Данилова Н.Н. Психофизиология
Подождите немного. Документ загружается.

К факторам, запускающим генерацию эндогенного ритма,
относятся синаптические и гуморальные влияния, действие
пепти-дов, диффузное внесинаптическое влияние медиаторов.
Они включают и выключают режим гтейсмекерной
активности. Управление пейсмекерной активностью экстра
клеточными факторами обеспечивается взаимодействием
локуса генерации эндогенного ритма с хемовозбудимой и
электровозбудимой мембраной. Показана зависимость
ритмической активности нейронного пейсмекера от уровня
деполяризации нейрона. При слабой деполяризации основной
вклад в генерацию эндогенных потенциалов вносят
низкопороговые Са^-каналы, при высоких значениях
деполяризации — высокопороговые. С заменой
низкопороговьтх Са^-каналов на высокопороговые частота
эндогенного ритма возрастает.
Пейсмекерные нейроны используются для управления
реакциями организма. С их помощью реализуются
генетические программы (например, локомоции). Благодаря
их зависимости от си-наптических влияний они могут
обеспечивать более гибкое применение генетических
программ в адаптивном поведении.
Пейсмекерный механизм обладает пластичностью,
которая выражается в том, что ответ нейрона меняется по
мере повторения действия одиночных раздражителей. В том
случае, если ответ пейсмекерного нейрона слабеет от
применения к применению, этот процесс по аналогии с
угасанием поведенческой реакции называют привыканием.
Кроме привыкания, имеет место обратный процесс
возрастания реакции нейрона от применения к применению.
Его называют фасилитацией. Пластические реакции у пей-
смекерного нейрона могут обеспечиваться изменением как
возбудимости пейсмекерного механизма, так и
эффективности синоптической передачи. Пластические
свойства пейсмекерного механизма существенно расширяют
адаптационные возможности организма в манипуляции
генетическими программами поведения.
6.4. РОЛЬ ИОННЫХ ПРОЦЕССОВ И
ВНУТРИКЛЕТОЧНЫХ ВЕЩЕСТВ В
ПЛАСТИЧНОСТИ НЕЙРОНОВ
Для того чтобы понять, что происходит в нейроне во время
привыкания или сенситизации, а также ассоциативного
обучения, следует рассмотреть ионный механизм нормальной
передачи сигнала через химический синапс от одного нейрона
к другому. Во время генерации ПД ионы натрия и кальция
входят в клетку, а ионы калия выходят из нее. Калиевый ток
из клетки создает волну гиперполязизации, которая обрывает
ПД и ВПСП. ПД распространяется от сомы вдоль аксона с
помощью потенциал-зависимых
140
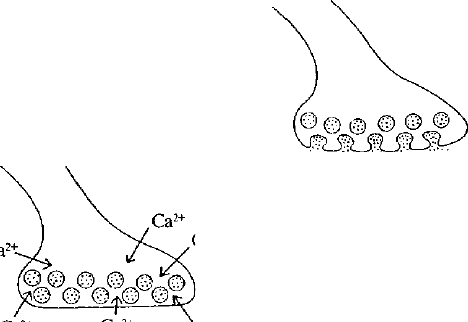
^--О^РН^ ^-^-^-~-
Постсиналтическая
клетка а
Са^
^ © © © ф ©)
Рис. 29. Важнейшие события, происходящие в
химическом синапсе после прибытия импульса в
окончание аксона.
а — потенциал действия открывает каналы для входа Са" в
пресинаптическое окончание; б — синаптические
пузырьки сливаются с пресинаптической мембраной,
высвобождая при этом медиатор; в — медиатор связывается
с белками постсинаптической мембраны, изменяя их
конформацию; г — после удаления медиатора из щели
постсинаптические белки возвращаются в исходное
состояние (по Б. Альбертсу и др., 1994).
натриевых каналов, которые открываются и закрываются по
мере движения нервного импульса. Когда ПД достигает
окончания аксона, под его влиянием открываются потенциал-
зависимые кальциевые каналы, находящиеся в мембране
окончания аксона. Ионы Са^ входят в аксон, в результате чего
в синаптическую щель выбрасываются кванты медиатора. На
рис. 29 представлено конечное звено высвобождения
медиатора в синаптическую щель. Синапти-ческая передача
осуществляется с помощью высвобождения ме-
141
диатора, которое невозможно без притока ионов кальция из
внеклеточной среды. Если удалить ионы кальция из
экстраклеточного пространства, то прибывший к окончанию
аксона ПД не высвобождает медиатор и передачи сигнала не
происходит. Если же искусственно увеличить содержание
ионов кальция в аксоне (ввести через пипетку), это вызовет
высвобождение медиатора даже в отсутствие электрического
импульса (результаты получены на синапсе между
гигантскими нейронами кальмара). Чем выше концентрация
ионов кальция в окончании аксона, тем выше скорость
выделения медиатора.
