Мак-Фарленд Д. Поведение животных: Психобиология, этология и эволюция
Подождите немного. Документ загружается.


наблюдения - в последние годы дополняется непрямыми методами, основанными
на использовании технических достижений, таких, как звукозапись и
радиотелеметрия. Естественное поведение животных можно изучать и посредством
экспериментов, включающих определенные изменения среды обитания животного
(см. гл. 6 и 23).
В лаборатории исследователь в значительной степени контролирует среду, в
которой находится животное, и поэтому может тщательно планировать
эксперименты для проверки конкретных гипотез, объясняющих различные стороны
поведения животных. Этот подход особенно важен при изучении сенсорных
способностей животных (гл. 12 и 13) и научения у животных (гл. 17 и 18). Помимо
лабораторных исследований все большее значение для понимания поведения
животных приобретают физиологические эксперименты. Физиологический подход
к изучению поведения пропагандировался уже давно, но лишь сравнительно
недавно с его помощью были получены ценные данные (см. гл. 11 и 12).
Поведение животных достаточно сложно, и для того, чтобы хорошо его понять,
необходим широкий набор теоретических и практических подходов. И именно
междисциплинарный подход делает поведение животных такой захватывающей
областью исследований.

Часть
Часть
I
I
.
.
Эволюция поведения
Эволюция поведения
В первой части книги мы обсудим поведение животного с эволюционной точки
зрения. Со времен Чарлза Дарвина стало возможным объяснить роль поведения в
выживании и размножении животного. Если глаза, уши, ноги или крылья можно
считать механизмами, предназначенными для конкретного образа жизни
животного, то к ним же можно отнести и механизмы, контролирующие
поведение. Итак, мы можем спросить, почему у животного наблюдаются
определенные типы поведения, и ожидать, что ответ можно найти, исходя из
эволюционной истории вида.
Эволюционные объяснения такого рода требуют определенных знаний генетики и
развития поведения, теории естественного отбора и эволюционной теории в
связи с социальным поведением животных. Первые три раздела этой книги
посвящены трем перечисленным проблемам.

I.1. Генетика и поведение
Этот раздел, состоящий из трех глав, вводит читателя в область генетики
поведения. В гл. 2 рассматриваются законы Менделя и клеточные основы
наследственности. В гл. 3 приводятся данные, касающиеся развития поведения в
онтогенезе и проблемы врожденного поведения. Гл. 4 посвящена генетике
поведения; она включает обсуждение действия мутаций, полигенной
наследственности и наследуемости поведения.

Грегор Мендель (1822-1884)
(ВВС Hulton Library)
Рожденный в крестьянской семье, Грегор Мендель обучался в монастыре и был посвящен в духовный
сан в возрасте 25 лет. Он готовился стать учителем, но не выдержал квалификационые экзамены на
право преподавания естественной истории в школе высшей категории. В 1856 г. он стал учителем в
школе более низкой категории, ученики которой его очень любили. Мендель начал свои исследования
в 1858 г. в саду при монастыре в Брно (Моравия). Полученные результаты и их теоретические
обоснования были изложены в докладе, прочитанном на заседании Брненского естественнонаучного
общества в 1865 г., и опубликованы в трудах этого общества в 1866 г. Работа Менделя оставалась
неизвестной широкому научному миру по 1900 г., когда на нее обратили внимание более или менее
одновременно и независимо друг от друга три исследователя -Корренз, фон Чермак и де Фриз
(Correns, von Tschermak, de Vries). Ее значение быстро оценили, а результаты, полученные Менделем,
были подтверждены и расширены.
Все свои основные опыты Мендель провел на обычном садовом горохе. Его особенно интересовали
такие признаки гороха, которые четко различаются у разных особей. К числу таких признаков
относятся окраска цветков (красная или белая), форма плода (гладкая или с перетяжками), окраска и
форма семян (зеленые или желтые, гладкие или морщинистые), расположение цветков (пазушные или
верхушечные), длина стебля (длинный или короткий). Мендель высевал каждый сорт отдельно.
Вручную перекрестно опылив выросшие растения, он получил различные гибриды на разных
делянках. Не пытаясь оценить поколения в целом, он изучал наследование определенных признаков у
всех потомков конкретной пары. На основе полученных данных Мендель сформулировал законы
наследственности, произведшие революцию в биологии. В 1866 г. Мендель написал знаменитому
биологу Карлу фон Негели, который использовал в качестве экспериментального объекта ястребинку.
