Каминская Э.А. Общая генетика
Подождите немного. Документ загружается.

'167
Образование
экстраэмбрцональной
энтодермы
Имплантация, .
рост внутренней
клеточной массы
Разбитие й
трофобластах
эндодерма IL
Ранний £
яйцевой ш
\ цилиндр $
t'
2
блас/ло-
\ Цис/77
Цилиндр
яйца
(urS^Or
/ Рост
цилиндра
яйца
М0/?<//70
/ Поздний
/ яйцевой цилиндр
Образование т
примитивнои
полоски
Образование нотохорды,
невральнои трубки, сомитов
Эмбриональная
_ олас/нинка
Сформировавшийся
эмдрион
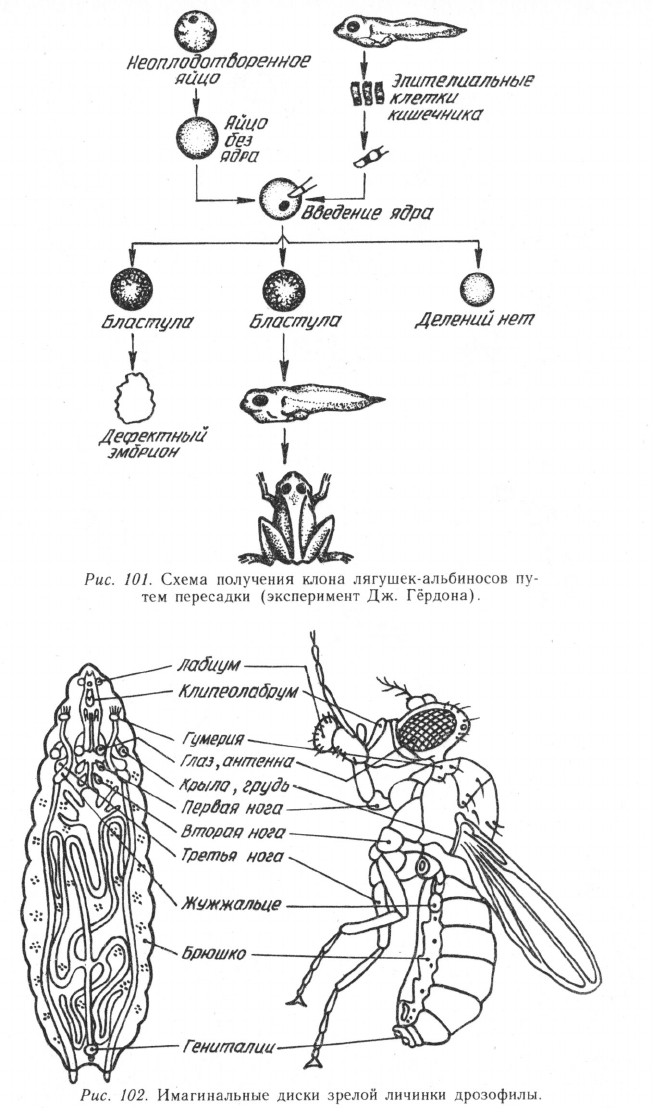
Рис. 103. Формы нарушения эмбриогенеза у мышей, гомозиготных по некоторым
мутациям локуса Т (по Дж. Беннету, 1975).
У мексиканского аксолотля обнаружена рецессивная летальная
мутация по гену О, обусловливающая гибель гомозиготных особей
на стадии гаструлы. У ннх формируется дефектная цитоплазма,
которая репрессирует функции всех генов, и у гомозигот в эмбрио-
нальных клетках не образуется и-РНК. Если в такие клетки добавить
небольшое количество нормальной цитоплазмы, функция генов вос-
станавливается, что обеспечивает нормальное развитие организма.
Регуляция дифференциальной активности генов может, по-види-
мому, осуществляться на уровне как транскрипции (генетический
уровень), так и трансляции (постгенетический уровень). В первом
случае в клетке контролируется активность генов и, следователь-
168

Рис. 104. Дифференциальная активность генов на участке политенной хромосомы
слюнных желез личинок комара на разных стадиях развития (из Р. Л. Берг и
С. Н. Давиденкова; 1971):
а — авторадиография; б — рисунок; в — микрофотография.
но, количество и состав и-РНК; во втором — активность пост-
транскрипционной и-РНК и трансляции.
