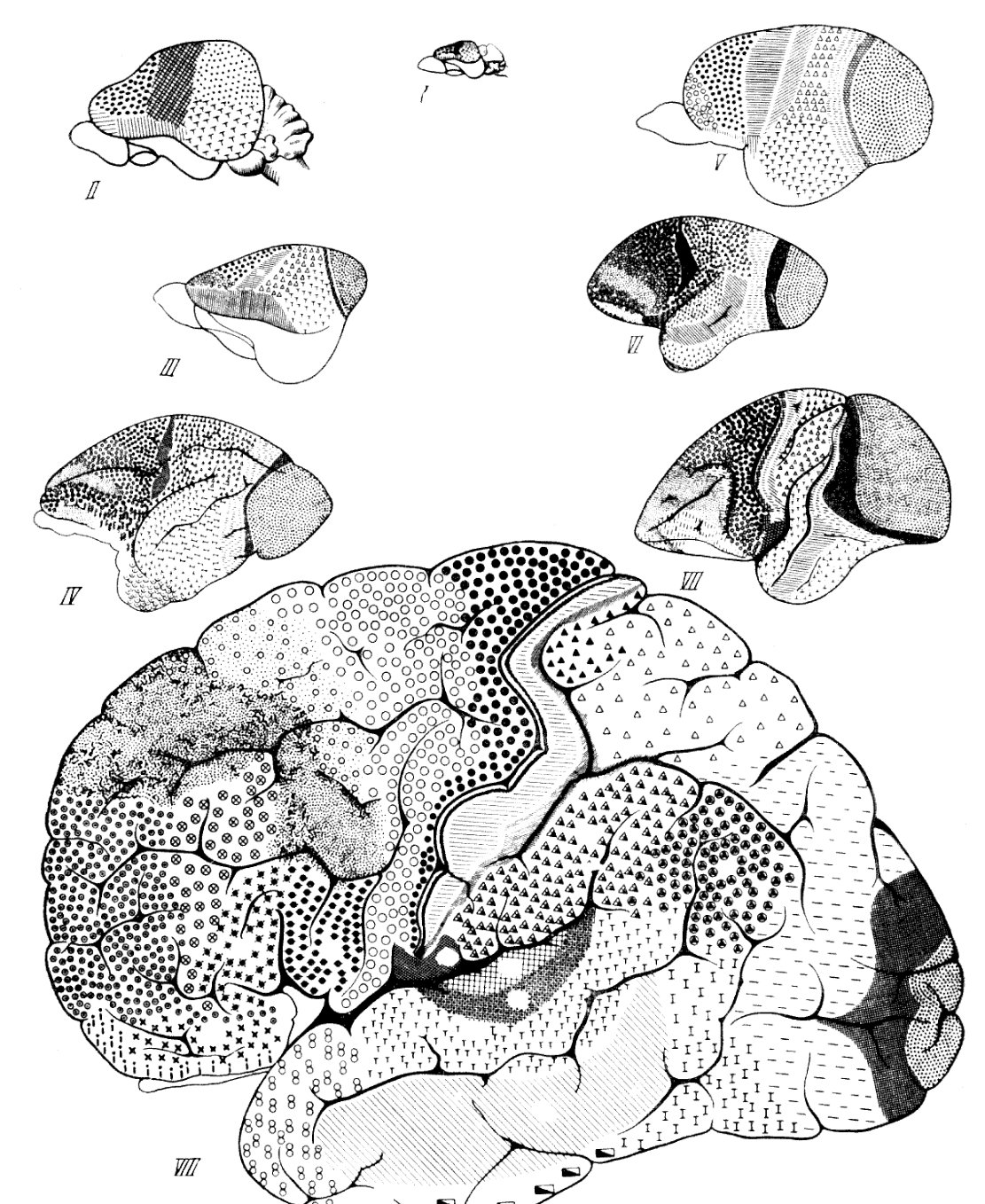
Рис. 68. Цитоархитектонические поля коры больших полушарий мозга приматов
I — насекомоядное млекопитающее хоботный прыгунчик (Macroscelides elephantulus); II — тупайя
собственно (Tupaia sp.); III — тупайя перохвостая (Ptilocercus lowii); IV — лемур (Lemur sp.); V —
долгопят (Tarsius spectrum); VI — игрунка (Hapale jacchus); VII — макак (Macacus sp.); VIII — человек
(Homo sapiens); I, II, II, V — по Ф. Вуду Джонсу, 1929; IV, V, VI и VII — по К. Бродманну, 1925
Рис. 69. Области коры больших полушарий головного мозга
1 — оранг; 2 — шимпанзе; 3 — горилла, 4 — человек; 5 — лемур; 6 — долгопят; 7 — капуцин; 8 —
макак; 9 — павиан; 10 — гиббон; а — двигательная область, б — чувствительная (сенсорная); в —
зрительная; г — слуховая. Вид слева. Лобная доля не заштрихована (на рисунках видно ее
прогрессивное развитие от лемура и долгопята до антропоидов и человека). Точками на лобной доле у
человека обозначена двигательная зона речи. 2/7 нат. вел. По Ф. Тильнею, 1928 (ширина изображения
117 мм)
разрослась и охватывает 7,7% (Шевченко, 1936—1938).