Особая функция ионов кальция в передаче сигнала
объясняется тем, что он в пресинаптическом нейроне
запускает цепочку биохимических процессов, которая
увеличивает число активированных Са
24
^ каналов. Это
создает эффект усиления выброса медиатора из
пресинаптического окончания. Последовательность био-
химических событий, развертывающихся в пресинаптичском
нейроне после входа в него ионов Са
2
"*", выглядит
следующим образом. В нейроне для ионов Са
2
^ существует
внутренний рецептор— каль-модулин. Это специфический
белок, с которым кальций образует комплекс. Возникнув,
последний активирует фермент аденилат-циклазу. С ее
помощью из АТФ синтезируется цинический адено-
зинмонофосфат (цАМФ), активирующий протеинкиназы.
Проте-инкиназа А фосфорилирует белки потенциал-
независимых кальциевых каналов в результате
присоединения к ним фосфатной группы. Это открывает
дополнительное число Са^-каналов. В клетку входит еще
большее количество ионов кальция, что дополнительно
увеличивает выход медиатора из нейрона. Ионы кальция, как
и цАМФ, выполняют роль посредников в выбросе медиатора
из пресинапти-ческой мембраны. Работа клеточного
аппарата, обеспечивающего преобразование ПД в выход
медиатора и тем самым передачу сигнала через химический
синапс от нейрона к нейрону, представлена на рис. 30 (вход
I).
Этот же аппарат клетки, обеспечивающий передачу
сигнала от нейрона к нейрону, используется и для
длительных изменений синаптической проводимости,
связанных с обучением: привыканием, сенситизацией, а
также с ассоциативным обучением, главным образом на
начальном его этапе. Критическая роль в этих процессах
принадлежит модулирующим нейронам. Они имеют свой вход
к этому клеточному аппарату — вход II (см. рис. 30). У
моллюсков это серотонинергические нейроны.
Внутриклеточной мишенью для серотонина, так же как для
комплекса кальций+кальмодулин, является аденилатциклаза,
за счет которой из аденозинтрифосфа-та (АТФ) —
универсального источника энергии для биохимичес-
142
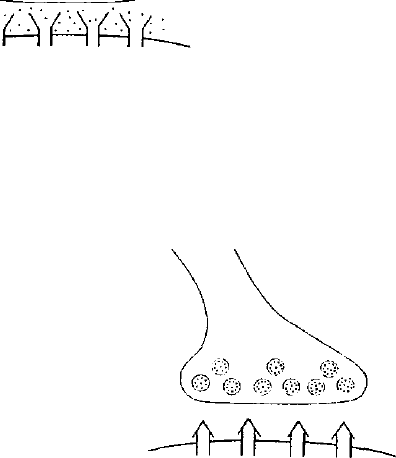
Вход II ,
(эффект серотонин- )
модулирующей
системы) '
Вход 1
(эффект ПД)
Открытие потенциал-
зависимых Са^-
каналов
Са^ + кальмодулин
Аденилатииклаза ^
АТФ
цАМФ
(циклический нуклеотид)
_А-
Рис. 30. Основные этапы фосфорилирования Са^- и К-
каналов, обеспечивающего кратковременное изменение
проводимости синапса.
ких процессов синтезируется циклический
аденозинмонофосфат (цАМФ). За счет двойного действия
на аденилатциклазу Са
2
^ и серотонина возрастает
образование цАМФ. Активированные киназы фосфорилируют
белки ионных каналов. Киназа А взаимодействует с белками
кальциевых каналов и открывает их для входа ионов кальция.
Киназа С фосфорилирует белки калиевых каналов, вызывая
143
Протеинкиназа А
Протеинкиназа С
Фосфорилировани
е белков Са^-
каналов
Фосфорилировани
е белков К-
каналов
Открытие
потенциал-
независимых
Са
2
"
1
" -каналов
Закрытие К-каналов
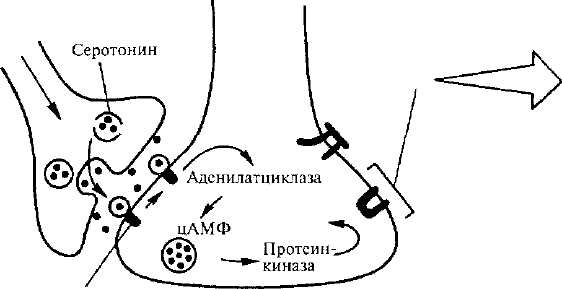
Серотонин
активирует
рецепторы
Повторное
раздражен
ие
головы
Протеинкиназа
фосфорилирует
калиевые
каналы и
ипактивируст
их
Регистрация
калиевого
тока через
изолиро-
ванный
кусочек
мембраны
Рис. 31. Зависимое от активности пресинаптическое
облегчение у аплизии (по Э. Кэнделу и Р. Хокинсу, 1992).