Одним из доказательств дифференциальной генной активности в
период органогенеза служит процесс формирования пуфов в поли-
тенных хромосомах слюнных желез Drosophila melanogaster и Chi-
ronomus tentans. Ряду авторов, в том числе И. И. Кикнадзе (1964—
1965), удалось обнаружить у этих насекомых так называемые стадие-
специфические пуфы, появляющиеся только на определенных ста-
диях развития. Оказалось, что у Chironomus повышенная активность
пуффинга проявляется в критические периоды развития — во время
личиночных и метаморфозных линек. При этом около 40 % пуфов,
возникающих между линьками (стабильные пуфы), увеличивается
в размерах, некоторые же из них уменьшаются или вообще исчезают.
Одновременно формируются новые стадиеспецифические пуфы.
У дрозофилы максимальная активность пуфов проявляется в период
превращения личинки в куколку (рис. 104). Все эти процессы контро-
лируются генотипом. Например, некоторые мутации у особей дрозо-
филы существенно меняют их способность образовывать пуфы в опре-
деленных группах дисков.
12. Зак. 5107
169
Дифференциальная активность свойственна не только отдельным
генам, но и родительским хромосомам и даже геномам в целом.
Так, у самцов мучнистого червеца хромосомный набор, полученный
от отца, претерпевает полную гетерохроматинизацию. В результате
отцовские гены инактивируются и даже доминантные признаки не
проявляются. Иногда в процессе развития гетерохроматиновые
хромосомы активизируются, переходят в эухроматиновое состояние,
и тогда некоторые ткани приобретают способность наряду с мате-
ринскими признаками нести признаки отца. У самок млекопитаю-
щих на ранних этапах эмбриогенеза может инактивироваться одна
половая хромосома: либо материнская, либо отцовская. Такие не-
активные ^-хромосомы представляют собой гетерохроматиновые
тельца. Однако они свойственны не всем клеткам. Их нет, например,
в клетках лимфоидного ряда, и это дает основание предполагать,
что обе половые хромосомы в данных клетках активны. Приведенные
сведения говорят о том, что в клетках имеются факторы, контроли-
рующие дифференциальную активность генов. Такими факторами,
вероятно, можно считать ооплазматическую сегрегацию (разделение
цитоплазмы яйца на зоны) и химическое влияние одного зачатка
на другой. Их природа и механизмы действия не установлены.
Регуляторами клеточной дифференцировки могут быть и гормо-
ны. В силу того что они распределяются по всему организму равно-
мерно, их регулирующее влияние сводится скорее не к определению
места дифференцировки, а к установлению момента ее начала и к ко-
ординации одновременных процессов в разных тканях и органах.
Так, фактором гормональной регуляции генной активности у насе-
комых служит экдизон — стероидный гормон линьки, от которого
зависят локализация и порядок появления пуфов в политенш
хромосомах. Доказательством гормонального влияния на процессы
клеточной дифференцировки является эксперимент по пересадке
у дрозофилы имагинальных дисков. Если диск разрезать пополам
и одну часть пересадить в брюшную полость личинки, а другую —
взрослой особи, в первом случае разовьется нормальный орган,
а во втором дифференциации клеток не произойдет и диск просто
«дорастет» до определенного размера. Это объясняется тем, что у
личинки имеются гормоны, контролирующие дифференцировку,
а у взрослой особи их нет. Гормоны у насекомых и амфибий контро-
лируют смену стадий метаморфоза, у большинства животных —
процессы гаметогенеза, лактации, половой активности. Нередко
введением гормона удавалось предотвратить развитие аномального
признака, формирующегося в результате нарушения структуры
гена. Так, у мышей с мутацией карликовости и недоразвитием гипо-.
физа после введения гормона его передней доли исчезали симпто-
мы поражения.
Установлено, что у человека соматотропный гормон роста влияет
на физическое развитие: при избытке гормона развиваются акроме-
галия и гигантизм, при недостатке — нанизм (гипофизарная карли-
ковость). Гиганты могут иметь как гармоническое телосложение,
так и признаки акромегалии (увеличение стоп, кистей, носа и т. д.).
