Caballero B. (ed.) Encyclopaedia of Food Science, Food Technology and Nutrition. Ten-Volume Set
Подождите немного. Документ загружается.


the spectra to be obtained on-line. However, the
widespread use of these two parameters as the only
criteria has led to misidentifications. Thus, it has been
recommended that the following minimum criteria be
fulfilled for identification: (1) the visible (or ultravio-
let for shorter chromophores) spectrum in at least two
different solvents must be in agreement with the chro-
mophore suggested; (2) chromatographic properties
must be identical in two systems, preferably TLC (R
F
)
and HPLC (t
R
), and cochromatography with the
authentic carotenoid should be demonstrated; and
(3) a mass spectrum should be obtained, which allows
at least confirmation of the molecular mass. The re-
quirement of a mass spectrum, however, would limit
carotenoid analysis to a very few laboratories around
the world, precluding its execution in areas where it is
very much needed. Moreover, major well-known
carotenoids can be conclusively identified by the
judicious and combined use of chromatographic
data, absorption spectra and specific chemical reac-
tions to confirm the type, location, and number of
functional groups. (See Chromatography: Thin-layer
Chromatography; High-performance Liquid Chro-
matography.)
0043 Mass spectrometry (MS) and nuclear magnetic res-
onance spectroscopy (NMR) are indispensable in the
elucidation of unknown or inconclusive structures of
carotenoids and are increasingly used with food
carotenoids. MS gives the molecular mass, and
typical fragmentations provide information about
the structure. NMR recognizes new end groups and
determines the location of cis double bonds. The
chemical shift data for many end groups and trans/
cis isomers are available from tables for both
1
H-NMR and
13
C-NMR. (See Spectroscopy: Nuclear
Magnetic Resonance.)
0044 The quantification step in OCC is straightforward.
The separated carotenoid fractions are collected and
quantified spectrophotometrically through the use of
tabulated absorption coefficients. HPLC quantifica-
tion is carried out by means of internal or external
calibration, for which the concentrations of the
standard solutions are also determined spectrophoto-
metrically. Unfortunately, few carotenoid standards
are available commercially and are of a widely vary-
ing purity. Other carotenoids have to be isolated and
purified from natural sources by OCC or by accumu-
lating separated fractions from several HPLC runs.
Both procedures are time-consuming and require ex-
perience and patience. The instability of the caroten-
oid standards is another serious problem.
0045 Notwithstanding the inherent difficulties and the
many possible sources of errors, reliable data on food
carotenoids can be obtained in the hands of careful
and well-informed analysts.
See also: Analysis of Food; Antioxidants: Natural
Antioxidants; Cancer: Diet in Cancer Prevention;
Chromatography: High-performance Liquid
Chromatography; Colorants (Colourants): Properties
and Determination of Natural Pigments; Guavas;
Mangoes; Mass Spectrometry: Applications; Oxidation
of Food Components; Papayas; Peaches and
Nectarines; Ripening of Fruit; Tomatoes; Vegetables
of Temperate Climates: Carrot, Parsnip, and Beetroot
Further Reading
Britton G, Liaaen-Jensen S and Pfander H (eds) (1995)
Carotenoids, vol 1A. Isolation and analysis. Basel: Bir-
khauser.
Britton G, Liaaen-Jensen S and Pfander H (eds) (1995)
Carotenoids, vol. 1B. Spectroscopy. Basel: Birkhauser.
Goodwin TW and Britton G (1988) Distribution and
analysis of carotenoids. In: Goodwin TW (ed.) Plant
Pigments, pp. 61–132. London: Academic Press.
Gross J (1987) Pigments in Fruits. London: Academic Press.
Gross J (1991) Pigments in Vegetables. Chlorophylls and
Carotenoids. New York: Van Nostrand Reinhold.
Krinsky NI, Mathews-Roth MM and Taylor RF (eds)
(1990) Carotenoids: Chemistry and Biology. New
York: Plenum Press.
Packer L (ed.) (1992) Carotenoids part A. Chemistry,
separation, quantitation, and antioxidation. Methods
in Enzymology 213: 3–538.
Pfander H (ed.) (1987) Key to Carotenoids, 2nd edn. Basel:
Birkhauser.
Rodriguez-Amaya DB (1993) Nature and distribution of
carotenoids in foods. In: Charalambous G (ed.) Shelf
Life Studies of Foods and Beverages. Chemical,
Biological, Physical and Nutritional Aspects, pp. 547–
587. Amsterdam: Elsevier Science.
Rodriguez-Amaya DB (1999a) A Guide to Carotenoid
Analysis in Foods. Washington, DC: International Life
Sciences Institute Press.