Описав свои опыты и сформулировав гипотезы, Мендель попросил помощи у Негели и предложил
повторить проведенные им эксперименты на ястребинке. Однако сообщение Менделя не произвело
впечатление на Негели, и он предложил Менделю продолжить работу на большем числе экземпляров
гороха. Мендель к этому времени провел наблюдение уже на 13 000 растений гороха и теперь пытался
повторить работы Негели на ястребинке. Но эти растения малы, их трудно опылять искусственно, и
они не годятся для работ по генетике. Некоторое время Мендель упорствовал, но успеха не достиг.
Негели предложил вырастить семена гороха в ботаническом саду в Мюнхене, и Мендель послал ему
140 пакетов. Однако Негели не вырастил эти семена и не сослался на Менделя, публикуя свою
основную работу в 1884 г. Забавно, что фон Негели, первым открывший и описавший хромосомы под
микроскопом в 1842 г., так и не понял важности работ Менделя. Связь между хромосомами и
менделевской наследственностью была установлена лишь в 1914г. В 1875 г. монастырь в Брно был
втянут в налоговую тяжбу, и Менделю, бывшему прелатом в этом монастыре, не осталось времени
для наблюдений. Умер он в 1884г.

2. Гены и хромосомы
После опубликования теории естественного отбора Чарлза Дарвина острый интерес
ученых вызвал вопрос о том, как наследуются признаки и как поддерживается
изменчивость у потомства. Фон Негели, знаменитый благодаря своим работам по
клеточному строению тканей, провел эксперименты по скрещиванию на
ястребинке. Бэтсон (Bateson) опубликовал свои обширные исследования по
наследственности и изменчивости в 1894 г. Голландский ботаник Гуго де Фриз
обнаружил, что измененные растения энотеры появляются от нормальных растений
и, по-видимому, дают потомков такого же генотипа; он назвал эти внезапные
резкие изменения мутациями. В 1868 г. Дарвин опубликовал свою книгу
«Изменения животных и растений под влиянием одомашнивания, в которой он
выдвинул идею пангенеза, гласившую, что каждая клетка тела выделяет
мельчайшие «геммулы», содержащие информацию о ней. «Они размножаются и
собираются вместе в почках и половых элементах». Однако до 1900 г., пока не
получили признания работы Грегора Менделя, генетику нельзя было увязать с
теорией эволюции путем естественного отбора и с развивающейся клеточной
физиологией.
В этой главе будет показано, каким образом работы Менделя позволили прийти к
современному пониманию клеточных основ наследственности, генетических
изменений и природы гена. Генетика имеет фундаментальное значение при
исследовании поведения животных, поскольку многое в поведении определяется
генетическим своеобразием животного. Постигнув законы генетики, мы сможем
понять природу такого влияния.
2.1. Естественный отбор и поведение
Естественный отбор оперирует с физическими признаками, или фенотипом, особи;
в число таких признаков входит и ее поведение. У особи мыши, которая не
пытается спастись от хищника, меньше шансов выжить, чем у мыши, которая
делает это, а мышь, которая не использует благоприятную возможность для
получения пищи, также окажется в невыгодном положении. Таким образом, к
признакам, имеющим важное значение для выживания животного, мы можем
отнести не только его морфологические признаки, но и характер поведения. Более
того, соотношение между различными типами поведения также может быть
важным. Например, мышь, которая уделяет слишком много внимания пище, может
не заметить приближающегося хищника, а мышь, которая слишком боится
хищников, может упустить возможность для получения пищи. Такие соображения
наводят на два важных вопроса о связи между естественным отбором и
поведением: 1) как можно определить или продемонстрировать значение для
выживания конкретных типов поведения? 2) существует ли для животного
наилучший способ распределить свое время между различными видами
активности, возможными при данных условиях? На первый из этих вопросов мы
ответим в разд. 6.1. Второй вопрос несколько сложнее и освещается в гл. 24.
Эффективность естественного отбора в изменяющихся популяциях зависит от
степени, в которой наследуются феноти-пические признаки. Дарвин знал, что среди
потомков от любых конкретных роди-
25

телей обычно наблюдается изменчивость и наличие этой изменчивости лежит в
основе его теории, однако он не мог сказать, как возникает изменчивость. Лишь в
1858 г. Грегор Мендель, моравский монах, начал эксперименты по скрещиванию
растений. Он описал свои результаты в 1865 г., но в относительно малоизвестном
издании, и они привлекли внимание научного мира лишь в 1900 г., уже после
смерти Дарвина. Мендель показал, что наследственные признаки не смешиваются,
они дискретны: потомки наследуют отдельные частицы, которые мы теперь
называем генами. Открытие заново работ Менделя привело к созданию Р. А.