170
CaivibiM высоким человеком на Земле был англичанин Лускин —
у него был рост 265 см. В Африке в настоящее время живут самые
высокие люди (2-метровые динка, шиллук, нуэр и племя ватуси) и
самые низкие (у бушменов рост около 150 см, у пигмеев— 140—
150 см у мужчин, 120—130 см у женщин). Низкорослые расы живут
также на Андаманских и Филиппинских островах, на полуостровах
Малакка. Низкорослость у них обусловливается пониженной чувстви-
тельностью их клеток и тканей к соматотропному гормону, хотя его
содержание в крови обычно бывает в пределах нормы. Встречаются
два типа гипофизарной карликовости, обусловленной недостатком
соматотропного гормона: сексуальный (характерна изолированная
недостаточность гормона роста) и асексуальный (свойственна не-
достаточность всех гормонов передней доли гипофиза — гонадотро-
пина, тиреотропина и адренокортикотропного гормона). У сексу-
альных карликов сохраняется нормальное половое развитие и они
могут иметь потомство, причем дети у них всегда бывают карликами.
Внутриутробно они развиваются нормально. Задержка роста насту-
пает лишь в возрасте нескольких месяцев, когда соматотропный
гормон становится необходимым для развития. У асексуальных кар-
ликов нарушены функции половых желез, поэтому они всегда бывают
бесплодны. Предполагается, что для обоих типов гипофизарной кар-
ликовости типична рецессивная форма наследования. Вероятно, ре-
цессивная мутация приводит к нарушению функции гена, контро-
лирующего синтез гормона роста.
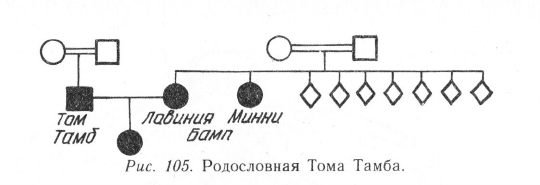
Известен случай наследования карликовости по рецессивному типу. У Чарльза
Стреттона («Генерал Том Тамб») и его жены Лавинии Бамп было нормальное
телосложение и рост соответственно 95 и 80 см. Есть сведения, что у них был ребе-
нок. Их родители состояли в родственном браке (двоюродные брат и сестра) и, оче-
видно, являлись гетерозиготными носителями рецессивного гена карликовости. Их
рост был нормальным. В семье Л. Бамп из 9 детей двое были карлики — Лавиния
и ее сестра Минни (рис. 105).
Оба описанных типа гипофизарной карликовости удается лечить
гормоном роста в период, пока зоны роста костей остаются от-
крытыми.
Существует еще один тип гипофизарной карликовости, при ко-
торой гормон роста образуется в избытке, но восприимчивость к нему
специфических рецепторов ограничена. У такой карликовости реакция
на введенный гормон ослаблена, и лечение обычно не дает результата.
Нарушение регуляции развития на генетическом и постгенетиче-
ском уровнях нередко приводит к замещению структур одной части
'171
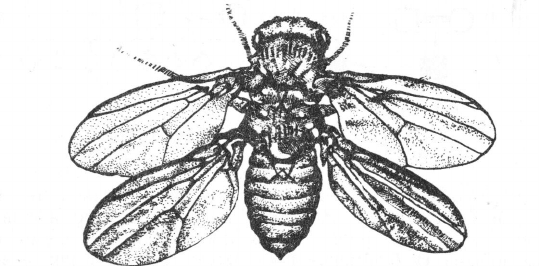
Рис. 106. Четырехкрылый мутант с удвоением грудного
сегмента у дрозофилы (по Е. Льюису, 1982).
организма гомологичными структурами, обычно формирующимися в
другом месте. В результате этого процесса, называемого гомеозисом
(от гр. homoios — сходный), развиваются новые, необычные при-
знаки, а также уродства развития. Описан гомеозис у членистоно-
гих, причем лучше всего он изучен у дрозофилы. В основе морфо-
генеза у насекомых лежит генетическая детерминация клеток бласто-
дермы. Они «выбирают» какой-то конкретный путь развития, и на
каждом из них проявляется избирательная активность строго спе-
цифических генов, контролирующих развитие различных частей
тела: конечностей, крыльев, гениталий, глаз и т. д. Таким образом,
еще на стадии бластодермы определяется судьба клеток, образующих
впоследствии имагинальные диски. В дальнейшем из каждого диска
развивается та или иная ткань либо орган. Контроль за нормаль-
ным развитием насекомого осуществляют специальные гомеозисные
гены. По-видимому, кодируемый ими белок выполняет функцию
регулятора генной активности.
Вследствие нарушения нормального развития клеток имагиналь-
ных дисков, уже детерминированных к развитию в определенном на-
правлении, у дрозофилы возникают различные нарушения формооб-
разования: гомеозисные мутации, гомеозисные фенокопии и транс-
детерминации.
