Глебов Ф.З. Взаимоотношения леса и болота в таежной зоне
Подождите немного. Документ загружается.

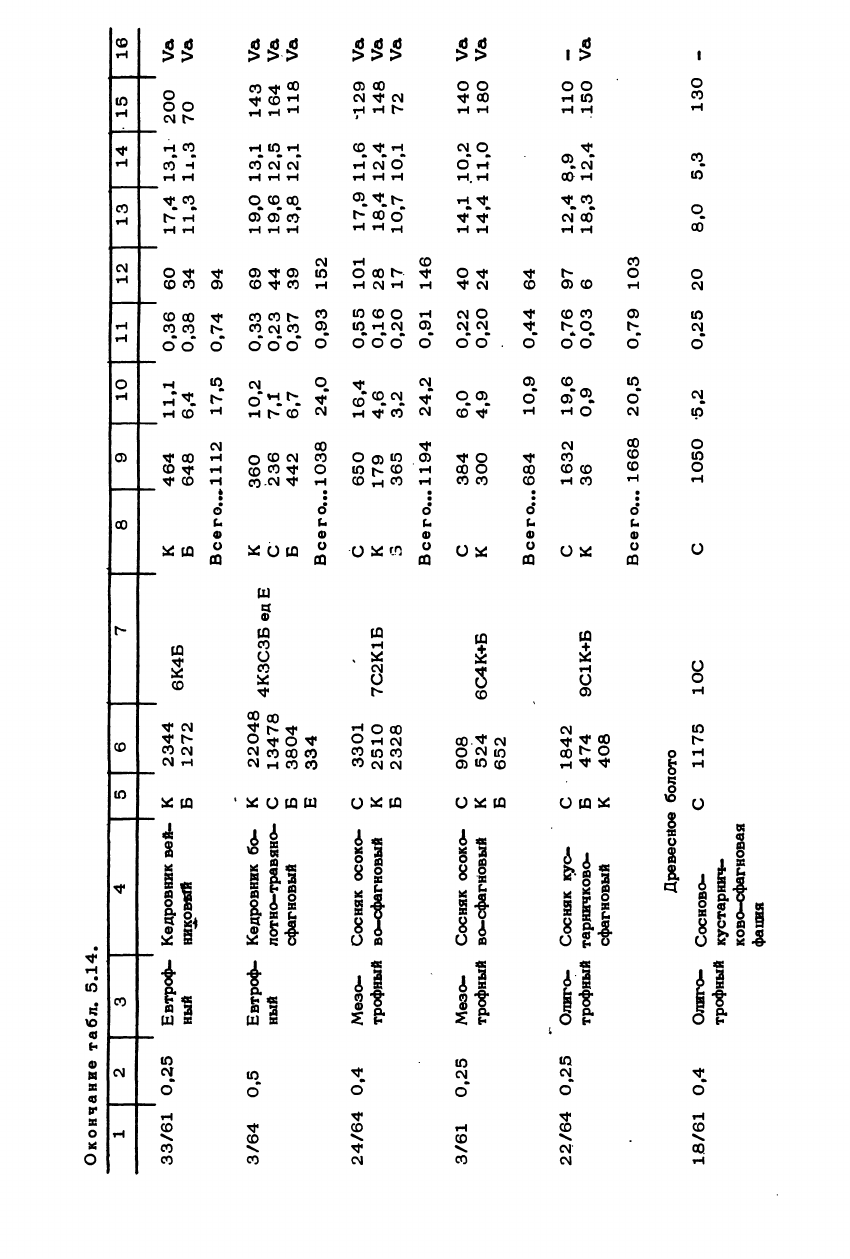
Окончание
табл.
5.14.
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16
33/61 0,25
Евтроф-
Кедровник
вей-
К 2344 К 464 11,1 0,36 60 17,4 13,1 200 Va
ный
никовет
Б 1272 Б 648 6,4 0,38 34 11,3 1±,3 70 Va
Всего..Л112 17,5 0,74 94
3/64 0,5
Евтроф-
Кедровник
бо- К 22048
4К
о
Г
о
К
р
к 36
°
10
»
2
°»
33
69 19
»°
13
»
1 143 Va
ный
лотно-травяно-
С 13478
4ISk
^
Udb
ед ь
с
i236
7,1 0,23 44 19,6 12,5 164 Va
сфагновый
Б 3804 Б 442 6,7 0,37 39 13,8 12,1 118 Va
Е 334
Всего...1038 24,0 0,93 152
24/64
0,4
Мезо-
Сосняк
осоко-
С 3301 С 650 16,4 0,55 101 17,9 11,6 '129 Va
трофный
во-сфагновый
К 2510
7С2К1Б
К 179 4,6 0,16 28 18,4 12,4 148 Va
Б
2328 3 365 3,2 0,20 17 10,7 10,1 72 Va
Всего...
1194 24,2 0,91 146
3/61 0,25
Мезо-
Сосняк
осоко-
С 908 С 384 6,0 0,22 40 14Д .10,2 140 Va
трофный
во-сфагновый
К 524
6С4К+Б
К 300 4,9 0,20 24 14,4 11,0 180 Va
Б
652
Всего...
684 10,9 0,44 64
22/64 0,25
Олиго- Сосняк
кус- С 1842 С 1632 19,6 0,76 97 12,4 8,9 110 -
трофный
тарничково-
Б 474
9С1К+Б
К 36 0,9 0,03 6 18,3 12,4 150 Va
сфагновый
К 408
Всего...
1668 20,5 0,79 103
Древесное болото
18/61 0,4
Олиго- Сосново-
С 1175 ЮС С 1050 5,2 0,25 20 8,0 5,3 130 -
трофный
кустарнич-
ково-сфагновая
фация

ном болоте свидетельствует о его большей стабильности, чем в заболоченном
лесу* Отсюда в болотном лесу и на древесном болоте более высокая, чем в
заболоченном лесу, степень разновозрастности /Колесников, 1947; Прозоров,
1961;
Комин, 1967; Пятецкий, Медведева, 1967; Глебов, 1969/, а с этим
связан особый характер кривых распределения количества деревьев по возра-
сту, толщине и высоте, при котором максимальное число стволов приходится
на минимальные ступени, а затем, по мере увеличения ступеней происходит
его падение (рис. 5.5-5.6). Кроме того, такое различие обусловлено мень-
шей подверженностью древостоев пожарам с увеличением гидроморфизма. В
заболоченном лесу разновозрастность выражена слабее.