Фишером и другими исследователями в 1930 г. генетической теории естественного
отбора. Согласно этой теории, в основе механизма действия естественного отбора
лежит изменчивость, существующая в популяциях свободно скрещивающихся
животных.
При изучении поведения животных важно знать, в какой степени наследуются
конкретные поведенческие признаки. Эти знания позволяют нам оценить степень
действия отбора на поведенческие признаки и, кроме того, выявляют различия
между врожденным и приобретенным поведением - вопрос, вызывающий
постоянные дискуссии с далеко идущими в философию и политику выводами.
В гл. 4 рассматривается генетика поведения. Для тех целей, которые мы
преследуем в данной книге, важно понимать, что эффективность влияния
естественного отбора на эволюцию зависит от того, насколько поведение
контролируется генетически. Эволюция поведения осложняется тем, что
естественный отбор не всегда является единственным важным механизмом. У
некоторых животных поведенческие признаки могут эволюционировать при
участии других механизмов; у человека, в частности, - посредством культурной
преемственности. Особи могут научиться различным навыкам, заимствуя их друг у
друга (см. гл. 27), и таким образом информация переходит от одного поколения к
другому. Поведение птиц и млекопитающих есть результат сложного
взаимодействия генетических факторов и приобретенного опыта. Чтобы
разобраться в этом взаимодействии, этолог должен быть знаком с основами
генетики.
2.2. Законы Менделя
Большую часть экспериментов Мендель проводил на садовом горохе. В одном из
экспериментов он скрестил растения с красными цветками и растения с белыми
цветками: у всего потомства цветки оказались красными. Затем он произвел
скрещивание между особями дочернего поколения и получил 705 растений с
красными цветками и 224 растения с белыми. Эти результаты важны в двух
отношениях. Прежде всего в первом дочернем поколении не было растений с
белыми цветками, а во втором поколении такие растения были. Кроме того,
отношение красных и белых цветков во втором поколении составляло 3:1. Дарвин
также проводил эксперименты на горохе и также получил отношение 3:1. Однако
Мендель установил это и другие важные отношения в многократно повторенных
экспериментах. Помимо этого, он понял причину исчезновения одного из
родительских типов (белого) среди потомков первого поколения. Мендель
предположил, что у каждого растения гороха окраску цветков, так же как и любой
другой признак, определяют два наследственных фактора. Один фактор
доминирует над другим, поэтому когда в растении присутствуют два разных
фактора, цветки могут быть только одной окраски. Следовательно, у растений с
белыми цветками имеются два белых фактора, тогда как у растений с красными
цветками - два красных или красный и белый факторы. Такой способ рассуждений
позволил Менделю объяснить все наблюдавшиеся соотношения типов потомства и
сформулировать законы наследования, совершившие переворот в биологической
науке.
