Castilho Leda R., Moraes Angela Maria (Ed.) Animal Cell Technology: From Biopharmaceuticals to Gene Therapy
Подождите немного. Документ загружается.

decrease in cell size, by a controlled and uniform DNA fragmentation,
and by specific changes in the chromatin, which becomes highly con-
densed and can appear concentric with the nuclear membrane. The only
alteration that may be observed in the organelles is a swelling of the
endoplasmic reticulum (ER) (McCarthy, 2002). However, during cultiva-
tion processes in vitro, apoptosis ends up in a stage called secondary
necrosis (Figure 7.3; see color section), where, due to the absence of
specialized phagocytic cells, the apoptotic cells and apoptotic bodies
release their contents into the extracellular medium, as occurs in necrosis
(Al-Rubeai, 1998).
7.4 Influence of environmental conditions on the induction of
cell death
During in vitro cultivation of animal cells, any change relative to the
optimal environmental conditions may result in a rapid decrease in cell
viability. This phenomenon is more frequently observed in cell lines that
exhibit high susceptibility to apoptosis (Singh and Al-Rubeai, 1998). Any
agent or condition that alters the cell metabolism may activate pro-
grammed cell death. Cells exposed to high stress levels due, for instance,
to the presence of a high concentration of toxins, to large changes in pH,
or to high agitation rates, generally die by necrosis, because there is no
time for the cell to generate a response, that is, cell death is instantaneous.
At intermediate stress levels, the cell can be damaged without instanta-
neously dying, having time to activate its programmed death mechanism.
Under low levels of environmental stress, the cells may produce heat-
shock proteins (HSPs), which allow their survival until the stimulus is
eliminated. However, once a threshold level of resistance is surpassed,
survival becomes impossible and the cells activate their apoptotic path-
ways (Cotter and Al-Rubeai, 1995).
Cells in culture are usually exposed to variations in environmental
conditions and, depending upon these conditions, cells can survive or die.
In biotechnological processes, the usual aim is to maintain cells alive and
productive. There are a large number of factors that influence the quality
of a cell culture, including an appropriate supply of nutrients, pH, shear
stress, accumulation of metabolites, and oxygen availability. Studies have
already shown that several alterations in culture conditions, such as the
lack of nutrients or serum, increase in osmotic pressure, and maintaining
cells in suspension culture, may induce apoptotic mechanisms. The critical
factors determining apoptosis vary between cell lines (Singh et al., 1994;
Zanghi et al., 1999; Kim and Lee, 2002).
7.4.1 Depletion of nutrients and growth factors
Cells survive and proliferate according to the addition of appropriate
amounts of nutrients and growth factors, which are provided by the
culture media. When these requirements are not met, cells die. The
availability of nutrients and growth factors is considered the most signifi-
cant factor influencing the biological activity of cells. The lack or depletion
152 Animal Cell Technology

of these substances can not only lead to arrest of proliferation, but also to
cell death by apoptosis.
In the cells, control of proliferation and apoptosis is carried out by an
integrated signaling mechanism that activates or avoids one of these
processes and also influences other aspects of cell activity. For instance,
the presence of mitotic agents that induce proliferation, and survival
factors that inhibit apoptosis, are necessary for maintaining cell viability
(Al-Rubeai, 1998). It is important, however, to distinguish between mito-
gens and survival factors. Insulin-like growth factor 1 (IGF-1) is a potent
survival factor, but a poor mitogen. Many cytokines are good mitogens,
but poor survival factors, since they induce cell division but do not avoid
apoptosis. In this way, induction of cell growth and apoptosis inhibition
become independent events. Macrophage-colony stimulating factor
(M-CSF), for instance, promotes cell survival when present at low concen-
trations, but promotes proliferation at high concentrations. Thus, the
correct balance of growth and survival factors is an important considera-
tion for maintaining high cell viability in culture (Cotter and Al-Rubeai,
1995).