Приведенные соотношения тем более представляются наглядными, что, хотя по размерам
площади поверхность коры полушарий большого мозга человека в три раза превосходит ее у
шимпанзе, вся нижнепариетальная область у человека в десять раз больше, чем у
человекообразных обезьян. Такое сильное развитие филогенетически новой нижнетеменной
области у человека, очевидно, зависело от ее более усиленного и своеобразного

функционирования под влиянием трудовых действий, звукового языка, второй сигнальной
системы. Нарушения деятельности этой области обнаруживаются в сложных нервных
расстройствах.
Таким образом, нижнетеменная область тесно связана с функциями по линии второй сигнальной
системы — со звуковой речью, чтением, письмом: филогенетически молодая область коры у
человека качественно отличает его мозг от обезьяньего.
Лобная область также включает весьма существенные филогенетические новые образования,
высоко ставящие мозг человека над животным. В Институте мозга (Москва) ее подробно
исследовала
221
Е. П. Кононова, которая сделала очень важное открытие, когда обнаружила в коре нижней
лобной доли мозга шимпанзе участки, соответствующие 44 и 45 полям полушарий
человеческого мозга, между тем как зарубежные ученые отрицали наличие их гомологов даже у
антропоидов. Кроме того, ей удалось найти следы или зачатки подобных участков даже у
некоторых низших обезьян. Почему так важны поля 44 и 45 для человека? Уже давно ученые
подметили связь этой структуры с функцией членораздельной речи. Соответствующий участок
даже называли по имени крупного французского антрополога «центром речи Брока». Но позже
было установлено, что к речевой функции имеет ближайшее отношение не только лобная
область, но также нижнетеменная и височная. Исследования показали, что в нижней лобной
извилине помещается двигательная зона речевой функции. При ее патологических состояниях в
левом полушарии у правшей возникает двигательная афазия и иные расстройства речи
(Пенфилд, Роберте, 1964).
О том, что развитие лобной доли человека носило прогрессивный характер, свидетельствуют и
другие ее анатомо-физиологические особенности, например очень многочисленные
ассоциативные связи с помощью пучков волокон с другими отделами мозга. Это подтверждают
факты поздней миэлинизации волокон (как и в нижней теменной доле), сравнительного
увеличения лобной доли в ходе эволюции, а также заметного усложнения узора борозд всех
размеров в связи с обогащением количества извилин на поверхности полушарий большого
мозга.
Следующей областью, связанной с речевой функцией, является височная: в ней находится зона
восприятия звуков, в частности членораздельной речи или, шире, звукового языка. Работа С. М.
Блинкова (1955) показывает, что в поле 41, которое здесь имеет особое значение в качестве так
называемой «слуховой зоны», кора более сложно дифференцирована, чем, у обезьян. То же
следует сказать и о филогенетически новой территории заднего участка височной доли. Еще
более резкие различия от мозга обезьяны находятся в тончайшей структуре поля 21 средней,
подобласти височной доли человеческого мозга.
О важности определенных территорий височной доли, в частности,: участка, носящего название
«зоны Вернике» (по имени немецкого невролога), свидетельствует их патологическое состояние,
приводящее к непониманию речи, т. е. к сензорной афазии. Некоторые заболевания в соседней
доле, затылочной. ведут к словесной слепоте типа оптической алексии, когда человек перестает
узнавать те или иные буквы и целые слова. Таким образом, и эта доля, вероятно, принимает
значительное участие в осуществлении функций специфической только для человека второй
сигнальной системы, так или иначе связанной со всей корой.