Процесс превращения заболоченного леса в первичный болотный лес в
условиях различной трофности идет по-разному /Глебов, 1970/. Рассмотрим
два крайних типа заболачивания.
Первый - это олиготрофное заболачивание атмосферными водами, мягки-
ми ненапорными грунтовыми водами и верховодками, а также подтопление во-
дами наступающих верховых болотных массивов. В этом типе заболачивание
проходит два этапа - экзогенный и эндогенный.
На экзогенном, свойственном заболоченному лесу, основными пионерами
заболачивания произрастающих в олиготрофных условиях лишайниковых и ли-
шай никово-зеленомошных боров являются сфагновые мхи. На границе с болот-
ными массивами на низко расположенных участках, где почвенно-грунтовые
воды выходят к самой поверхности, заболачивание начинается с выклинивания
со стороны болота мочажин, заросших гигрофильными мхами - Sphagnum
obtusum, S* majus, S. jensehnii, S. balticum и др. Если поверхность поч-
вы относительно приподнята, то заболачивание осуществляется наползанием
вала из мезогигрофильных видов - S. angustifolium, S. Russowii, S. ne-
moreum и в отдельных случаях - S. rubellum, S. magelianicum и даже
S Ли scum. Высота такого вала 40-50 см, ширина может достигать 20 см.
При еще большей приподнятости почвенной поверхности, а также с удалением
от болота вал расчленяется на отдельные сфагновые подушки (наиболее рас-
пространенный вариант), первоначально возникающие в микропонижениях. В
результате создается столь характерный для сосняков лишайниково-сфагновых
и, в меньшей степени, кустарничково-зеленомошно-сфагновых мозаичный на-
почвенный покров. При достаточном расчленении микрорельефа почвенной по-,
верхности выклинивание мочажин со стороны болота в микропонижениях мо-
жет сопровождаться образованием сфагновых подушек на макроповышениях.
Одновременно происходит поселение на сфагнах гидрофитов (преимущественно
Сагех globularis и кустарнички), увеличивается мощность лесной подстил-
ки и идет почвенный процесс образования иллювиального горизонта, мешаю-
щего просачиванию воды и усиливающего застойность. До определенного мо-
мента, пока основным компонентом системы являются лабильные живые ор-
ганизмы, описываемый процесс легко обратим за счет климатических флук-
туации. В сухие годы, когда средний уровень воды понижается, накопившаяся
подстилка разлагается, чему способствуют пожары, свойственные сухим пери-
одам,
и гидрофиты сменяются мезофитами. По-видимому, разрушается и поч-
венный иллювиальный горизонт. Продолжительность таких циклов, фиксируемая
анализом особенностей развития древостоев, не больше 200-300 лет.
Заболачивание лесов этого типа чрезвычайно широко распространено в
таежной зоне, особенно в ее северной части, и издавна привлекает внимание.
Годичный прирост по высоте сфагновых мхов достигает 10 см /Тюремнов,
1949/.
Соответственно очень велика скорость заболачивания суходолов и со-
ставляет несколько метров в год: Р.П.Спарро /1924/ - 11 м/год, Ю.А.Льво-
ва /1976/ - 1 м/год.
102
лг,%
гол
мл
п—i—I—i—i—i—I—i—г
12 20 23 JS 44М,СМ
А А
10 50 90 130 170 210 250 А^ет
5о\
40Л
30
гол
ю\
2\
\
\
X
\\
\
\
т—I—I—I—I—I—I—I—г
/р
^
50^ 30 130 170 ^
210 Ажт
4 12 20 25 3SM.CM
—п—i—i—i—ill
i—i—л—V
s
*
1 5 9 13 17 21 25М,М
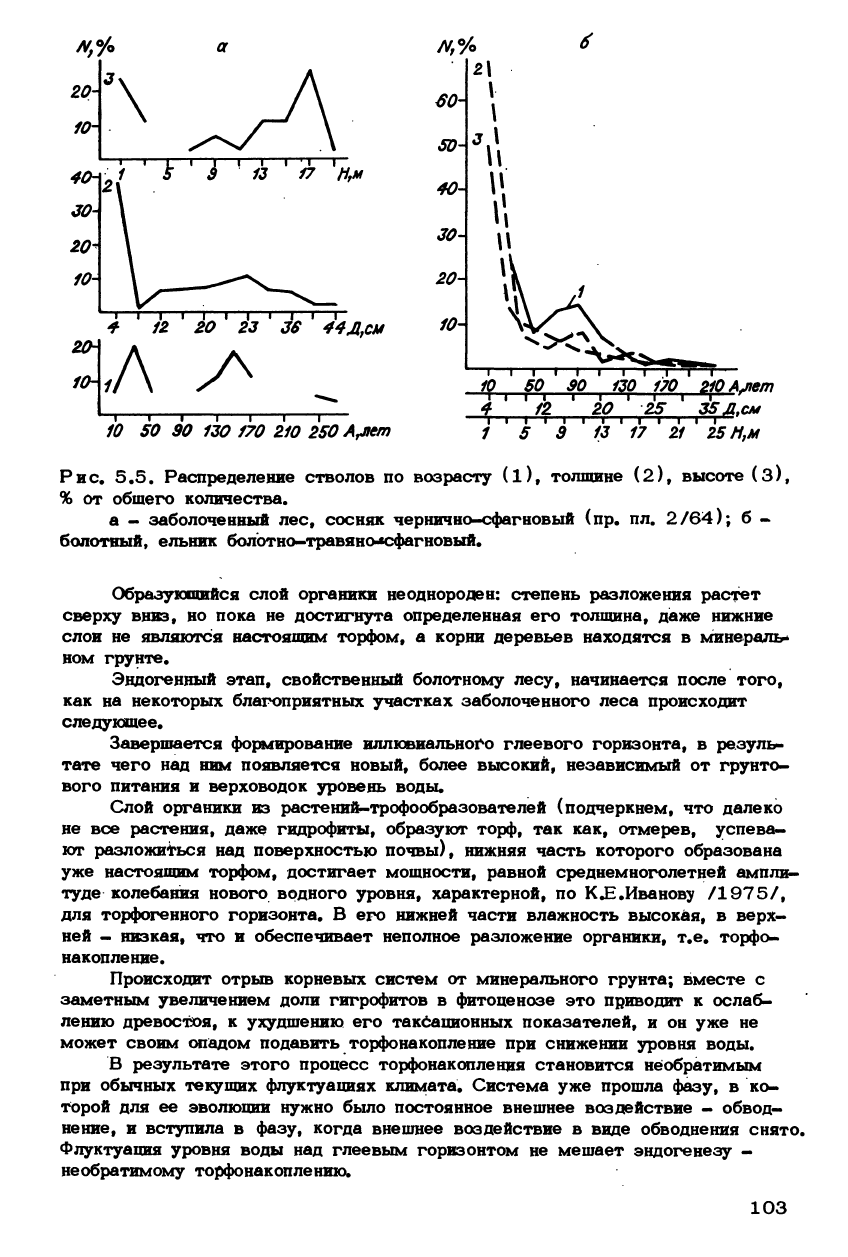
Рис. 5.5. Распределение стволов по возрасту (l)
f
толщине (2), высоте (3),
% от общего количества.