На основе своих экспериментов по скрещиванию гороха Мендель пришел к ряду
заключений:
1. Наследственность дискретна, и генетический вклад каждого родителя одинаков.
Генетический материал одного родителя не смешивается или не сливается с
26
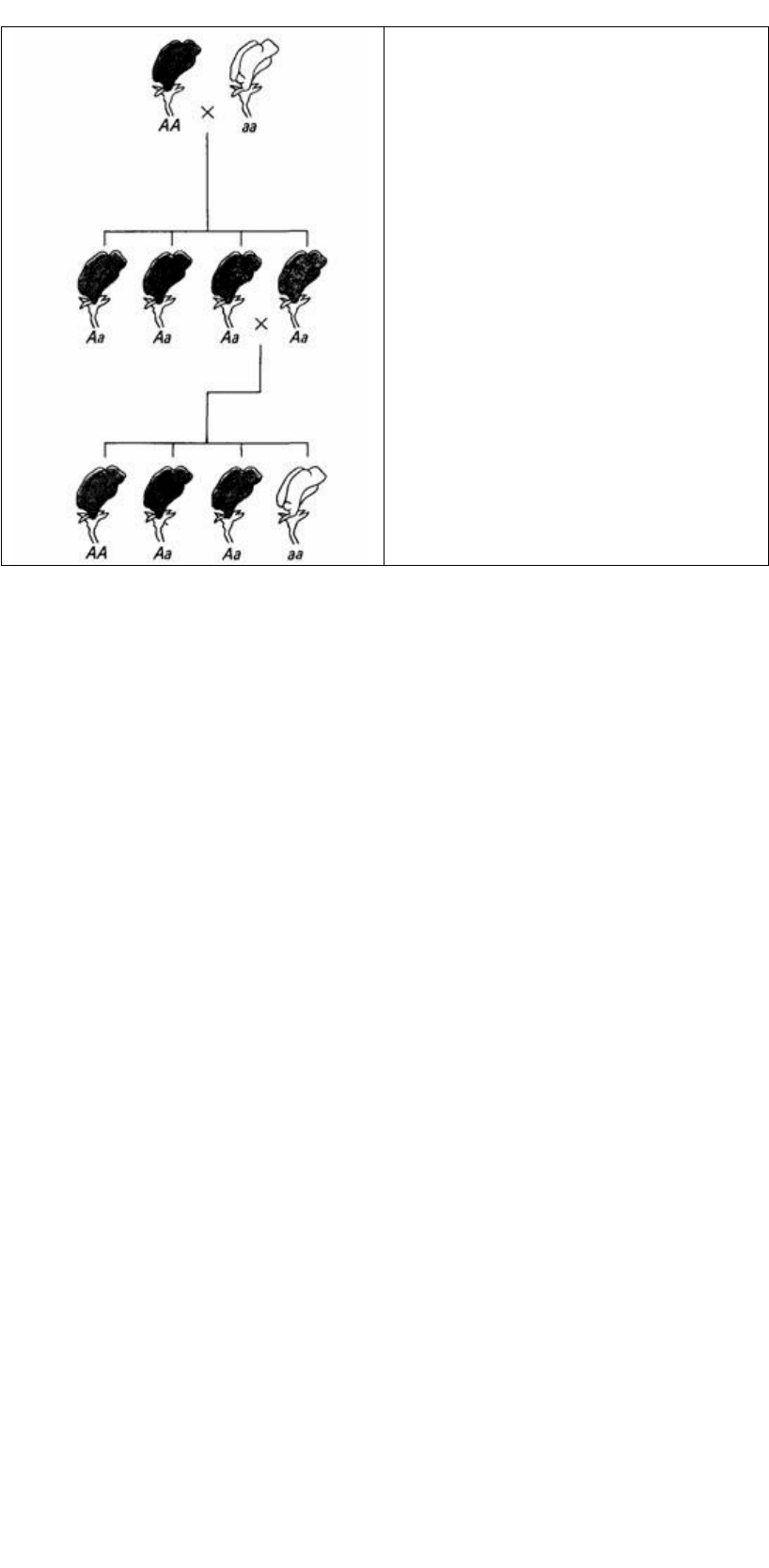
Рис. 2.1. Схема опытов Менделя по скрещиванию
сортов гороха, различающихся по окраске
цветков (красные и белые). Красная окраска (А)
доминирует над белой (а).
генетическим материалом другого родителя. Дискретные факторы Менделя теперь
называют генами.
2. Любой признак, проявляющийся в строении или внешнем виде особи,
контролируется парой генов, каждый из которых наследуется от одного родителя.
Совокупность этих внешних признаков называется фенотипом.
3. Каждый ген может существовать в двух или более альтернативных формах,
называемых теперь аллелями. Совокупность всех генов особи называется
генотипом. Таким образом, при двух аллелях возможны три их комбинации. У
особи, содержащей два альтернативных аллеля А и а, возможны три генотипа: АА,
аа и Аа. Если особь несет два одинаковых аллеля, как в первых двух случаях, то
говорят, что особь гомозиготна по данному признаку. Если особь несет по одной
копии альтернативных аллелей, как в третьем случае, то говорят, что этот организм
гетерозиготен.
4. Один аллель может доминировать над другим, и в этом случае недоминантный
аллель называют рецессивным. Доминантные гены определяют фенотип в
гомозиготном и гетерозиготном состояниях. Рецессивные гены определяют
фенотип только в гомозиготном состоянии (рис. 2.1).
5. Из альтернативных генов каждого родителя потомком наследуется только один.
Другую копию потомок получает от другого родителя. Таким образом, в первом
дочернем поколении (F
1
возможен только один фенотип (первое поколение,
следующее за межсортовым скрещиванием), но более одного во втором дочернем
поколении (F
2
) (рис. 2.1). Этот закон расщепления (иногда называемый первым
законом Менделя) дает в результате типичное менделевское отношение фенотипов
3:1 (рис. 2.1).
6. Если две или более пар генов расщепляются одновременно, то распределение
какого-либо одного гена не зависит от распределения остальных. Это называется
законом независимого распределения генов или вторым законом Менделя. Из этого
закона следует, что признаки, проявляющиеся вместе в одном поколении, не
обязательно должны проявиться у одной особи в следующем поколении (рис. 2.2).
Законы наследования Менделя справедливы для многих видов растений и
животных, но они все же не носят универсальный характер. Одно из отклонений от
классического менделевского расщепления является следствием вариабельности
связи между наследственной структурой, или генотипом, и наблюдаемыми
признаками, или фенотипом. Для большинства признаков, изученных Менделем,
доминирование альтернативного аллеля было полным. Однако теперь мы знаем,
что существует определенный диапазон возможных проявлений гена (рис. 2.3) и
что доминирование не всегда бывает полным.