Затылочная область коры, в которой располагается мозговой конец зрительного анализатора, как
раз и служит у человека главнейшим органом восприятия зрительных образов в связи с первой и
второй сигнальными системами.

222
Неудивительно,что и тут исследователи устанавливают заметные отличия по сравнению с
мозгом других приматов, можно сказать даже обнаруживают качественную перестройку в связи
с общим процессом преобразования мозга обезьяны в мозг человека.
Воспользуемся в данном случае материалами исследования, сделанного в Институте мозга на
полушариях большого мозга человека, высших и низших обезьян (Преображенская, Филимонов,
1949).
Установлено, что у человека из трех полей затылочной области наибольшего развития и
сложности строения достигло поле 19, в то время как поле 17 невелико. И если у человека поле
19 составляет 4,5% всей поверхности коры, у орангутана — 6,4% и у низшей,
мартышкообразной обезьяны — 6,0%, то поле 17 соответственно занимает 3,0, 8,5 и 10,0%.
Отсюда видно, что и в затылочной доле большого мозга человека имеются такие черты, которые
придают ему значительное своеобразие. Другими словами, развитие полей 17, 19 и всей
затылочной области у человека пошло совсем в другом направлении, чем у его ближайших
сородичей — обезьян.
Иной представляется и картина расположения полей на полушарии: так, поле 17 у человека
располагается не на боковой, или латеральной, его стороне, а почти целиком на серединной, или
медиальной, вследствие разрастания теменно-височных частей коры. Туда же переместилась и
шпорная борозда, которая у обезьян находится частично на латеральной стороне полушария у
самого затылочного полюса. У низших обезьян зрительно-окципитальная кора надвигалась на
теменную и частично перекрыла ее заднюю грань. В связи с этим здесь и возникла обезьянья
борозда, ярко обозначающаяся на латеральной поверхности больших полушарий в пограничном
районе теменной и затылочной долей.
У человека затылочная доля испытала сильное потеснение и некоторое уменьшение
относительных размеров, что зависит от разрастания нижнетеменной, височной и лобной долей.
Следовательно, в этом случае прогрессивное развитие одних отделов коры большого мозга
вызывало относительный регресс других ее частей или элементов, что справедливо и для всего
мозга в целом: специфически человеческие черты в процессе антропогенеза появились на базе
прежних и совсем заново, обезьяньи же ослабевали и исчезали. Проиллюстрируем сказанное
еще одним примером.
На границе лобной и височной долей, в глубине сильвиевой борозды, располагается центральная
(стволовая) доля, или так называемый рейлиев островок (рис. 70). Он обнаруживается при
раздвигании краев борозды, образованных покрышками, или оперкулярными частями, соседних
долей, в том числе теменной. Иная картина у обезьян, так как у них центральная, или
инсулярная, доля: частично выдается на поверхность, располагаясь между названными выше и
окружающими долями и занимая срединное положение (Дешин, 1934).
Говоря о мозге предшественников древнейшего человека,
223

Ю. Г. Шевченко (1967) считает, что он
развивался под влиянием прогресса
передних конечностей в сторону
прогрессивного конструктивного
манипулирования предметами при
одновременном совершенствовании
двуногого хождения, о чем может
свидетельствовать пример
австралопитеков. И качественные
особенности развития больших
полушарий их коры, равно как и
системогенеза мозга человека в целом,
можно познать лишь в свете онто- и
филогенетического развития при
сопоставлении в первую очередь с
характерными чертами структуры,
функций и развития мозга других групп
приматов, в частности узконосых
низших и высших обезьян (Шевченко).
С другой стороны, важнейшее значение
в процессе развития мозга
австралопитеков и формировавшихся древнейших и древних гоминид имело появление и все
более активное применение посредника между организмом и средой в виде орудий, у гоминид
— уже искусственных. В последнем случае опосредованные действия по-новому, качественно
характеризовали постоянный, еще более активный и могучий трудовой фактор преобразования
головного мозга ископаемых людей. При этом, как показывает его эволюция по слепкам
эндокранов, сперва усиливается развитие их задних отделов с сенсорными областями, а затем
передние, которые, в особенности префронтальные, проецируют самую сложную часть
двигательной зоны коры (Кочеткова, 1967) и где у современного человека реализуется
программирование на высшем уровне сознания (Лурия, 1969).
