Каминская Э.А. Общая генетика
Подождите немного. Документ загружается.

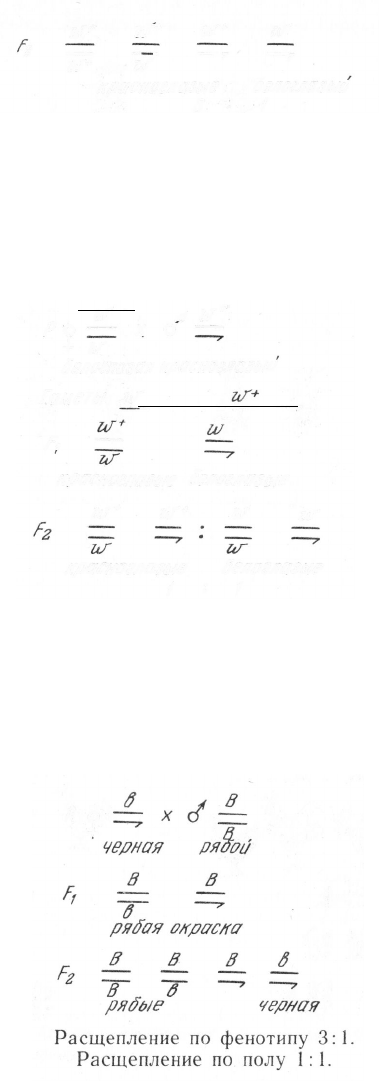
и* w* иг
2
TF ~иг
7 7
красноглазые бологлазыа
3 - /
В случае обратного скрещивания в потомстве первого поколе-
ния происходит перекрестное (крисс-кросс) наследование признака:
признак матери наследуют сыновья, а признак отца — дочери. Это
возможно только при условии, что ген, контролирующий развитие
признака, локализуется в половой хромосоме. Во втором поко-
лении расщепление по фенотипу составляет 1:1, что совпадает
с таковым по полу: половина самок и самцов имеют красные глаза,
половина — белые (рис. 96,6):
иГ У Ы+'
Р О X О —
4- is
белоглазая красноглазый
Гаметы тлГ_ 7
F
—
красноглазые белоглазые
и/
4
' иГ+ и иг
красноглазые белоглазые
i ; /
Примером наследования признаков, сцепленных с полом, при
гетерогаметности женского пола (рис. 97) служит наследование
гена в черной окраски оперения, локализующегося в Х-хромосоме
(доминантный аллель В контролирует рябую расцветку перьев)
у кур. Если гомогаметный пол (а здесь это самцы) несет в гомо-
зиготном состоянии доминантный признак, последний проявится у
всего потомства первого и трех четвертых второго поколений:
"2
'157
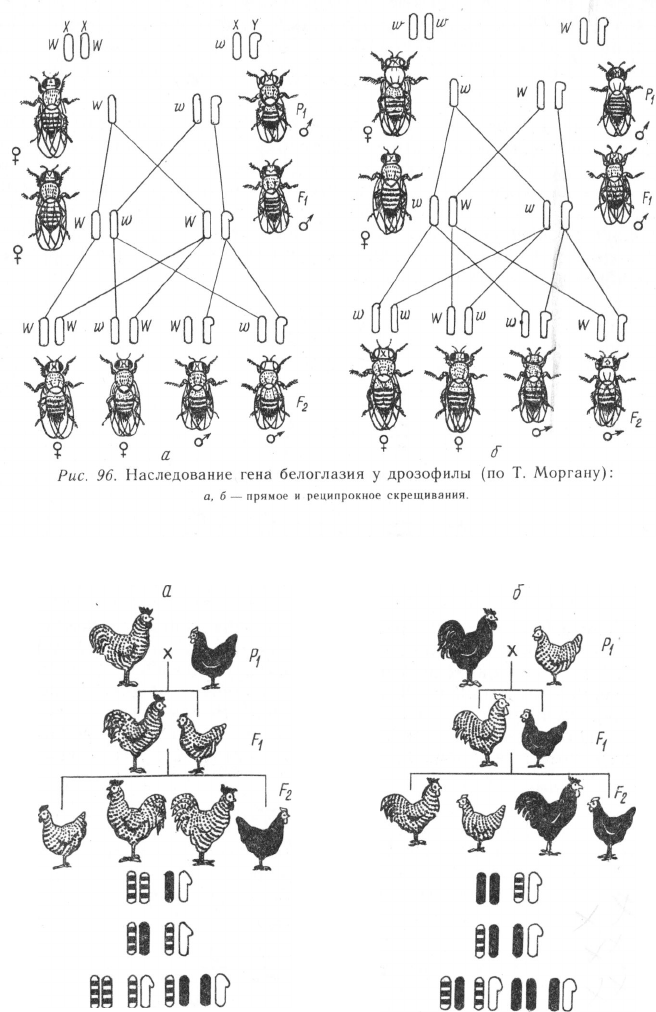
Рис. 97. Наследование пестрой окраски у кур (по Дж. Кру):
а, б —прямое и реципрокное скрещивания.