Animal serum, especially serum from bovine fetuses, plays an important
role in the growth and survival of animal cells in culture. Serum contains
growth factors, proteins, and other nutrients. The removal of serum has
been associated with apoptotic induction in different cells, including
hybridomas and Chinese hamster ovary (CHO) (Zanghi et al., 1999) and
plasmacytomas (Singh et al., 1994). The mechanism by which the removal
of growth factors, such as those contained in serum, induces apoptosis
seems to involve the expression of the proto-oncogene c-myc (Al-Rubeai,
1998). In CHO cells, apoptosis is induced by nutrient depletion due to
accelerated nutrient consumption in the absence of serum. On the other
hand, when serum or plasma is present at high concentrations, death by
apoptosis is also induced, but this phenomenon can be avoided by adding
supplements containing thiol groups, such as L-cysteine or L-tryptophan
(Al-Rubeai, 1998).
When cells are deprived of an energy source such as glucose or glutamine,
apoptosis can be induced. The effects of lack of glutamine on apoptosis
induction have been observed in hematopoietic cells, such as hybridomas
and plasmacytomas (Mercille and Massie, 1994a; Singh et al., 1994). Sanfeliu
and Stephanopoulos (1999) demonstrated that the lack of glutamine is the
major factor activating apoptosis in a CHO cell line producing interferon-
gamma, and that this effect could be attenuated by the expression of the
gene Bcl-2, which codes for an anti-apoptotic protein that participates in
the signaling pathway that is triggered in response to apoptotic stimuli. The
influence of glucose and glutamine on the apoptotic pathways is not yet
completely understood, but the concentrations of both compounds regulate
cell growth and the catabolic pathways. Their depletion can block DNA
synthesis and arrest the culture in the G
1
phase, causing apoptosis. How-
ever, this does not explain why cells in phases S, G
2
, and M also undergo
apoptosis. It is suggested that apoptosis can be induced when the energy
required for synthesis of the ATP, needed for the duplication of cellular
material, is decreased to a critical level, resulting in the activation of
regulatory proteins that promote apoptosis (Al-Rubeai, 1998).
Mechanisms of cell proliferation and cell death in animal cell culture
in vitro
153

The relationship between apoptosis induction and glutamine depletion
in batch cultures of hybridomas led to studies suggesting that apoptosis is
activated upon depletion of any amino acid. Although the lack of essential
amino acids is critical, the depletion of any amino acid can induce
apoptosis, since the synthesis of molecules involved in the regulation of
cell death is blocked (Simpson et al., 1998). There are two probable
explanations for this. According to one of them, the decrease in the
intracellular concentration of amino acids can lead to a depletion of tRNA
molecules, blocking translation (protein synthesis). According to the
second theory, transcription and translation can be impaired by the de-
crease in the intracellular ATP levels. Although not all amino acids
contribute to the energetic requirements of the cells, the absence of any
amino acid results in the induction of apoptosis. Experiments with murine
hepatocytes showed that protein synthesis is inhibited at the level of the
initiation of the peptide chain. This indicates that the role of amino acids
as protein precursors could be one of the most significant factors inducing
death by apoptosis (Singh and Al-Rubeai, 1998).
7.4.2 Oxygen limitation
The apparent involvement of free radicals in apoptotic processes has led to
studies of the effects of dissolved oxygen tension on animal cells in culture.
Hydrogen peroxide was demonstrated to be a factor inducing apoptosis,
whereas antioxidants such as catalase and superoxide dismutase protect
cells. Formerly, it was believed that the Bcl-2 protein acted as an antiox-
idant, but today it is known that anoxic conditions can induce apoptosis
(Mercille and Massie, 1994a) and that Bcl-2 expression presents an anti-
apoptotic action also under limiting conditions of oxygen. These studies
are very important for large-scale cultivation processes and for processes
where oxygen may be limiting, since the supply of oxygen is a key factor
that limits the viable cell concentration attained in these culture systems.
The effects of Bcl-2 protection under anoxic conditions have already been
demonstrated for different cell lines. For cells derived from Burkitt’s
lymphoma transfected with the Bcl-2 gene, the resistance to anoxic
conditions increased, and cell viability could be maintained at high levels
for a longer period than with control cells (Singh et al., 1997). In the case
of a hybridoma, the expression of this gene also conferred a higher
viability and integrity of the cells when exposed to anaerobic conditions
(Simpson et al., 1997).