Причиной гомеозисных мутаций является нарушение функции го-
меозисных генов в некоторых клетках имагинальных дисков. Оно
обусловливает развитие из имагинального диска структур, в норме
продуцируемых другим имагинальным диском. К примеру, вместо
антенны может сформироваться конечность (мутация Antennapedia),
вместо глаза —-крыло (мутация Ophtalmopedia), вместо нормальной
груди — ее удвоение и появление дополнительной пары крыльев
(мутация — bithorax; рис. 106).
Гомеозисные фенокопии по эффекту аналогичны гомеозисным
мутациям, но обусловливаются нарушениями развития на постгене-
тическом уровне, вызванными неблагоприятными внешними факто-
рами. К примеру, у дрозофилы вместо жужжалец может развиться
одно или пара недоразвитых крыльев.
'172
При гомеозисе вследствие трансдетерминации возникают нару-
шения имагинального диска, уже детерминированного к развитию в
определенном направлении. Так, при пересадке генитального имаги-
нального диска, взятого у куколки, в брюшную полость взрослой мухи
из него может развиться другая часть тела (например, конечность).
Генотип и фенотип
В процессе развития яйцеклетки под влиянием условий внешней
среды в результате взаимодействия ядра и цитоплазмы наследст-
венная информация, закодированная в генотипе, реализуется в фено-
тип. Генотип представляет собой наследственный потенциал организ-
ма. Он лишь обусловливает возможность развития определенных
признаков и свойств, т. е. фенотипа. Характер же реализации на-
следственной информации зависит от того, какие гены в тот или иной
период развития проявляют действие. Вместе с тем формирование
фенотипа контролируется также факторами окружающей среды,
оказывающими влияние на функциональную активность генов.
Закономерности реализации наследственной информации в про-
цессе развития признака изучает феногенетика. Она рассматривает
такие вопросы, как характер взаимоотношений ядра и цитоплазмы в
развитии, сущность генетического контроля процессов биосинтеза,
клеточной дифференцировки и др.
Еще в 30-е годы С. С. Четвериков с сотрудниками пришли к
выводу, что, с одной стороны, «каждый ген влияет на все признаки
организма, хотя его влияние на некоторые из них может быть исче-
зающе мало», с другой — «любой признак зависит от всех генов всего
генотипа в целом, хотя бы зависимость от некоторых генов и была
незаметная».
Впоследствии было установлено, что процесс формирования от-
дельных признаков зависит от характера генетической детермина-
ции, т. е. что наследование может быть моно- и полигенным, и что
проявление некоторых признаков обусловливается взаимодействиями
неаллельных и сцепленных генов.
В обычных условиях наследственные факторы контролируют фор-
мирование нормального фенотипа. При изменении же структуры
гена (мутация) нормальное развитие организма нарушается: нор-
мальный белок, контролируемый данным геном, либо не образуется
вовсе, либо синтезируется какой-то другой белок с иным амино-
кислотным составом. В результате вместо одного признака формиру-
ется другой. К примеру, у человека рецессивная мутация гена, ответ-
ственного за синтез фермента, участвующего в метаболизме опре-
деленных гликолипидов (ганглиозидов), обусловливает нарушение их
метаболизма, в результате чего липиды накапливаются в клетках
мозга и последние перестают нормально функционировать. У де-
тей грудного возраста при такой мутации развивается тяжелая бо-
лезнь — болезнь Тея-Сакса, приводящая к задержке развития, пара-
личам, слабоумию, слепоте и в конечном итоге — к смерти (в первые
два года жизни).