а - заболоченный лес, сосняк чернично-сфагновый (пр. пл. 2/64); б -
болотный, ельник болотно-травяно-*сфагновый.
Образующийся слой органики неоднороден: степень разложения растет
сверху вниз, но пока не достигнута определенная его толщина, даже нижние
слои не являются настоящим торфом, а корни деревьев находятся в минераль-
ном грунте.
Эндогенный этап, свойственный болотному лесу, начинается после того,
как на некоторых благоприятных участках заболоченного леса происходит
следующее.
Завершается формирование иллювиального глеевого горизонта, в резуль-
тате чего над ним появляется новый, более высокий, независимый от грунто-
вого питания и верховодок уровень воды.
Слой органики из растений-трофообразователей (подчеркнем, что далеко
не все растения, даже гидрофиты, образуют торф, так как, отмерев, успева-
ют разложиться над поверхностью почвы), нижняя часть которого образована
уже настоящим торфом, достигает мощности, равной среднемноголетней ампли-
туде колебания нового водного уровня, характерной, по К .Е.Иванову /1975/,
для торфогенного горизонта. В его нижней части влажность высокая, в верх-
ней - низкая, что и обеспечивает неполное разложение органики, т.е. торфо-
накопление.
Происходит отрыв корневых систем от минерального грунта; вместе с
заметным увеличением доли гигрофитов в фитоценозе это приводит к ослаб-
лению древостоя, к ухудшению его таксационных показателей, и он уже не
может своим опадом подавить торфонакопление при снижении уровня воды.
В результате этого процесс торфонакопления становится необратимым
при обычных текущих флуктуациях климата. Система уже прошла фазу, в ко-
торой для ее эволюции нужно было постоянное внешнее воздействие - обвод-
нение, и вступила в фазу, когда внешнее воздействие в виде обводнения снято.
Флуктуация уровня воды над глеевым горизонтом не мешает эндогенезу -
необратимому торфонакоплению.
103
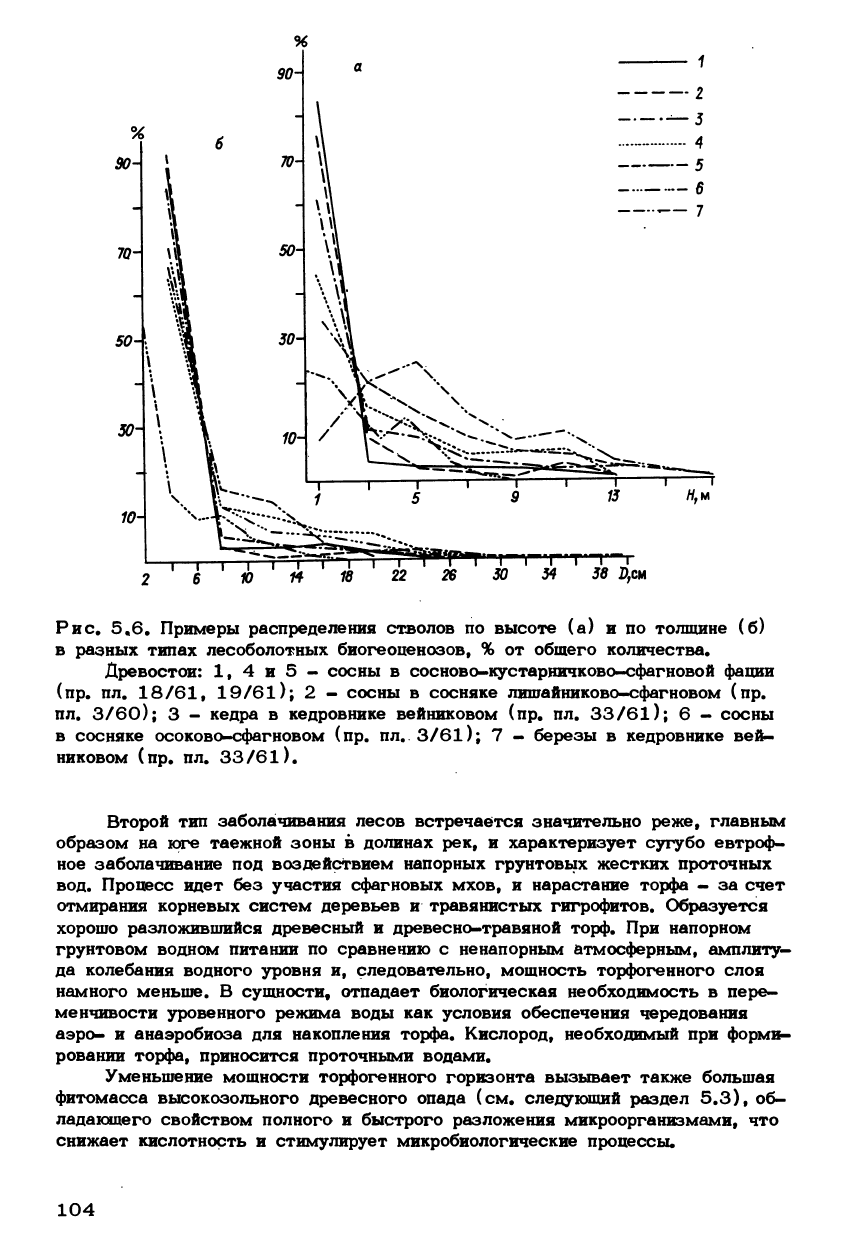
Рис. 5.6. Примеры распределения стволов по высоте (а) и по толщине (б)
в разных типах лесоболотных биогеоценозов, % от общего количества.