'158

При обратном скрещивании наблюдается перекрестное наследо-
вание признаков:
В j в
ря5ая черный
У человека известны признаки, гены которых локализуются
в Х-хромосоме. Среди них ряд доминантных мутаций: дефект эмали
зубов, вызвающий их потемнение; недостаточность фосфора в крови,
приводящая к развитию рахита, устойчивого к воздействию обычных
доз витамина Д; ферментативный дефект эритроцитов и др. Харак-
терным для этих заболеваний является то, что больной отец передает
аномалию только дочерям, а сыновья остаются здоровыми. В то же
время больная, гетерозиготная по данной аномалии женщи-
на, передает дефектный ген половине дочерей и половине сы-
новей.
Рецессивные, сцепленные с полом признаки у человека, такие,
как гемофилия (замедленная свертываемость крови), цветовая сле-
пота (невозможность различать цвета), проявляются лишь у мужчин,
тогда как носителями их являются женщины. Больной отец передает
аномальный ген только дочерям, но они остаются фенотипически
здоровыми, и лишь в потомстве у половины сыновей может проявить-
ся данное заболевание.
У большинства живых организмов К-хромосома инертна и не не-
сет активных генов. Однако у человека известны заболевания, гены
которых локализуются в F-хромосоме. Это ихтиоз (кожа уплотняется,
покрывается грубыми чешуями, щетинообразными утолщениями),
синдактилия (наличие перепонок между вторым и третьим пальцами
стопы) и др. Характерным в наследовании таких заболеваний являет-
ся то, что болезнь проявляется только у лиц мужского пола и пере-
дается от отца к сыну. Женщины не страдают этими болезнями и не
передают их детям. Такое наследование называется голандрическим.
ЛИТЕРАТУРА
Астауров Б. Л.
Генетика пола//Актуальные вопросы современной генетики. М., 1966.
Гершензон С. М. Основы современной
генетики. Киев,
1979.
Лобашев М. Е.
Генетика. Л.,
1967.
'159
Медведев И. Н. Беседы по биологии пола. Ми., 1976.
Мюнтцинг А. Генетика: Пер. с англ./Под ред. В. Н. Столетова. М., 1967.
Основы цитогенетики человека/Под ред. А. А. Прокофьевой-Бельговской. М., 1969.
Скаврон С. Общая биология. Варшава, 1968.
Глава 6. ГЕНЕТИЧЕСКИЕ ОСНОВЫ
ИНДИВИДУАЛЬНОГО РАЗВИТИЯ
Индивидуальное развитие живого орга-
низма, или онтогенез,— это развитие особи от момента слияния поло-
вых клеток до момента смерти. Нередко началом онтогенеза счи-
тается образование гамет у родительских особей.
У одноклеточных в процессе онтогенеза в период от одного де-
ления до другого формируются клеточные органеллы, происходят
определенные структурные изменения в ядерном аппарате и биохи-
мическом составе цитоплазмы. Но одноклеточные организмы прак-
тически «бессмертны», так как клетка не умирает, а лишь дает начало
двум новым, дочерним, клеткам.
У многоклеточных в процессе онтогенеза из одной клетки разви-
вается сложный организм с дифференцированными клетками и функ-
ционально специализированными тканями и органами. У них раз-
личают как бы два типа клеток: зародышевые, обеспечивающие
появление потомства (они «бессмертны») и соматические, не участ-
вующие в половом размножении (они погибают). Как зародышевые,
так и соматические клетки обладают тотипотентностью, т. е. содер-
жат полный запас генетической информации, необходимой для нор-
мального развития организма. В процессе развития происходит
дифференциация в экспрессии различных генов, что обусловливает
структурные и функциональные отличия разных тканей и органов и,
как следствие, различия между отдельными частями организма.