Частные примеры разрастания извилин, например вокруг инсулярной дольки, превращающейся
в островок на дне сильвиевой щели, свидетельствуют об интенсивном процессе развития
извилин, или гирификации, и увеличения количества борозд второго и особенно третьего
порядка, или сулькации, в мозгу человека.
Процесс разрастания коры больших полушарий человеческого мозга есть выражение огромного
преобразования центрального органа высшей нервной деятельности в ходе развития организма
человека под влиянием труда и речи в резкое отличие от эволюции мозга обезьяны под влиянием
одних лишь биологических или, шире, природных факторов.
Если усиленная тарификация и сулькация коры человеческого мозга частично и связаны с тем,
что вызывались самим процессом увеличения его размеров, то основная причина лежит в иной
плос-
224
кости, в качественном преобразовании внутреннего клеточного состава больших полушарий.
Оказывается, что поразительная сложность тонкой архитектоники коры очень сильно
обусловлена и непропорционально богатым составом нервных клеток, а именно невронов со
всеми их отростками.
Рис. 70. Центральная доля большого полушария
мозга человека (видна на дне сильвиевой ямы,
обнажившемся вследствие удаления покрышек,
которые образуются сдвинувшимися здесь
частями лобной, теменной и височной долей)
По Э. Виллигеру, 1930

Если в мозгу орангутана или шимпанзе кора содержит лишь несколько миллиардов невронов, то
в большом мозгу человека кора их вмещает около десяти и даже больше миллиардов. Отсюда,
как это и выясняется при исследовании, огромное усложнение связей между частями мозга, что
опять-таки подтверждает мнение о качественном отличии человеческого мозга от обезьян.
Итак, в свете данных современной науки полностью подтверждается вывод о том, что головной
мозг современного человека является конечным продуктом чрезвычайно долгого хода развития
животного мира. Во время предыстории и в последнее время, когда из обезьян возникли первые
люди, началось под влиянием труда и звукового языка более интенсивное развитие нервной
системы, формировалось человеческое сознание, позволяющее понять законы природы,
расшифровать свое собственное происхождение (Леонтьев, 1959).
Весь ход изучения строения, развития и функций головного мозга человека является сплошным
опровержением религиозных представлений о душе как о частице божественного духа,
управляющей человеческим телом, как о бессмертном двойнике материального тела. Учение о
локализации функций в головном мозгу наглядно показывает полную зависимость всех
психических процессов от нормального функционирования неповрежденных частей мозга и
всего этого наиважнейшего органа в целом (Поляков, 1964).
Уяснению нормальной деятельности головного мозга чрезвычайно способствует глубоко
материалистическое учение И. П. Павлова об условных рефлексах. Созданное величайшим
физиологом мира, это учение неопровержимо доказывает наличие общих закономерностей
деятельности нервной системы млекопитающих животных и человека, общность у них первой
сигнальной системы, помогает уяснить особенности онтогении, филогении, строения и
жизнедеятельности человеческого тела, являющегося продуктом также и более древних стадий
развития животного мира.
Вместе с тем на основе своей физиологической концепции И. П. Павлов пришел к созданию
представления о второй сигнальной системе как специфической для одного человека в отличие
от животных, и трудно переоценить важность этой идеи для исследования самых глубоких
научных, философских проблем, связанных с речью, мышлением.
Рефлекторная теория И. П.Павлова служит разительным естественнонаучным подтверждением
ленинской теории отражения. Тем самым она бросает яркий свет на проблему антропогенеза в
части, касающейся развития мозга как органа мышления.
225
Высшая нервная деятельность
обезьян
Физиологи изучают высшую нервную деятельность животных строго объективным методом
условных рефлексов. Физиологическое учение, созданное русскими учеными И. М. Сеченовым
и И. П. Павловым, рефлекторная теория, учение о нервизме — все это краеугольные камни
единой концепции, позволяющей биологам, в том числе антропологам, правильно и глубоко
подойти к решению даже труднейших вопросов эволюции на основе материализма.
Физиологическое учение И. П. Павлова — сильное, действенное оружие в борьбе против
реакционных течений в зоопсихологии, особенно в США, Англии, ФРГ. Субъективный метод
исследований высшей нервной деятельности должен в конце концов уступить место
объективному. Как известно, И. П. Павлов никогда не позволял трактовать физиологические
опыты по изучению высшей нервной деятельности в психологических терминах. В то же время
он, борясь против дуалистической, идеалистической психологии, утверждал правомерность
существования и развития психологии материалистической. Основы последней лежат в учении