173
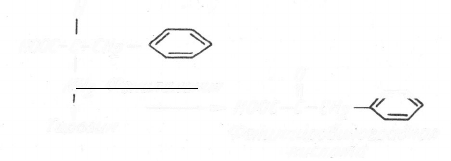
Аналогичный эффект вызывают рецессивные мутации, приво-
дящие к развитию фенилкетонурии и альбинизма. При фенилкетону-
рии, например, нарушается метаболизм фенилаланина на стадии его
превращения в тирозин и в организме накапливается фенилпиро-
виноградная кислота (оказывает токсичное влияние на клетки
мозга ребенка):
Н
Н00С-С-СН2 ~
' О
А/И2 Фенилалаяин ц ч
f — Н00С-С-СН
2
Тирозин Фенилпиробиноградная
кислота
Во всех описанных случаях отчетливо прослеживается действие
мутации одного гена на формирование многих признаков.* Примером
такого же плейотропного влияния гена служит наследование корот-
коногости у кур (ген коротконогости оказывает летальное действие
у гомозиготных по нему особей) и гидроцефалии у мышей. Послед-
няя проявляется целым рядом признаков: аномальные число и рас-
положение вибриссов, незакрытие век у зародыша, атипичная форма
гипофиза, отсутствие плоских покровных костей черепа, кровоизлия-
ние в мозг, приводящее к смерти сразу после рождения. Гидроце-
фалия обусловливается изменением структуры одного гена, приво-
дящим к множественным нарушениям морфогенеза. При этом можно
установить начало формирования нарушений и проследить весь его
путь: замедленный рост передней доли гипофиза, обусловленный
мутацией, вызывает нарушение синтеза гормонов, что в свою очередь
приводит к формированию синдрома гипофизарной карликовости с
описанными выше признаками. Нарушение же нормального развития
гипофиза в данном случае вызывается мутацией гена.. Поэтому все
данные симптомы можно устранить гормональным лечением, но
добиться нормализации развития гипофиза не удается.
У человека ряд мутаций, вызывающих у гетерозигот формирова-
ние аномальных признаков, у гомозигот нередко обусловливает их
гибель. Например, доминантная мутация брахидактилии (коротко-
палости) у гетерозигот обусловливает недоразвитие фаланг и, как
следствие, укорочение пальцев, а у гомозигот — резкие аномалии
развития скелета и гибель в раннем возрасте.
В ряде случаев действие одного гена может спровоцировать из-
менение фенотипического выражения другого. Так, у дрозофилы до-
минантная мутация Notch, локализующаяся в ^-хромосоме и детер-
минирующая образование вырезки на крыльях, у гомозигот оказы-
вает летальное действие, которое усиливается под влиянием мутант-
ного гена short (укороченное тело) и ослабляется под действием
гена slight (маленькое тело).
'174

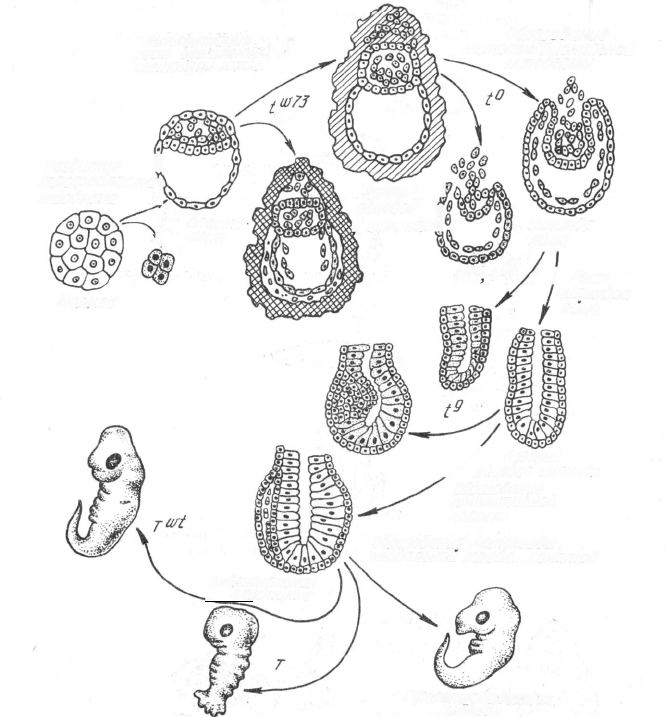
Четвериков Сергей Сергеевич
(1880—1959).
Механизмы генных взаимоот-
ношений на уровне конечных про-
дуктов их деятельности до сих пор
не выяснены. В настоящее время
предпринимаются попытки их ана-
лиза с помощью химерных орга-
низмов. Клеточные химеры полу-
чают путем соматической гибриди-
зации и клеточной инженерии, хи-
мерных животных — путем гибри-
дизации бластомеров. В 1961 г.