Процессы дифференцировки наиболее интенсивны на начальных
стадиях развития, затем они замедляются, но никогда не прекра-
щаются полностью.
Развитие в пределах яйцеклетки либо в утробе матери называется
эмбриональным или эмбриогенезом. В нем различают следующие
стадии: бластулы, гаструлы, органогенеза. На протяжении этих ран-
них этапов онтогенеза происходят накопление клеточного материала,
рост и дифференцировка клеток и тканей, формирование частей
зародыша.
В постэмбриональный период осуществляется дальнейшее фор-
мирование и изменение морфологических, физиологических и биохи-
мических признаков и свойств организма под влиянием внешней
среды, вырабатывается норма его поведения.
В процессе эмбрионального и постэмбрионального развития
происходит предусмотренная генетической программой гибель не-
которых типов дифференцированных клеток. Например, у эмбрионов
человека погибают клетки, соединяющие зачатки пальцев. Если этот
160
процесс нарушается, что может случиться вследствие мутации
гена, контролирующего гибель указанных клеток, пальцы останутся
соединенными перепонками (синдактилия).
Продолжительность жизни особей, т. е. онтогенез, также контро-
лируется генотипом. Это можно продемонстрировать с помощью так
называемого близнецового метода. У однояйцевых близнецов чело-
века, генетически тождественных, симптомы старения развиваются
почти одновременно, и они практически умирают в одно и то же
время, независимо от условий жизни.
У разных видов животных и растений продолжительность жизни
различная и служит видовым признаком. В процессе старения орга-
низма в клетках накапливаются ошибки репликации ДНК, пере-
стают выполнять свои функции системы репарации этих поврежде-
ний. Все это приводит к гибели клеток, уменьшению их количества
в органах и тканях, а затем к естественной гибели организма.
Реализация наследственной информации
Оплодотворенное яйцо является носителем генетической инфор-
мации материнского и отцовского организмов, и все индивидуальное
развитие сводится к ее реализации. Информация начинает реали-
зовываться еще в оогенезе, когда происходит транскрипция мате-
ринских генов. В результате в яйце формируются структуры, ко-
торые впоследствии определяют план дальнейшего развития (ось
симметрии, соединяющая вегетативный и анимальный полюсы
клетки; слоистое расположение питательных веществ; серый серп,
или организующий центр, определяющий билатеральность, и др.).
На стадии яйца гены нового организма не транскрибируются.
Это доказывается следующим экспериментом. В оплодотворенную
яйцеклетку вводили антибиотики (например, актиномицин), способ-
ные ингибировать транскрипцию. Но ее дробление продолжалось.
Это свидетельствовало о том, что в клетке, вероятно, сохранялся
некоторый запас синтезированной еще в ооците и-РНК, вполне доста-
точный для синтеза белка на ранних стадиях эмбриогенеза и исклю-
чающий необходимость в активной транскрипции.
Активность материнского генома находится под контролем цито-
плазмы яйцеклетки. Это можно установить путем либо изменения
цитоплазматического окружения ядра, либо пересадки ядер, либо
соматической гибридизации клеток. Например, установлено, что у на-
секомых направление развития яйцеклетки зависит от материнского
ядра, гены которого находятся в активном состоянии еще до оплодо-
творения. Это явление носит название предетерминация. Так,
вследствие мутации dor (deep orange) в яйцеклетках самки дрозо-
филы происходит блок развития. Но если в такую яйцеклетку доба-
вить цитоплазму нормальной яйцеклетки дрозофилы дикого типа,
блок устраняется. Следовательно, в цитоплазме нормальной яйце-
клетки содержатся какие-то вещества, подавляющие транскрипцию
ядерных генов. Это подтверждается также опытами, проведенными
на дрозофиле К. Ильмензее с сотрудниками в 1972 г. Они вводили
II. Зак. 5107
161
ядро из яйцеклетки, находящейся на стадии гаструлы, в неоплодотво-
ренную яйцеклетку, в которой собственное ядро было предварительно
разрушено воздействием ультрафиолетовых лучей. В результате
активность транскрипции введенных генов сразу же прекраща-
лась, начиналось дробление яйцеклетки и в дальнейшем ее раз-
витие шло нормально. Это свидетельствовало, с одной стороны,
о регулирующей роли цитоплазмы, с другой — о том, что ядро на
стадии гаструлы еще сохраняет все свои потенции.