7.4.3 Susceptibility to shear stress
Animal cells do not have a cell wall and are, thus, highly susceptible to the
effects of shear stress. Cells respond to hydrodynamic stress within
minutes, altering their metabolism and the gene expression pattern (Nol-
lert et al., 1991). Under sub-lethal levels of shear stress, there is initially an
increase in passive transmembrane transport, simultaneously with damage
to surface receptors. The plasma membrane is generally the main site for
shear damage, and it may lose its capacity to mediate the transport of ions
and molecules, so that the cell loses its viability. It has been demonstrated
154 Animal Cell Technology

that the susceptibility of some cell lines to the effects of hydrodynamic
stress is related to cell size and cell cycle, with cells in S and G
2
phases
more prone to damage and smaller cells more resistant to injury. It has
also been observed that cell damage and the dominating death mechanism
(necrosis or apoptosis) depend on the intensity and exposure time to shear
stress (Al-Rubeai et al., 1995a, 1995b). Exposing hybridoma cells to
moderate agitation levels resulted in a decrease of viability and an increase
in cell death by apoptosis (Al-Rubeai et al., 1995b), whereas extreme shear
levels led to cell death by necrosis, with complete cell fragmentation.
The hydrodynamic environment in a stirred-tank bioreactor can be
stressful for animal cells. This arises from the high energy introduced into
the system to maintain homogeneity and cells in suspension. Furthermore,
gas bubbling results in cell damage, especially at the gas–liquid interface
near the headspace of the bioreactor. While knowledge of the effects of
hydrodynamic conditions has improved bioreactor design and operation
strategies, so as to decrease hydrodynamic stress, a compromise still exists
concerning the need for efficient agitation and aeration and the suscept-
ibility of cells to shear. Certainly, if apoptosis is induced by hydrodynamic
stress, then the suppression of this death mechanism, e.g. by molecular
strategies, would allow enhanced agitation and aeration, improving mass
transfer with a decreased effect on cell viability (Singh and Al-Rubeai,
1998).
7.4.4 Osmolality
Culture medium osmolality has a large influence on animal cell culture.
Hyperosmotic conditions, achieved by using salts, CO
2
, or concentrated
media, has been shown as a low-cost method to increase specific produc-
tivity of cells. Hybridoma cultures, for instance, show improved produc-
tivity with an increase in osmolality (Oh et al., 1993). However, this
method is not very popular due to the negative effects on cell growth. The
drop in cell growth under hyperosmotic conditions occurs, probably, due
to cell death by apoptosis. Thus, expression of anti-apoptotic genes, such
as Bcl-2, could allow the use of hyperosmotic conditions to simulta-
neously limit cell death and increase cell productivity (Kim and Lee,
2002).
7.5 Methods of detection of cell death by apoptosis
Currently, there are many methods available for determining cell death by
apoptosis in cell cultures and tissues. These methods are based essentially
on changes that occur in apoptotic cells. During apoptosis, several
phenomena can be observed, such as DNA fragmentation, chromatin
condensation, nuclear fragmentation, cytoplasm acidification, cytochrome
c release from the mitochondria, exposure of intracellular phospholipids
and the activation and breakdown of proteins. These apoptotic phenomena
can be detected by direct or indirect methods, on cell populations or on
individual cells that are representative of a population. The main principles
used by the different detection methods are:
Mechanisms of cell proliferation and cell death in animal cell culture
in vitro
155

(i) DNA fragmentation;
(ii) morphological changes;
(iii) changes in membrane asymmetry;
(iv) activation of apoptotic proteins;
(v) release of cytochrome c in the cytoplasm.
7.5.1 DNA fragmentation
During controlled cell death, the DNA is initially cleaved into 300–500 kb
fragments (Brown et al., 1993). Later on, these fragments are decreased to
multiples of 180–200 bp, which is the distance between histone proteins,
around which the condensed chromatin is wrapped. This phenomenon
occurs simultaneously with morphological changes, such as chromatin
condensation. The resulting fragments are known as LMW-DNA (low
molar mass DNA) fragments, which are able to pass through pores of the
nuclear membrane and reach the cytoplasm, and HMW-DNA (high molar
mass DNA) fragments, which remain in the nucleus. The main enzyme
involved in this process is a caspase-activated DNAse (CAD) (Nagata,
2000).