А. Тарковский попытался объеди-
нить делящиеся мышиные эмбрио-
ны, но развитие зародыша дальше
образования бластоцисты не по-
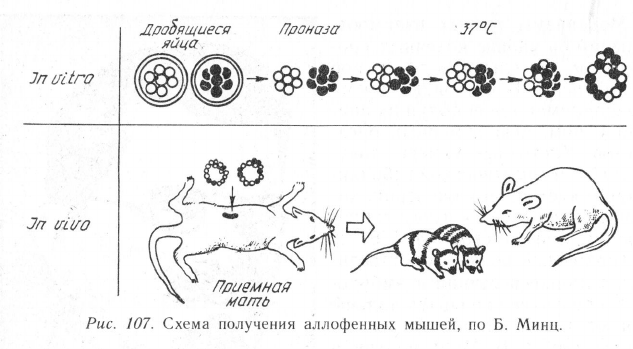
шло. В 1962 г. Б. Минц получила
первых химерных животных (ал-
лофенные, т. е. мозаичные, мыши)
методом конструирования бласту-
лы из бластомеров, взятых от раз-
ных животных (рис. 107). Эту
бластулу она имплантировала в
матку мыши-воспитательницы. По-
следняя становилась матерью мы-
шат нормальных размеров, но являющихся генетическими мозаиками
по разным тканям. Аллофенные мыши различались по 9 парам генов,
контролирующих окраску шкурки и радужки глаз, длину хвоста и
ушей, синтез фермента изоцитратдегидрогеназы. Все эти гены услов-
но подразделяются на гены автономного и неавтономного действия.
У первых доминантные аллели," проявляющиеся в одних клетках, не
подавляют их рецессивные аллели (даже если они находятся в гомо-
зиготном состоянии) в соседних клетках. Продукты же неавтономных
генов способны проникать в соседние клетки и оказывать влияние
на их фенотип.
Фенотип может изменяться и под влиянием факторов внешней
среды на зачатки различных органов, находящихся на определен-
ных, так называемых фенокритических, стадиях развития. Фено-
типически такие изменения напоминают мутации и называются
фенокопиями. К примеру, если цыплятам вводить инсулин в течение
первых трех дней эмбрионального развития, они рождаются бес-
хвостыми, если в возрасте пяти дней, у них остается недоразвитым,
укороченным, верхний клюв. Эти изменения представляют собой
фенокопии мутаций соответственно short upper beak и rumpless ness.
От условий внешней среды (температура, свет, состав пищи,
химические вещества, радиоактивные излучения и др.), а также от
характера взаимодействия генов нередко зависят степень Ьыражен-
ности признака (экспрессивность) и способность гена проявляться
в фенотипе (пенетрантность). К примеру, добавление в корм живот-
ных с мутацией карликовости гормона гипофиза экспрессивность
175
этого признака уменьшается, а при ранней и постоянной даче гормона
карликовость может вообще не развиться. В процессе развития
зародыша экспрессия генов может измениться и под влиянием поло-
вых гормонов. Так, ряд признаков (молочность скота, яйценоскость
кур, облысение у человека и др.) проявляются только у особей опреде-
ленного пола; противоположный пол может нести гены этих призна-
ков и передавать их потомству, но фенотипически они не проявляют-
ся. Примером изменения пенетрантности признака служит развитие
красной окраски цветка у примулы, что наблюдается только в том
случае, если растение выдерживается при низких температурах.
Недостаток витаминов нередко приводит к замедлению процессов
развития животных, а нарушение светового режима дня изменяет
направление и темпы развития растений.
Мутация не проявляется или не всегда бывает выражена также в
силу эпистатического действия других генов.
Следовательно, процесс формирования признаков и свойств орга-
низма представляет собой сложный путь реализации наследствен-
ной информации, проявляющейся лишь при определенном взаимо-
действии генотипа и факторов внешней среды.
Айала Ф., Кайгер Дж. Современная
генетика. М., 1988. Т.
2.
Алиханян С. И., Акифьев А. П., Чернин Л. С. Общая генетика. М., 1985.
Бердышев Г. Д., Криворучко И. Ф. Генетика человека
с
основами медицинской
генетики. Киев, 1978.
Гершензон С. М. Основы современной
генетики. Киев,
1979.
Гёрдон Дж. Регуляция функции генов в развитии
животных. М.,
1977.
Дубинин И. П.
Генетика. Кишинев,
1985.
Зуссман М. Биология
развития. М.,
1977.
Константинов А. В. Биология индивидуального
развития. Мн.,
1978.
Корочкин Л. И. Взаимодействие генов
в развитии. М.,
1977.
Кемп П., Арме К. Введение
в биологию. М.,
1988.
Ньют Д. Рост и развитие
животных. М.,
1973.
Скаврон С. Общая
биология. Варшава,
1968.
Хадорн Э., Венер Р. Общая
зоология. М.,
1989.
ЛИТЕРАТУРА
'176
#