Древостой: 1, 4и5- сосны в сосново-кустарничково-сфагновой фации
(пр.
пл.
18/61,
19/61); 2 - сосны в сосняке лишайниково-сфагновом (пр.
пл.
3/60); 3 - кедра в кедровнике вейниковом (пр. пл. 33/61); 6 - сосны
в сосняке осоково-сфагновом (пр. пл. 3/61); 7 - березы в кедровнике вей-
никовом (пр. пл. 33/61).
Второй тип заболачивания лесов встречается значительно реже, главным
образом на юге таежной зоны в долинах рек, и характеризует сугубо евтроф-
ное заболачивание под воздействием напорных грунтовых жестких проточных
вод.
Процесс идет без участия сфагновых мхов, и нарастание торфа - за счет
отмирания корневых систем деревьев и травянистых гигрофитов. Образуется
хорошо разложившийся древесный и древесно-травяной торф. При напорном
грунтовом водном питании по сравнению с не напорным атмосферным, амплиту-
да колебания водного уровня и, следовательно, мощность торфогенного слоя
намного меньше. В сущности, отпадает биологическая необходимость в пере-
менчивости уровеиного режима воды как условия обеспечения чередования
аэро-
и анаэробиоза для накопления торфа. Кислород, необходимый при форми-
ровании торфа, приносится проточными водами.
Уменьшение мощности торфогенного горизонта вызывает также большая
фитомасса высокозольного древесного опада (см. следующий раздел 5.3), об-
ладающего свойством полного и быстрого разложения микроорганизмами, что
снижает кислотность и стимулирует микробиологические процессы.
104

Наши исследования /Глебов, Александрова, 1973/ показали, что в оли-
готрофном (омбротрофном) болотном лесу с сильнопеременчивым у решенным
режимом вод и развитым торфогенным горизонтом наиболее насыщен аэроб-
ными микроорганизмами верхний слой торфа в микроповышениях рельефа, тог»
да как в евтрофном лесу напорного грунтового питания, где переменчивость
водного уровня и такой горизонт почти не выражены, наиболее богат такими
микроорганизмами слой торфа в обводненных микропонижениях.
Как и при олиготрофном типе заболачивания, в почве происходит форми-
рование глеевого горизонта, но напор воды препятствует сплошному оглее-
нию.
Образование заболоченного, а затем болотного леса все время идет под
воздействием этого напора. Разделить заболачивание на экзо- и эндогенную
стадии нельзя. Между этими двумя крайними типами заболачивания - олиго-
трофным, сопровождаемым непременным формированием торфогеиного горизон-
та,
и евтрофным, при котором этот горизонт не образуется, - находится ряд
промежуточных типов, отличающихся трофностью условий его формирования и
соответственно мощностью.
Прирост торфа есть» результат двух разнонаправленных процессов - на-
копления и разложения отмершей фитомассы. При экзогенезе оба процесса
идут быстро и скачкообразно, заболоченная территория может полностью раз-
болотиться. (Напомним, что Н.И. Пьявченко считал, что свыше 70% заболо-
ченной площади составляют временно переувлажненные леса). При эндогенезе
заболачивание медленное, постепенное и неуклонное; торфонакопление может
приостанавливаться, но не прекращаться (при обычных текущих флуктуациях
климата). Поэтому определить время, необходимое для формирования торфо-
генного горизонта, можно лишь приблизительно, приняв, что оно обусловлено
средней скоростью торфонакопления в голоцене - 0,5мм/год (см. разд. 6.2.3).
Минимальная толщина торфоге«ного горизонта в болотном лесу 40 см (кор-
невые системы деревьев уже вышли из минерального грунта и находятся в
торфе). Отсюда следует, что примерное время формирования горизонта 800 лет.
За такое же время должен сформироваться глеевый горизонт. Уклоны почвен-
ной поверхности в заболоченном и болотных лесах 0,01-0,02 /Иванов, 1975/.
Простейший расчет показывает, что при скорости торфонакопления 0,5 мм/год
скорость заторфовывания. таких уклонов в автоморфных лесах под воздействи-
ем наступления соседних болот 3-6 см/год, т.е. при эндогенезе горизонталь-
ная скорость заболачивания на два порядка ниже, чем при экзогенезе (см/год
против м/год). За 800 лет граница болотного леса продвинется всего на
25-50 м.