Аналогичные результаты были получены и в опытах на амфибиях
по пересадке в ее яйцеклетку с разрушенным ядром ядра из диффе-
ренцированной клетки кишечного эпителия или кожи: активность
генов введенного ядра подавлялась, а ранее подавленные гены ак-
тивизировались.
Цитоплазматический контроль за развитием детерминируется
еще в неоплодотворенных яйцеклетках на стадии ооцита. Это под-
тверждается экспериментом Дж. Гёрдона (1970). Он осуществил
пересадку ядер из клеток почки шпорцевой лягушки в ооциты трито-
на, белки которого по составу отличаются от белков ооцитов лягуш-
ки. Через 3 дня в ооцитах тритона появились белки, характерные
для ооцитов лягушки, а гены, специфические для клеток почки лягуш-
ки, «выключились». Значит, ооциты содержат какие-то вещества,
способные «пробудить» ранее неработавшие гены и, следовательно,
яйцеклетки имеют их изначально (получают из ооцитов).
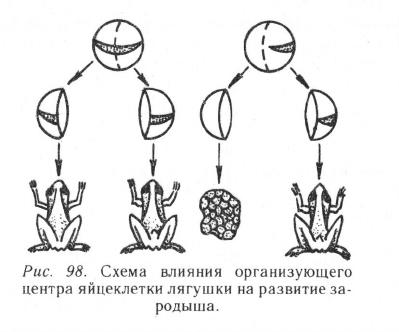
В цитоплазме яйцеклеток некоторых организмов (амфибии,
морские ежи) был обнаружен так называемый организующий центр.
Его химическая природа пока не установлена, но известны форма и
местоположение. Например, у лягушек он имеет форму серпа серо-
го или желтого цвета, у морских ежей — форму полоски. Установ-
лено, что яйца земноводных, содержащие организующий центр,
обычно развиваются нормально. Если же яйцеклетку на стадии
бластомера разделить на две части так, чтобы весь организующий
центр попал только в одну из них, то нормальный зародыш разовьет-
ся именно из нее, даже если обе части бластомера будут иметь нор-
мальные ядра (рис. 98). Очевидно, организующий центр каким-то
'162
образом регулирует направление развития зародыша, и при его от-
сутствии дифференцировка и дальнейшее развитие бластомера
прекращаются.
Таким образом, можно сделать вывод, что в развитии, особенно
на ранних стадиях, определенную регулирующую роль играет цито-
плазма яйцеклетки.
Взаимоотношение ядра и цитоплазмы
В период онтогенеза организм формируется под контролем на-
следственной информации, заложенной в ядре и цитоплазме. При-
чем ядерная информация, очевидно, несет основную ответствен-
ность за развитие.
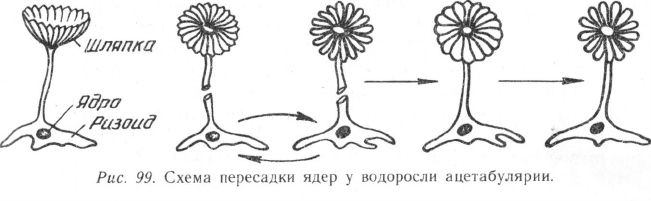
Оригинальным доказательством роли ядра в развитии являются
данные, полученные в серии опытов по пересадке ядер у морской од-
ноклеточной водоросли ацетабулярии. Очень крупная клетка ацета-
булярии (размер 0,2—10 см в высоту) дифференцирована на три
части: на одном ее конце имеется образование, напоминающее
по форме зонтик, на другом — ризоид («корешок»), соединенный с
зонтиком стебельком. В ризоиде располагается одно круглое ядро
диаметром 0,2 мм. Если у ацетабулярии удалить зонтик, он восста-
навливается, и это происходит сколько угодно раз. Если же у нее
предварительно отрезать ризоид с ядром, она продолжает жить, хотя
и теряет способность размножаться, но зонтик восстанавливается
только один раз. Если у двух разных видов ацетабулярии обменять
ризоиды с ядрами, т. е. произвести их пересадку, а затем удалить
зонтики, то они при регенерации будут приобретать форму, свойст-
венную форме зонтика той ацетабулярии, от которой было взято
ядро (рис. 99). Все это свидетельствует о том, что ядро принимает
участие в жизни клетки.