Другое осложнение простой менделев-ской картины заключается в том, что данный
ген может влиять на множество разных признаков. Это явление известно
27
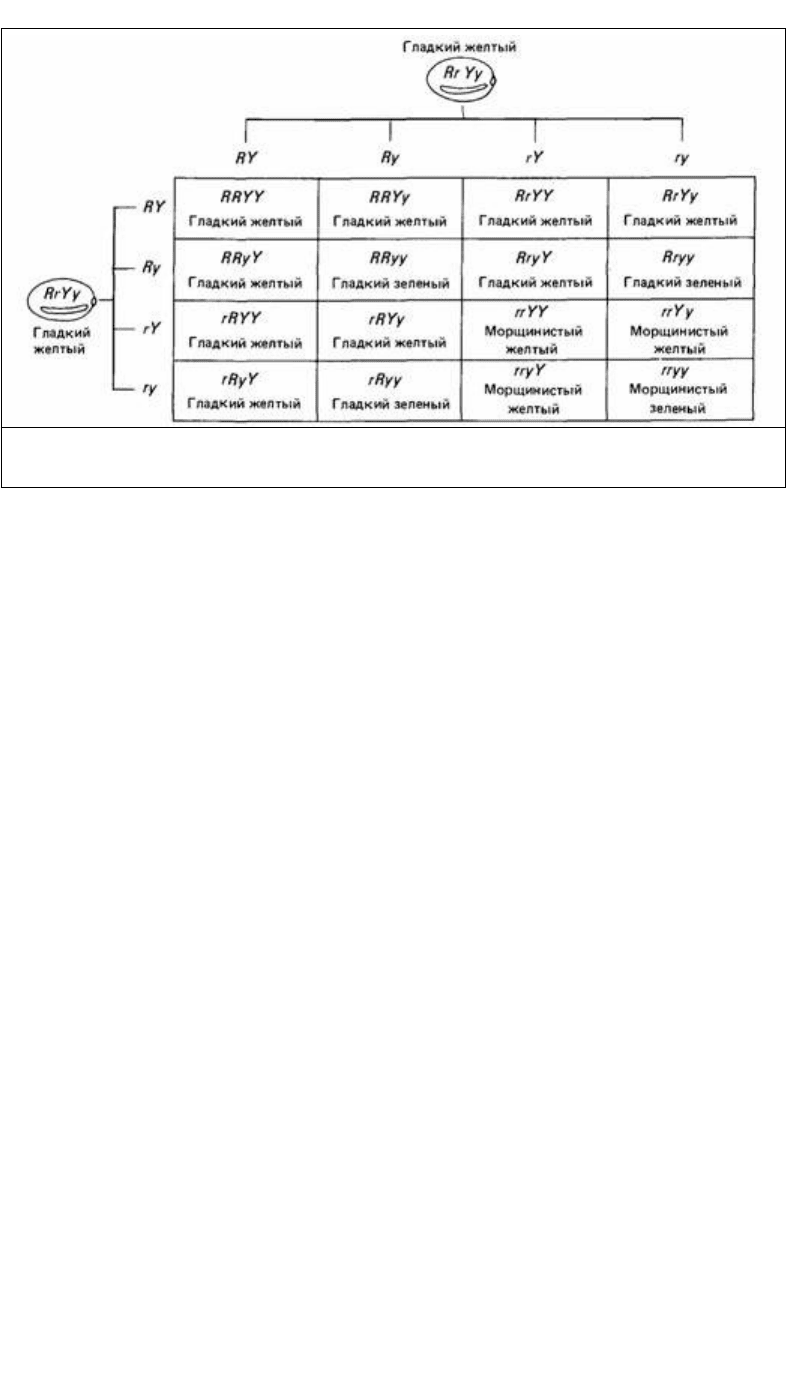
Рис. 2.2. Скрещивание двух гибридов первого поколения (F
1
) в одном из опытов Менделя на горохе.
Гладкие семена (R) доминируют над морщинистыми (r). а желтые (Y) над зелеными (r).