В олиготрофных условиях лесная обстановка сохраняется недолго. Сразу
же после смыкания мочажин и сфагновых подушек произрастающий до забо-
лачивания древостой выпадает и формируется сосново-кустарничково-сфагновая
фация, в которой деревья уже не являются эдификаторами. Таким образом,
первичные болотные, леса здесь не развиваются, а в заболоченных лесах - сос-
няках лишайниково-сфагновом и кустарничково-зеленомошно-сфагновом - вслед-
ствие быстроты заболачивания древостой состоит из двух поколений. Одно из
них, усыхающее и разреженное, является остатком древостоя, бывшего зЪесь
до заболачивания, другое - возникло после заболачивания. Деревья этих поко-
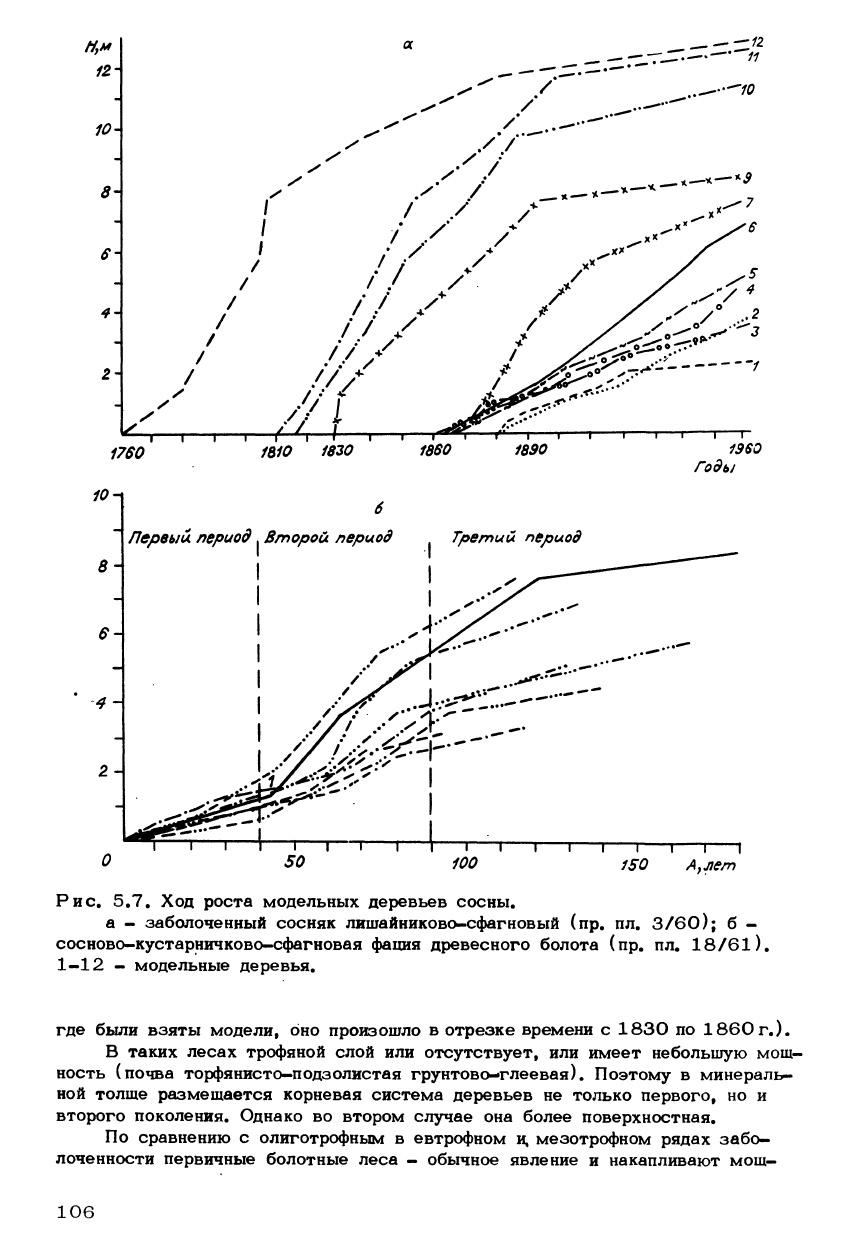
лений весьма отличны по характеру хода роста (рис* 5.7, а). Для деревьев
первого поколения (модели 7, 9-12) характерны относительно энергичный
рост в первую половину жизни, протекающую до заболачивания, и снижение
его во второй. Деревья второго поколения (модели 1-6) с начала существо-
вания растут намного медленней, чем в этом возрасте росли деревья первого
поколения. Интересно отметить, что анализы хода роста моделей дают возмож-
ность устанавливать время заболачивания. (Например, на пробной площади,
105
—
— -~.^
11
8А
еЛ
*
Л
24
Первый период
Второй период Третий период
/У
Л**??—
•'ШР'Г"
—г—
50
/00
т г
/50
П 1 1
А,
лет
Рис. 5.7. Ход роста модельных деревьев сосны.
а - заболоченный сосняк лишайниково-сфагновый (пр. пл. 3/60); б -
сосново-кустарничково-сфагновая фация древесного болота (пр. пл. 18/61),
1-12 - модельные деревья.
где были взяты модели, оно произошло в отрезке времени с 1830 по 1860г.).
В таких лесах трофяной слой или отсутствует, или имеет небольшую мощ-
ность (почва торфянисто-подзолистая грунтово-глеевая). Поэтому в минераль-
ной толще размещается корневая система деревьев не только первого, но и
второго поколения. Однако во втором случае она более поверхностная.