их закрытие. Это уменьшает калиевый ток из нейрона и
калиевую гиперполяризацию, следующую за ПД и ВПСП, что
приводит к удлинению ПД. На рис. 31 показана цепочка
реакций, приводящих к инактивации К-каналов, вызываемой
возбуждением модулирующих нейронов. Продление ПД
удерживает потенциал-зависимые Са^-каналы в открытом
состоянии, что увеличивает в нейроне содержание свободного
кальция и выброс медиатора. Фосфорили-рование калиевых
каналов (8-каналов ) рассматривается как одна из форм
кратковременной памяти, которая ограничена временем
жизни белка, образующего 5-каналы, и легко стирается при
воздействии фосфопротеинфосфотаз (дефосфорилирующих 5-
каналы).
События, разворачивающиеся на постсинаптическом
нейроне у млекопитающих в ситуации обучения, связаны с
особой группой рецепторов, медиатором для которой служит
глутамат. Среди них различают NМ^А-рецепторы (они
селективно активируются синтетическим аналогом глутамата
Т^-метил-0-аспартатом) и не-NМ^А-рецепторы. Каналы,
связанные с NМ^А-рецепторами, имеют двойные ворота,
открывающиеся при выполнении двух условий: мембрана
должна быть сильно деполяризована, а к рецептору должен
присоединиться медиатор глутамат (рис. 32). Каналы,
связанные с ММОА-рецепторами, находясь в открытом
состоянии, пропускают очень большой поток ионов Са
2
"*".
Эта их отличительная особенность очень важна для
механизма памяти. Не-КМОА-рецепторы, реагируя на
глутамат, сразу открывают кальциевые
144
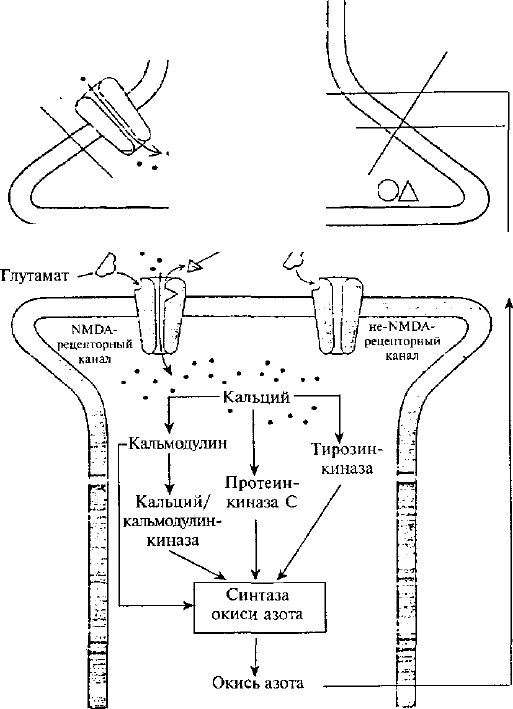
Рис. 32. Внутриклеточные процессы в постсинаптическом
нейроне гиппо-кампа, реагирующего на глутамат (по Э.
Кэнделу, Р. Хокинсу, 1992).
ионные каналы без дополнительных условий и Са^ входит в
нейрон. Это создает уровень деполяризации нейрона, который
необходим для реакции г^МОА-рецепторов. Деполяризация
нейрона снимает блокаду с гШОА-каналов, которая
осуществляется внеклеточным магнием (Мё). Изменения на
посте инаптической мембране с участием NМ^А- и не^МОА-
рецепторов наиболее хорошо изучены у нейронов гиппокампа
на модели долговременной
145
10-3341
Пресинапти-
чсскии
нейрон
Кальций
Пузырек 'уГ\ с
нейромедиа-
тором
Усиленное
выделение
нейромедиатор
а
Участок
выделения
нейромедиатор
а
трансфераза
сОМР <—
Гуанилат-< *
циклаза
АОАОАОАОАОАОА
Кальций • . Магний /. ^
^
Постеннапти
-ческий
нейрон
Ретроград
ный
мессендже
р (окись
азота)