как плейотропия. Впервые плейотропию в поведении продемонстрировал Стёрте-
вант (Sturtevant) в 1915 г. Работая с плодовой мушкой Drosophila, он обнаружил,
что самцы мух предпочитают спариваться с мутантными самками, имеющими
белые глаза или желтое тело. Кроме того, он обнаружил, что самки разных типов
избегают спариваться с желтотелыми и белоглазыми самцами. Эти результаты
связаны с проявлением плейотропии, поскольку и у белоглазых, и у желтотелых
мутантов в качестве побочного эффекта наблюдается снижение общего уровня
активности. Поскольку нормальные активные самки имеют тенденцию избегать
самцов, мутантные самки оплодотворяются чаще. Точно так же, поскольку самцы
должны найти самок, чтобы спариться с ними, у неактивных мутантных самцов
шансы на успех ниже, чем у их нормальных соперников. Другими словами,
мутантные гены плейотропны, т. е. один ген обусловливает проявление нескольких
признаков. В ситуации, где спаривание происходит случайным образом, мутантные
мухи будут элиминироваться из популяции вследствие их низкой сексуальной
активности.
Отклонение от простых менделевских законов может быть обусловлено также
взаимодействием генов. Если два или более генов определяют один признак, то в
результате их взаимодействия могут появляться особи с новым фенотипом.
Например, форма гребня у домашних кур определяется двумя генами, R и Р.
Доминантный ген R дает розовидный гребень (рис. 2.4), а его рецессивный аллель r
в гомозиготном состоянии - простой гребень. Доминантный ген Р дает
гороховидный гребень, а его рецессивный аллель р — простой. Если гены R и Р
окажутся вместе в одном генотипе, то в результате их взаимодействия образуется
ореховидный гребень, который не детерминируется ни одним из этих генов
самостоятельно.
Другая форма взаимодействия генов - комплементарность генов; в этом случае
гены взаимозависимы, и ни один из них не может определить фенотипический
признак без другого. Иногда один ген маскирует действие другого. Это явление
называют эпистазом. Эпистаз похож на доминирование, но доминирование
включает различные аллели, а эпистаз - различные гены.
Ген-модификатор может изменять экспрессию другого гена. Так, например, цвет
глаз у человека определяется в основном одной парой генов. Ген В детерминирует
карие глаза, а ген b-голубые. Генотипы ЕЕ и ВЬ определяют карие глаза,
28
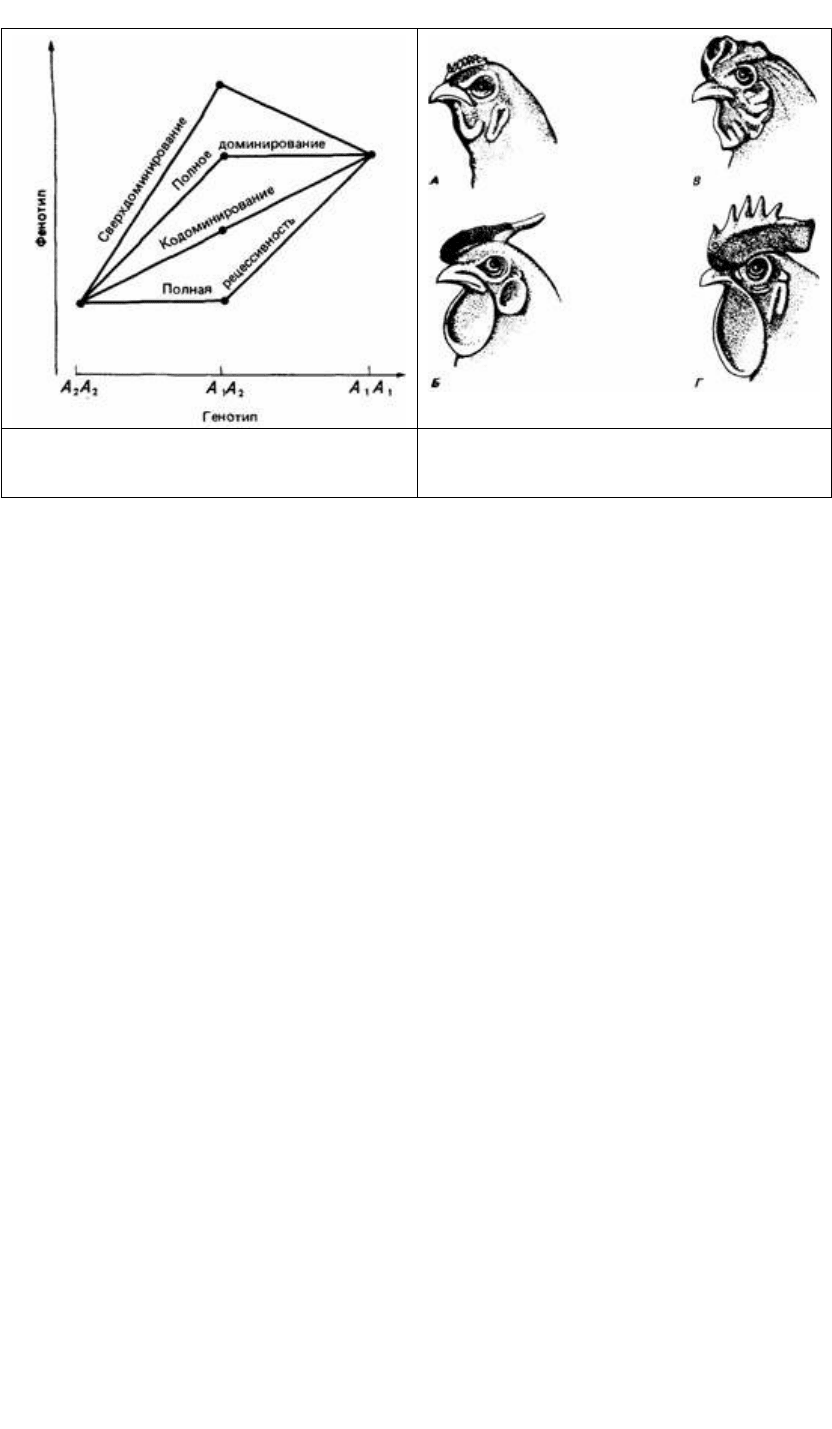
Рис. 2.3. Графическое представление четырех
различных типов экспрессии генов. (По МсClearn,
DeFries, 1973.)