По сравнению с олиготрофным в евтрофном и» мезотрофном рядах забо-
лоченности первичные болотные леса - обычное явление и накапливают мощ-
106
0 SO 100 150 200 250А,ле/п
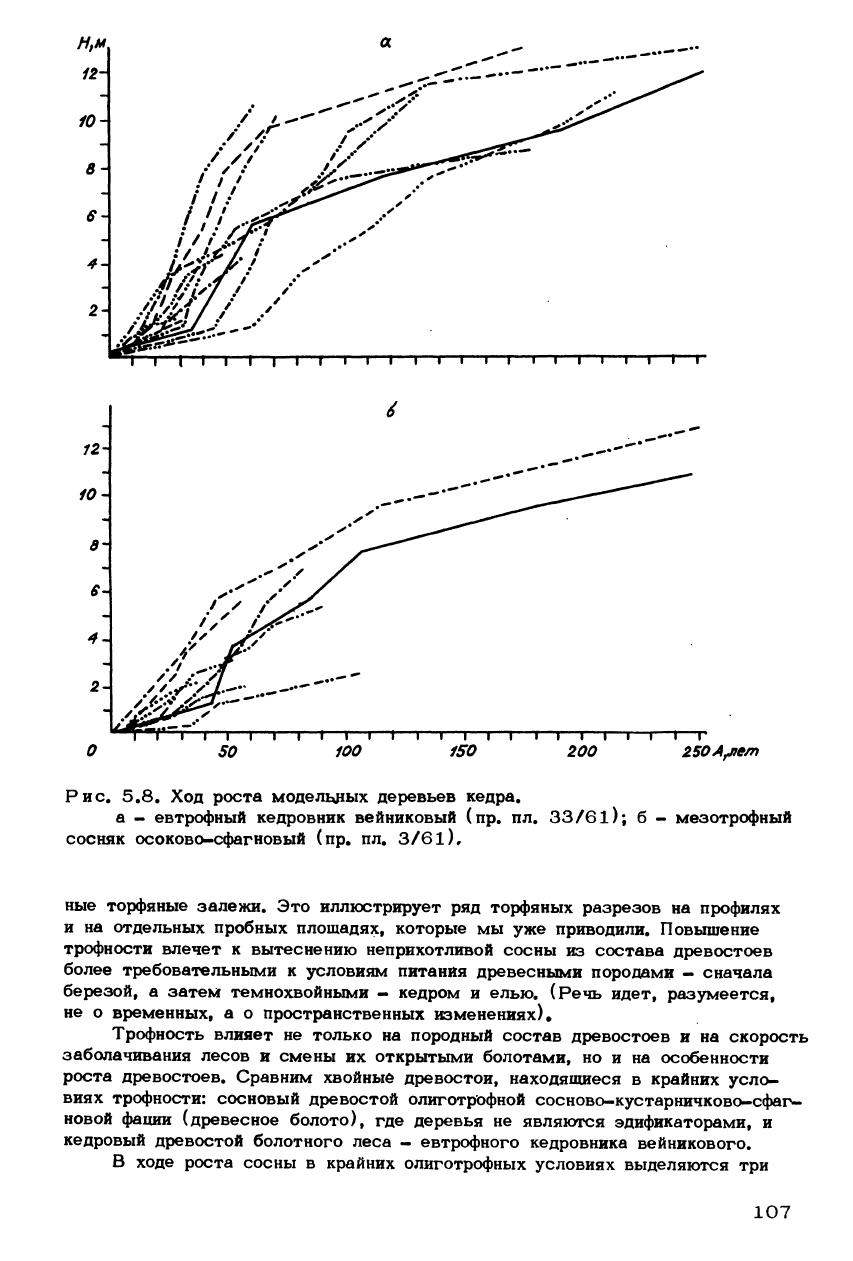
Рис. 5.8. Ход роста модельных деревьев кедра.
а - евтрофный кедровник вейниковый (пр. пл. 33/61); б - мезотрофный
сосняк осоково-сфагновый (пр. пл. 3/61),
ные торфяные залежи. Это иллюстрирует ряд торфяных разрезов на профилях
и на отдельных пробных площадях, которые мы уже приводили. Повышение
трофности влечет к вытеснению неприхотливой сосны из состава древостоев
более требовательными к условиям питания древесными породами - сначала
березой, а затем темнохвойными - кедром и елью. (Речь идет, разумеется,
не о временных, а о пространственных изменениях) •
Трофность влияет не только на породный состав древостоев и на скорость
заболачивания лесов и смены их открытыми болотами, но и на особенности
роста древостоев. Сравним хвойные древостой, находящиеся в крайних усло-
виях трофности: сосновый древостой олиготрофной сосново-кустарничково-сфаг—
новой фации (древесное болото), где деревья не являются эдификаторами, и
кедровый древостой болотного леса - евтрофного кедровника вейникового.
В ходе роста сосны в крайних олиготрофных условиях выделяются три
107
—I—I—I—I—I—I
-
200
250А,лет
16-
14-
12-
/0-
8-
e-
4-
2-
0
И
ijjf
T i—i—i—i—i—i—i—i—r-T—i -r-
IIIIIIIIII
50 100 150 200
i
i i i i
250