Agarose gel electrophoresis
The DNA fragments can be visualized in electrophoretic runs in agarose
gels showing a distinct band pattern (DNA laddering). The sizes of the
bands are multiples of the smallest nucleosomal fragments. Unlike those of
apoptosis, the DNA fragments arising from necrosis are of irregular size,
resulting in a smear in the sample lane. Although the gel electrophoresis
pattern is a biochemical indication of apoptosis, in some cases the classic
morphological features of apoptosis may be noted without DNA fragmen-
tation (Al-Rubeai, 1998; Nagata, 2000).
TUNEL
TUNEL stands for terminal deoxynucleotidyl transferase x-dUTP nick
end labeling. This assay is based on the detection of DNA fragments
marked by an enzyme that incorporates modified nucleotides to the 39-
OH ends of the fragments, which can be then specifically detected. The
enzyme is a deoxynucleotidyl transferase, which can act in absence of a
complementary strand. Among the nucleotides, there is one specifically
marked with a fluorochrome, an enzyme, or an antigen. This allows
different methods of detection.
When the deoxynucleotide is associated with a fluorochrome, the cells
can be observed under a fluorescence microscope, whereby apoptotic cells
present an intense fluorescence and, in advanced stages, nuclear fragmenta-
tion can be visualized. For a quantitative analysis, either a hemocytometer
in an optical microscope or a flow cytometer can be used (Tinto et al.,
2002). Peroxidase-marked deoxynucleotides can be quantified by chromo-
genic tests that use the enzyme substrate. Indirect methods use antigens
linked to the nucleotide and recognized by labeled antibodies. However,
156 Animal Cell Technology

this is a less sensitive technique, which is used mainly in pre-fixed
histological samples.
In the TUNEL test, it is necessary to permeabilize the cells to introduce
the enzyme and the deoxynucleotides, but the permeabilization is carried
out after weak fixation with formaldehyde, so that low molar mass
fragments are not lost. Permeabilization is carried out in an ice bath,
followed by labeling with the reaction solution.
Propidium iodide, ethidium bromide, and DAPI
These methods use fluorescent labels, such as propidium iodide, ethidium
bromide, or DAPI (49,69-diamidino-2-phenylindole), which are incorpo-
rated into the DNA, allowing chromatin condensation and nuclear frag-
mentation to be visualized under a microscope with the appropriate
fluorescence filters. To allow fluorochromes to enter the cells and reach
the nucleus, the cells need to be prepermeabilized, for example, with 70%
ethanol at –208C. LMW-DNA fragments may be lost by the permeabili-
zation, decreasing the amount of DNA inside the cells. The lower nucleic
acid concentration results in a lower fluorescence intensity in apoptotic
cells, which can be detected by fluorescence microscopy or flow cytome-
try (Calle et al., 2001).
7.5.2 Morphological changes
Several methods are based on microscopy observations to detect morpho-
logical changes in the cells, mainly chromatin condensation. Different
markers can be used to differentiate between the stages of apoptosis.
To distinguish apoptotic from normal and necrotic cells, fluorescent
dyes can be used to form complexes with DNA, such as acridine orange,
ethidium bromide, or propidium iodide (Mercille and Massie, 1994b).
Fluorescence microscopy following cell labeling clearly reveals chromatin
condensation and fragmentation in apoptotic cells. By combining the use
of fluorescent dyes that are able to label the chromatin in intact cells (e.g.
acridine orange) with those that just label the chromatin in cells with a
damaged membrane (e.g. ethidium bromide and propidium iodide), it is
possible to identify different levels of cell viability (Mercille and Massie,
1994a).
Acridine orange is able to penetrate cells independently of the integrity
of the cell membrane. It intercalates the bases in double strands of DNA
and emits green fluorescent light. It can also form complexes with RNA
and single-stranded DNA, but in this case it does not intercalate the bases
and emits red fluorescent light. Thus, a viable cell can be visualized with a
green nucleus and, possibly, reddish spots in the cytoplasm.