Рис. 2.4. Типы гребня у кур. А - гороховидный; Б -
розовидный; В - ореховидный; Г - листовидный.
а генотип bb- голубые. Однако благодаря действию генов-модификаторов
существует множество вариаций этих двух основных цветов глаз. Гены-
модификаторы могут включать фенотипическое действие других генов.
На некоторые фенотипические признаки человека, например на рост, оказывает
влияние большое число генов, и поэтому такие признаки широко варьируют у раз-
ных членов популяций. Что касается например роста, то даже если его определяют
всего лишь несколько генов, то мы можем ожидать, что одни значения роста будут
преобладать в большей степени, чем другие.
2.3. Клеточные основы наследственности
Представление о том, что растения и животные состоят из клеток, развивалось
постепенно; впервые вполне определенно это высказал в 1839 г. Шванн (Schwann),
установивший связь между клеточным строением и ростом растений и животных.
В 1840 г. фон Кёлликер (von Koelliker) установил, что сперматозоиды и яйца
представляют собой клетки; вскоре было выяснено, что все живые ткани состоят из
клеток.
В 1842 г. фон Негели впервые зарегистрировал тот факт, что деление клетки
включает деление ядра и его хромосом; однако более подробное описание процесса
деления клеточного ядра еще ждало усовершенствования микроскопической
техники. Первую подробную и точную запись о поведении хромосом во время
клеточного деления сделал в 1882 г. Флемминг (Flemming), наблюдавший этот
процесс в клетке личинки саламандры. К 1885 г. ван Бенеден (van Beneden)
обнаружил, что хромосомы не изменяются при последовательных клеточных
делениях. Он обнаружил также, что число хромосом постоянно у каждого вида. В
1856 г. Прингшейм (Pringsheim) впервые наблюдал слияние сперматозоида с
яйцеклеткой, но только Хертвиг (Hertwig) в 1876 г. понял, что в тех случаях, когда
в оплодотворенной яйцеклетке обнаруживаются два ядра, одно из них должно
принадлежать сперматозоиду. Таким образом, он предположил, что именно
хромосомы несут генетический материал, который предоставляется обоими
родителями в момент оплодотворения. Все это создавало ясную картину
клеточного деления и оплодотворения, подготавливая цочву для быстрого
восприятия работ Менделя о наследственности, которые вновь увидели свет в 1900
г.