300
А
9
ле/п
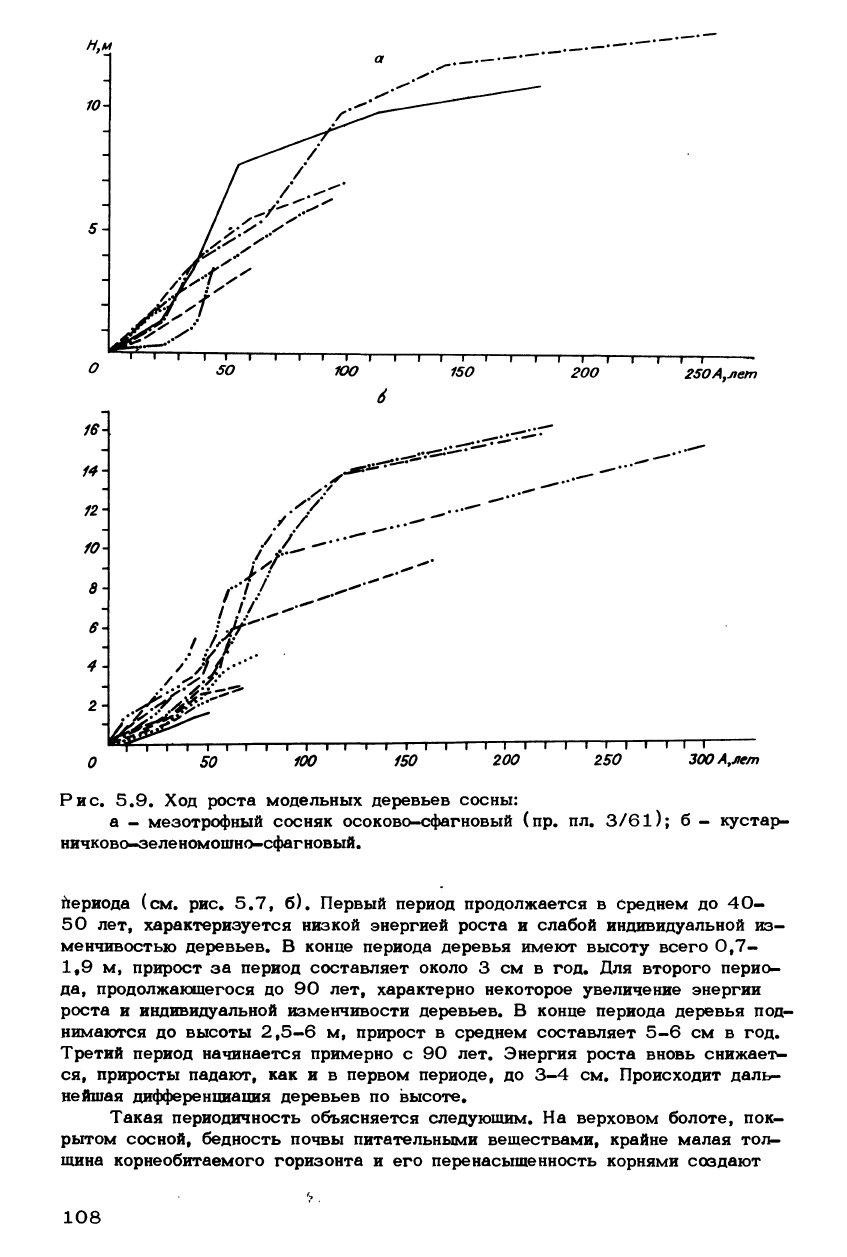
Рис. 5.9. Ход роста модельных деревьев сосны:
а - мезотрофный сосняк осоково-сфагновый (пр. пл. 3/61); б - кустар-
ничково-зе ле номошно-сфаг новый •
периода (см. рис, 5.7, б). Первый период продолжается в Среднем до 40-
50 лет, характеризуется низкой энергией роста и слабой индивидуальной из-
менчивостью деревьев. В конце периода деревья имеют высоту всего 0,7-
1,9 м, прирост за период составляет около 3 см в год. Для второго перио-
да,
продолжающегося до 90 лет, характерно некоторое увеличение энергии
роста и индивидуальной изменчивости деревьев. В конце периода деревья под-
нимаются до высоты 2,5-6 м, прирост в среднем составляет 5-6 см в год.
Третий период начинается примерно с 90 лет. Энергия роста вновь снижает-
ся,
приросты падают, как и в первом периоде, до 3-4 см. Происходит даль-
нейшая дифференциация деревьев по высоте.
Такая периодичность объясняется следующим. На верховом болоте, пок-
рытом сосной, бедность почвы питательными веществами, крайне малая тол-
щина корнеобитаемого горизонта и его перенасыщенность корнями создают
108
/
/
"I
1 1 1 1 1
10 20 30 40 SO SO
А,лет
-i 1 1 1 1 1 1 1 1 1
0 10 20 30 40 SO SO 70 80 SO
fOO
А,лет
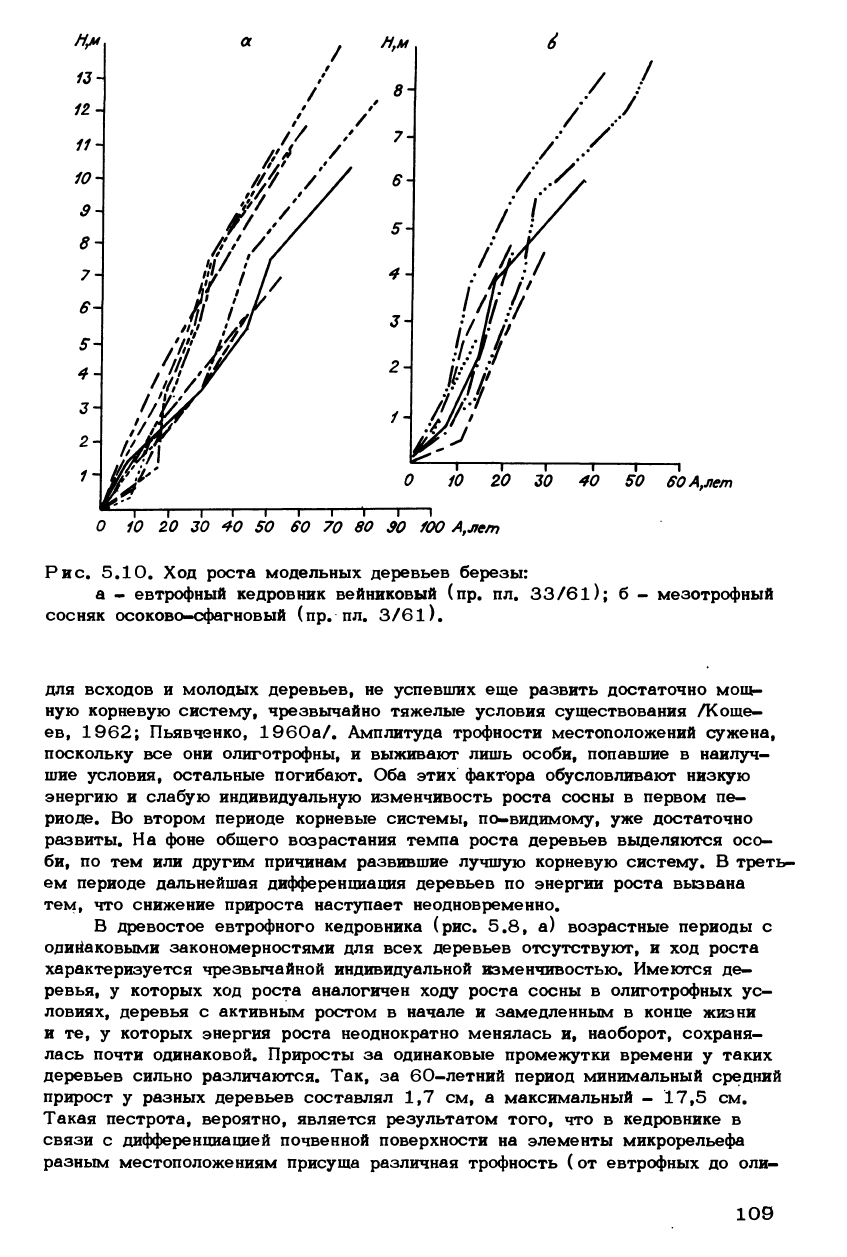
Рис. 5.10. Ход роста модельных деревьев березы:
а - евтрофный кедровник вейниковый (пр. пл. 33/61); б - мезотрофный
сосняк осоково-сфагновый (пр. пл. 3/61).