Однако до сих пор не совсем ясен механизм регуляции актив-
ности ядерных генов в процессе развития клетки. Тем не менее со-
вершенно очевидно, что не для всех этих процессов присутствие
ядра обязательно. В ряде случаев его роль как регуляторного меха-
низма несущественна, поскольку данную функцию, как уже отме-
чалось, выполняет цитоплазма: она определяет, когда и где должны
активизироваться те или иные ядерные гены либо генетическая ин-
формация, содержащаяся в цитоплазме в виде и-РНК.
Формирование признаков организма детерминируется и ядерны-
ми, и цитоплазматическими генами, но ведущая роль здесь при-
'163
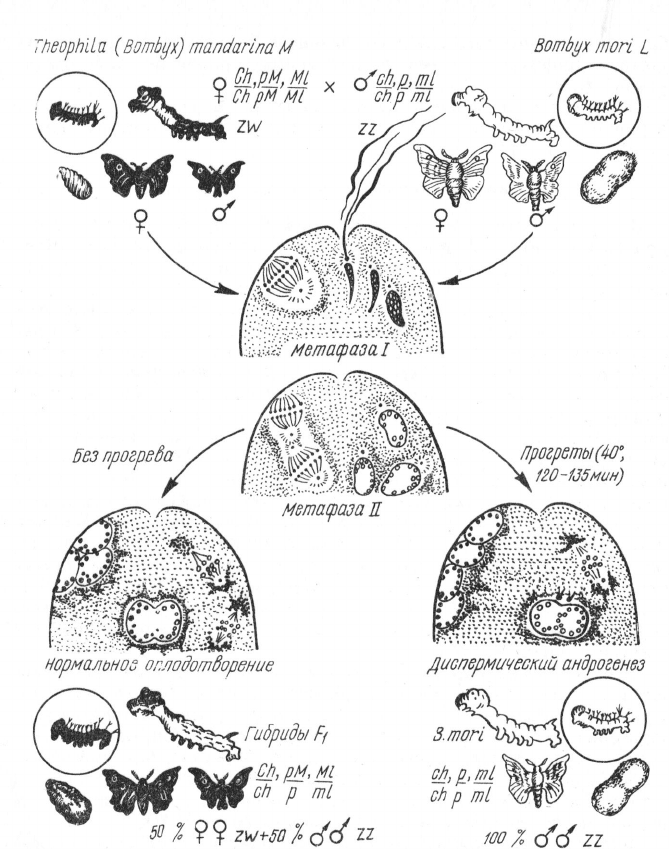
Рис. 100. Схема межвидового андрогенеза (по Б. JI. Астаурову, 1937).
надлежит ядру Убедительным доказательством этого является рабо-
та Б. Л. Астаурова (1957) по андрогенезу у тутового шелкопряда.
Астауров инактивировал нагреванием ядро яйцеклетки шелкопряда
Bombyx mori и оплодотворял ее спермиями самцов Bombyx man-
darina. При этом в ряде случаев два мужских пронуклеуса сливались
в диплоидное ядро, в результате чего все потомство получалось
нормальное диплоидное. Признаки его полностью повторяли отцов-
ские, и все оно оказывалось мужского пола (рис. 100).
На развитие организма могут также оказывать воздействие
'164

некоторые цитоплазматические
структуры (пластиды, митохонд-
рии, рибосомы). Известно, напри-
мер, влияние материнской цито-
плазмы у гибридов. Так, гибрид
лошади и осла обладает повышен-
ной выносливостью и жизнеспо-
собностью, большой физической
силой; реципрокное же скрещи-
вание не дает такого эффекта.
У растений по материнской линии
наследуется пестролистность в за-
висимости от характера распре-
деления дочерних клеток двух
типов пластид: зеленых, содержа-
щих хлорофилл, и белых, не спо-
собных к его образованию (см.
рис. 88). Примером такого насле-
дования служит и цитоплазмати-
ческая мужская стерильность у
растений, обусловленная цито-
плазматическим фактором стерильности цит
5
. Однако и здесь про-
слеживается контролирующая роль ядра: стерильность пыльцы у
растений развивается только в том случае, если фактор цит
5
соче-
тается с хромосомными генами rfrf (стерильность). Все это свиде-
тельствует о том, что в клетке устанавливаются определенные ядерно-
цитоплазматические взаимоотношения при ведущей роли ядра.