29
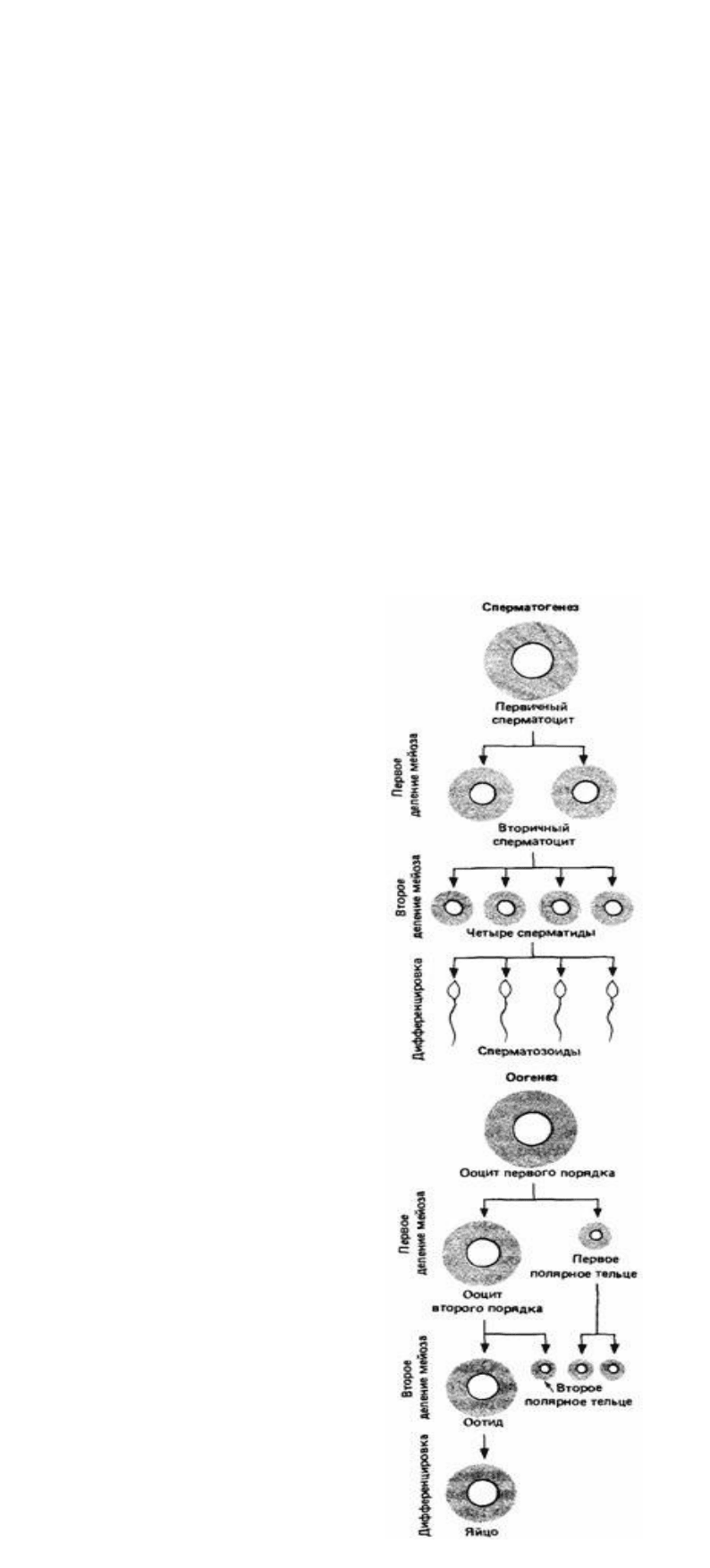
Соматические клетки большинства животных диплоидны, т.е. в каждой клетке
содержатся две копии хромосом одного типа. В соматических клетках человека
содержится 46 хромосом, или 23 различные пары, а у плодовых мушек дрозофил -
восемь хромосом, образующих четыре различные пары. В результате нормального
клеточного деления, или митоза, образуются диплоидные клетки. Клетки всех
типов, за исключением половых, которые дают начало мужским и женским
гаметам- сперматозоиду и яйцеклетке,-образуются посредством митоза. Поскольку
сперматозоид и яйцеклетка, сливаясь, образуют одну клетку с двумя наборами
хромосом (диплоидную зиготу), каждая гамета несет только один набор хромосом
и называется гаплоидной. Поэтому при образовании половых клеток диплоидное
число хромосом уменьшается до гаплоидного в результате процесса, называемого
мейозом.
При мейозе происходят два самостоятельных клеточных деления. Во время первого
деления уменьшается число хромосом, а второе деление похоже на обычный митоз,
за исключением того, что дочерние клетки гаплоидны. Полный мей-оз, таким
образом, дает в результате деления одной диплоидной клетки четыре гаплоидные.
У мужских особей все четыре гаплоидные клетки дифференцируются в
сперматозоиды (рис. 2.5). У женских особей во время первого клеточного деления
обычно образуются клетки разной величины. Маленькая клетка, которую называют
первым полярным тельцем, второй раз может и не делиться. Большая клетка снова
претерпевает неравное деление в процессе которого образуются второе полярное
тельце и клетка, дифференцирующаяся в яйцо (рис. 2.5). Полярные тельца не
сохраняются. Таким образом, во время мейоза у самцов обычно образуются четыре
сперматозоида одинакового размера, а у самок - одно большое яйцо и два или три
нефункционирующих полярных тельца.
Рис. 2.5. Схема сперматогенеза и оогенеза у животных.
30