для всходов и молодых деревьев, не успевших еще развить достаточно мощ-
ную корневую систему, чрезвычайно тяжелые условия существования /Коще-
ев,
1962; Пьявченко, 1960а/. Амплитуда трофности местоположений сужена,
поскольку все они олиготрофны, и выживают лишь особи, попавшие в наилуч-
шие условия, остальные погибают. Оба этих фактора обусловливают низкую
энергию и слабую индивидуальную изменчивость роста сосны в первом пе-
риоде. Во втором периоде корневые системы, по-видимому, уже достаточно
развиты. На фоне общего возрастания темпа роста деревьев выделяются осо-
би,
по тем или другим причинам развившие лучшую корневую систему. В треть-
ем периоде дальнейшая дифференциация деревьев по энергии роста вызвана
тем,
что снижение прироста наступает неодновременно.
В древостое евтрофного кедровника (рис. 5.8, а) возрастные периоды с
одинаковыми закономерностями для всех деревьев отсутствуют, и ход роста
характеризуется чрезвычайной индивидуальной изменчивостью. Имеются де-
ревья, у которых ход роста аналогичен ходу роста сосны в олиготрофных ус-
ловиях, деревья с активным ростом в начале и замедленным в конце жизни
и те, у которых энергия роста неоднократно менялась и, наоборот, сохраня-
лась почти одинаковой. Приросты за одинаковые промежутки времени у таких
деревьев сильно различаются. Так, за 60-летний период минимальный средний
прирост у разных деревьев составлял 1,7 см, а максимальный - 17,5 см.
Такая пестрота, вероятно, является результатом того, что в кедровнике в
связи с дифференциацией почвенной поверхности на элементы микрорельефа
разным местоположениям присуща различная трофность (от евтрофных до оли-
109

готрофных включительно). В этом - одно из существенных отличий евтрофных
условий среды от олиготрофных.
Ход роста деревьев кедра (рис. 5.8, б) и сосны (рис. 5.9, а) в мезо-
трофном сосняке осоково-сфагновом носит черты промежуточного характера
между описанными выше. С одной стороны, здесь, как и в евтрофном древо-
стое, отсутствуют возрастные периоды, характеризующиеся одинаковыми за-
кономерностями для всех деревьев, а с другой - индивидуальная изменчи-
вость деревьев выражена значительно слабее, что сближает мезотрофный дре-
востой с олиготрофным. Примерно такими же чертами обладает сосновый дре-
востой сосняка кустарничково-зеленомошно-сфагнового (см. рис. 5.9, б), от-
ражающего хотя и олиготрофные, но более мягкие условия, чем в сосново-
кустарничково-сфагновой фации. Но в отличие от мезотрофных древостоев
здесь намечается слабая дифференциация на три возрастных периода, ярко
выраженные, как отмечено выше, в крайне олиготрофных условиях.
По сравнению с хвойными лородами ход роста модельных деревьев бе-
резы как в евтрофных, так и в мезотрофных (рис. 5.10) условиях харак-
теризуется более высоким темпом, относительной равномерностью и меньшей
индивидуальной изменчивостью.
5.3.
ПЕРВИЧНАЯ БИОЛОГИЧЕСКАЯ ПРОДУКТИВНОСТЬ
КАК ПОКАЗАТЕЛЬ СООТНОШЕНИЯ
ЛЕСО- И БОЛОТООБРАЗОВАТЕЛЬНОГО ПРОЦЕССОВ
Международная биологическая программа (МБП) стимулировала в свое
время появление множества публикаций, посвященных первичной биологической
продуктивности различных наземных биогеоценозов, Однако в заболоченных
и болотных лесах исследований выполнено относительно мало, что связано в
основном с недостаточной разработанностью их методик и большой трудоем-
костью /Солоневич,
1971/.
Среди отечественных исследований следует наз-
вать работы Н.И. Пьявченко /19606, 1967/, Н.И. Пьявченко и З.А. Сибире-
вой /1962/, Н.Г. Солоневич /1963/, Н.И. Базилевич /1967/, Н.И. Шадри-
ной /1968/, В.И. Валуцкого
/1971/,
Н.И. Казимирова и P.M. Морозовой
/1973/,
Ф.З. Глебова, Л.С. Толейко /1975/, F..Z. G-lebov, L.S. Tolei-
ko /1974/, В.И. Валуцкого и А.А. Храмова /1976/, О.С. Ватковского
/1976/, В.М. Медведевой и др. /1977/, А.А. Храмова и В.И. Валуцкого
/1977/, С.Э. Вомперского и А.И. Иванова /1978, 1982/, Л.С. Козловской
и др. /1978/, В.В. Никонова /1978/,. А.Д. Вакурова и А.Ф. Поляковой
/1982/,
А.И. Иванова /1982/, А. Елина и др. /1984/. Из иностранных ли-
тературных источников существенный интерес в свете наших исследований
представляют работы, относящиеся к лесоболотным биогеоценозам лесной зо-
ны Финляндии /Kosonen, 1981; Solmari, Vasander, 1981; Vasander,
1982/,
Канады /Reader, Stewart, 1972/, Англии /Heal, Perkins,
1978/,
и обобщающая монография П. Мура и Д. Беллами /Moore, Bella-
my, 1974/.
Количество и распределение биомассы в ярусах и фракциях фитоценозов
и заключенных в ней химических элементов, характеризуя первичную продук-
тивность, дают также представление о биологическом круговороте, который
можно рассматривать как интегральный показатель характера биогеоценоти-
ческих процессов.
Целью настоящей работы было установить на основе стационарных иссле-
дований в болотных лесах различной трофности количественные показатели
110