Ethidium bromide is not able to permeate the plasma membrane, and
only penetrates non-viable cells that have lost the selective permeability of
the membrane. It can intercalate the bases of double-stranded DNA
molecules, emitting an orange fluorescent light. Ethidium bromide also
forms weak complexes with RNA, resulting in red fluorescence. Thus,
non-viable cells will present a strong orange fluorescence, since ethidium
Mechanisms of cell proliferation and cell death in animal cell culture
in vitro
157

bromide labeling overlays acridine orange fluorescence. If there is still
cytoplasmic material inside the cell, it will be labeled dark red.
As shown in Figure 7.3 (see color section), the morphologies observed
in a sample incubated with acridine orange and ethidium bromide allow
the identification of four levels of cell viability, as described below:
(i) Viable nonapoptotic (VNA) cells show the nucleus uniformly colored
green, with no chromatin condensation. In this cell population, only
acridine orange is able to penetrate the cells and bind to the DNA.
(ii) Early apoptotic cells, also called viable apoptotic (VA), show brilliant
green granules in the nuclear region. These granules correspond to the
condensed and fragmented chromatin, which means that the cells
have already initiated the apoptotic process, but still keep the selective
permeability of the membrane. In this phase, the formation of
apoptotic bodies containing material that will be expelled by the cells
can be observed.
(iii) Late apoptotic cells, also known as non-viable apoptotic (NVA), are
colored reddish-orange, with granules in the nuclear region showing
brilliant orange fluorescence. The cells have lost membrane integrity
and have become permeable to ethidium bromide. Another character-
istic of this phase is the decrease in cell size, due to the loss of cell
material as a result of elimination of apoptotic bodies.
(iv) Necrotic cells are totally colored orange, indicating loss of selective
permeability of the membrane. These cells do not show any conden-
sation or fragmentation of the chromatin, and present no decrease in
size. On the contrary, there is a tendency for an increase in cell
volume. No apoptotic bodies are formed. This stage is very short, and
cell lysis occurs within a short time.
7.5.3 Membrane asymmetry
Translocation of the phosphatidylserine from the inner to the outer leaflet
of the plasma membrane is an initial event related to the apoptotic process
and possibly serves as a signal for the removal of apoptotic bodies by
phagocytic cells (Martin et al., 1995). The exposure of this phospholipid
has been largely used as a specific apoptosis marker.
Membrane asymmetry changes can be detected by flow cytometry using
a fluorescent marker (e.g. fluorescein, FITC) conjugated to annexin V, a
protein that has high affinity for phosphatidylserine. When using a
fluorescence microscope, this technique can be quantitative if a hemocyt-
ometer is used.
Unfortunately, annexin V also binds to phosphatidylserine residues in
the inner leaflet of the plasma membrane of necrotic cells due to the loss of
integrity of the membrane. However, the use of propidium iodide as a
counter-marker allows viable, apoptotic, and necrotic cells to be distin-
guished (Pla
¨
sier et al., 1999).
158 Animal Cell Technology

7.5.4 Apoptotic proteins
There are a large number of genes related to apoptosis, and the corre-
sponding proteins modulate the cell death process. Tests that detect these
proteins or their activities are largely used to detect apoptosis in cell
culture. This group of apoptotic proteins is very large and, theoretically,
any key protein participating in apoptotic pathways, such as p53 protein
and caspases 3 and 7, can be used to detect apoptosis (Kim and Lee, 2002;
Arden and Betenbaugh, 2004).
7.5.5 Cytochrome c release
Alterations of the mitochondrial membrane may precede those that occur
in the nucleus. These changes can involve both the inner and the outer
membrane, leading to a dissipation of the transmembrane potential and/or
to the release of intermembrane proteins through the outer membrane.
The main group of proteins responsible for mitochondrial alterations
consist of the proteins known as Bax, which form pores in the outer
membrane, causing the release of cytochrome c to the cytoplasm (Loeffler
and Kroemer, 2000; Arden and Betenbaugh, 2004). Western blot techni-
ques can be used to specifically detect the presence of cytochrome c in the
cytoplasm of apoptotic cells. However, complex purification protocols are
required, and there is the possibility of incomplete separation of
mitochondria from the cytoplasm; therefore, this technique is not very
popular.