Астауров Борис Львович
(1904—1974).
Дифференциальная активность генов
На стадии дробления, когда формируется бластула, проявляет-
ся в основном митотическая активность клеток, которая регулирует-
ся за счет информации, содержащейся в яйце. При этом ни материн-
ский, ни отцовский геном не транскрибируются. По-видимому, во
время дробления активность генов направлена только на воспроиз-
ведение самих себя.
На стадии гаструляции происходят главным образом перемеще-
ния клеток относительно друг друга и изменение их формы. Однако
они еще не дифференцируются на клетки специфических тканей
взрослого организма. После определенного числа дроблений, когда
число клеток в зародыше достигает примерно 1000, в яйце начинается
синтез и-РНК. В конце периода бластулы начинает синтезировать-
ся т-РНК, а на стадии гаструлы — р-РНК.
В период органогенеза в основе дифференцировки органов и
тканей лежит дифференциальная активность генов и синтез специ-
фических белков. Именно в этот период функцию контроля над раз-
витием организма берет на себя генотип зародыша. С данного мо-
мента начинается реализация записанной в нем наследственной
информации. В этот период цитоплазматический контроль за раз-
165
витием заменяется ядерным. Однако, как из одной клетки с одним
набором хромосом формируются специфические строго дифферен-
цированные клетки, каков путь от гена до признака, до сих пор
окончательно не выяснено. У прокариот этот процесс более изучен.
Установлено, что у них гены контролируют развитие признаков,
а активность последних регулируется соответствующими процес-
сами. У эукариот все выглядит гораздо сложней. У человека, на-
пример, клеточный цикл занимает 24 часа, а весь геном транскри-
бируется в течение 1000 дней. Это дает основание предполагать,
что в клетках работает какая-то активно выбирающая система ре-
гуляции, контролирующая транскрипцию информации с определен-
ных генов. Можно было бы предположить, что дифференциальная
активность генов в разных клетках обусловливается не только на-
личием системы регуляции, но и утратой специализированными
клетками ряда генов при сохранности тех из них, которые контро-
лируют синтез белков, специфических для данного органа или ткани.
Однако это опровергается результатами экспериментальных иссле-
дований. Так, Дж. Гёрдон (1960), используя методику, разработан-
ную Р. Бриггсом и Т. Кингом, ядра неоплодотворенных яйцеклеток
темно-зеленой шпорцевой лягушки заменил ядрами клеток кишечного
эпителия лягушек-альбиносов. Все родившиеся головастики оказа-
лись альбиносами (рис. 101). Это свидетельствует о том, что ядра
дифференцированных соматических клеток лягушки содержат пол-
ный набор генетической информации. Следовательно, при нормаль-
ном развитии в процессе дифференцировки гены не утрачиваются,
а лишь теряют активность; но при изменении условий она может
восстановиться.
К аналогичному выводу пришел и Э. Хадорн (1968), проделав-
ший опыты по пересадке имагинальных дисков, формирующихся у
личинок дрозофилы из имагинальных клеток, которые, в отличие
от клеток тела личинки, делятся, в результате чего их количество
по мере ее развития возрастает. На стадии куколки эти клетки претер-
певают дифференцировку и дают начало различным органам взрос-
лой мухи (рис. 102). Хадорн пересадил из зрелой личинки в молодую
имагинальный диск, из которого в обычных условиях образуются
половые протоки. После этого диски в процессе метаморфоза ли-
чинки в некоторых случаях давали не гениталии, а антенны или
аристы.
Следовательно, клеточная дифференцировка обусловливается не
утратой тех или иных генов, а их избирательной репрессией и де-
репрессией, определяющими последовательность стадий развития.
Это означает, что в процессе развития клетки или особи происходит
как бы переключение транскрипции с одних генов на другие на фоне
общей согласованной регуляции функции всех генов. Дифферен-
циальное включение в активную транскрипцию разных генов на
различных этапах эмбриогенеза можно продемонстрировать на
примере серии рецессивных аллелей по гену Т у мышей: эмбрионы,
гомозиготные по той или иной аллели, достигают соответствующей
стадии развития и погибают, не завершив его полностью (рис. 103).
166