7.6 Apoptosis suppression by molecular techniques
7.6.1 Molecular basis of apoptotic cell death
Much of the current knowledge of the genetic regulation of apoptosis
began with genetic studies of the nematode Caenorhabditis elegans,in
which 131 of 1090 somatic cells formed during adult development die by
apoptosis. This cell death is regulated by a group of approximately 13
genes (Horvitz et al., 1994). The apoptotic mechanism identified in C.
elegans seems to be highly evolutionarily conserved in vertebrates and
invertebrates, since homolog genes were also discovered in mammals,
although in a much greater amount.
The events involved in apoptosis can be divided into four phases:
initiation, signaling, effector, and degradation (Mastrangelo and Beten-
baugh, 1998). During the initiation phase, the cell is exposed to insults that
trigger the apoptotic cascade. The main insults found in cell cultures are:
nutrient depletion (such as glucose, glutamine, and other amino acids),
deprivation of growth and survival factors (e.g. factors in fetal bovine
serum), accumulation of toxic metabolites (ammonia and lactate), oxygen
concentration (hypoxia and hyperoxia), hydrodynamic stress, loss of cell
adhesion (e.g. during suspension adaptation), viral infection (e.g. when
viral vectors are used for recombinant DNA transfer), and protein over-
expression. Once apoptosis has been triggered (initiation), the message is
transmitted through the signaling cascade during the signaling phase. This
Mechanisms of cell proliferation and cell death in animal cell culture
in vitro
15 9

stage of apoptosis includes several parallel pathways that converge to a
few or even a single pathway of the effector phase. Each insult probably
activates a different signaling cascade pathway, finally leading to the
effector phase, which involves the activation of caspases, the proteases
responsible for cellular degradation (Dickson, 1998). During this last
phase, which is irreversible, it is possible to observe the specific morpholo-
gical changes of apoptosis, such as chromatin condensation, nuclear
fragmentation, cellular shrinkage, blebbing of plasma membrane, and
formation of apoptotic bodies. Different cell lines respond to different
insults with varying intensities, possibly as a result of activation of differ-
ent apoptotic pathways (Singh and Al-Rubeai, 1998).
The caspase family
The characteristic morphological features of apoptosis are due to the direct
or indirect action of caspases, a highly conserved protease group (Alnemri
et al., 1996). Caspases are cysteine-aspartate proteases, members of a
cysteine-protease family that specifically cleave their substrates after
aspartic acid residues. To date, at least 14 mammalian caspases have been
identified. Although they share a common structure, they can be divided
into two functional groups: those that participate in the apoptosis process
(caspases 2, 3, 6, 7, 8, 9, 10, and 12) and those that are responsible for
cytokine processing during immune response and are involved in the
inflammatory process (caspases 1, 4, 5, and 11) (Alnemri et al., 1996).
However, due to the lack of functional information on some of the
caspases (e.g. caspases 2, 4, 5, 7, 11, 12, and 13), the exact division between
the two groups is still ambiguous.
Caspases are expressed as zymogens or procaspases and require a
proteolytic cleavage at specific aspartic acid residues to become active.
These cleavages generate four fragments: the N-terminal prodomain (2–25
kDa), the large subunit (17–21 kDa), the C-terminal small subunit (10–13
kDa), and a small linker between large and small subunits (Figure 7.4).
Some caspases do not possess the small linker. An active caspase consists
of a heterotetramer containing two small and two large subunits, which
are derived from two procaspase molecules (Walker et al., 1994). The N-
terminal prodomain and the linker (if present) are removed from the
mature and active caspase. The observation that caspases, to become active,
undergo the same process as their substrates (they are cleaved at aspartic
acid residues) suggests a caspase proteolytic cascade, that is, an active
caspase can subsequently cleave and activate other procaspases (Thorn-
berry et al., 1997).
Caspases involved in the apoptotic process can be subclassified as
initiator or effector caspases, depending on the structure of their prodo-
main. This also reflects their different roles in the apoptotic cascade
(Earnshaw et al., 1999).
Initiator caspases (caspases 2, 8, 9, 10, and 12) possess a functional
prodomain. This prodomain is large and contains interaction domains
(structural motifs) where adaptor proteins, like Apaf-1 and FADD, can
bind. The interaction with adaptor proteins is important for the caspase
activation process. There are two types of interaction domains: DED
160 Animal Cell Technology
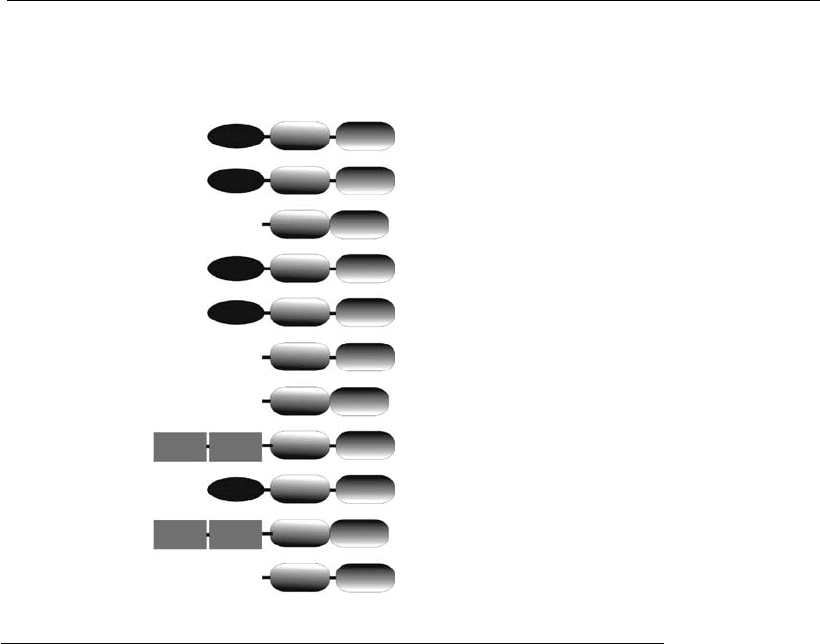
(death effector domain) and CARD (caspase recruitment domain) (Muzio
et al., 1996; Hofmann et al., 1997). The contact with CARD domains is
based on electrostatic interactions, while the contact with DEDs is based
on hydrophobic interactions. Caspases 2, 9, and 12 have CARDs and
caspases 8 and 10 have DEDs. Generally, initiator caspases act upstream of
the effector caspases and are responsible for their activation (Thornberry
et al., 1997; Earnshaw et al., 1999).
The main activation mechanism of initiator caspases is autoproteolytic,
which is dependent on procaspase aggregation. Caspases possess low
intrinsic enzymatic activity when in a zymogen form. Their catalytic
activity increases when they are cleaved and assembled into tetramers
(Yamin et al., 1996). It has been observed that procaspase aggregation or
oligomerization facilitates intermolecular proteolysis, which results in
autocatalysis (Srinivasula et al., 1998; Yang et al., 1998). Adaptor proteins
recruit and bring together initiator caspases into activating complexes,
which allows the autoactivation.
Effector caspases (caspases 3, 6, and 7) may possess a small inactive
prodomain or may lack it completely, since they do not need to form
Caspase 1
Caspase 2
Caspase 3
Caspase 4
Caspase 5
Caspase 6
Caspase 7
Caspase 8
Caspase 9
Caspase 10
Caspase 14
CARD
CARD
CARD
CARD
CARD
DED DED
DED DED
316 331152 435
297 317119 404
17528 277
270 290 377
311 331 418
179 19423 293
19823 303
374 385
216
479
315 331 416
415
219
521
242
Procaspase domain structure
Preferred
substrate
sequence
WEHD
YEVD
DEHD
VDVAD
DEVD
DNQD
LEVD
(W/L)EHD
(W/L)EHD
VEID
VEHD
DEVD
(I/L)ETD
LEHD
IEAD
?
Function
Inflammation
Apoptosis
Apoptosis
Inflammation
Inflammation
Apoptosis
Apoptosis
Apoptosis
Apoptosis
Apoptosis
Keratinocytes
differentitation?
Figure 7.4
Caspase family. This scheme illustrates the domain structures, internal cleavage
sites, preferred peptide substrate sequences, and bio logical function of caspases.
Each procaspase consists of a large and small domain and may also possess DED
(death effector domain) and CARD (caspase r ecruitment domain) (adapted from
Hill et al., 2003 ).
Mechanisms of cell proliferation and cell death in animal cell culture
in vitro
161