Rodriguez-Amaya DB (1999b) Changes in carotenoids
during processing and storage of foods. Archivos Latino-
americanos de Nutricio
´
n 49: 36S–47S.
Physiology
H C Furr, Craft Technologies, Wilson, NC, USA
R M Clark, University of Connecticut, Storrs, CT, USA
Copyright 2003, Elsevier Science Ltd. All Rights Reserved.
Background
0001The carotenoids are polyisoprenoid compounds (usu-
ally tetraterpenes); most common carotenoids have
40 carbon atoms, although shorter (apocarotenoids)
and longer forms exist. Frequently, carotenoids have
936 CAROTENOIDS/Physiology
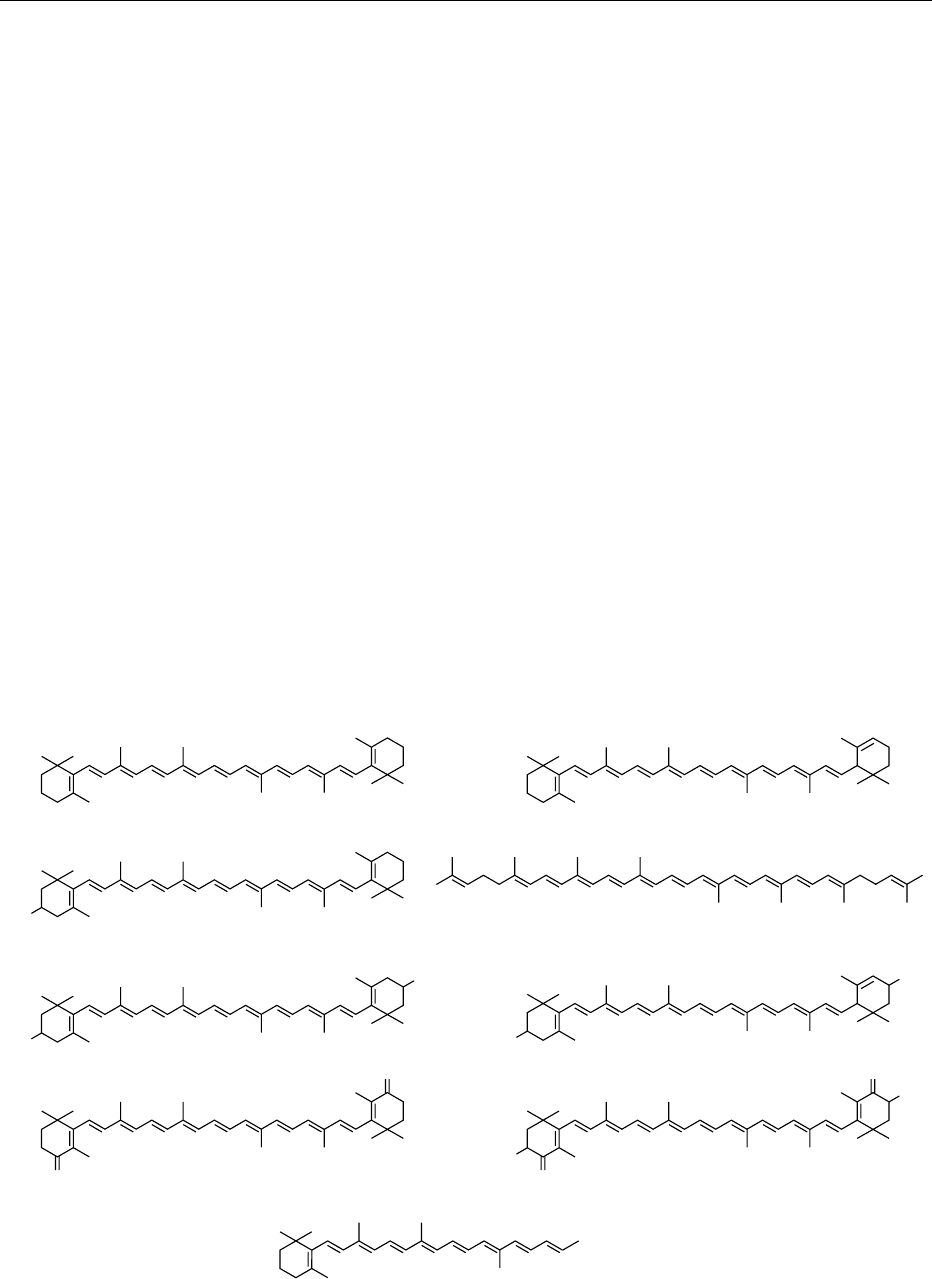
six-membered rings at each end of the polyprenoid
chain, although lycopene (with no end rings) is a
prominent exception (Figure 1). b-Carotene (C
40
H
46
,
MW 536), with two b-ionone rings connected by a
conjugated polyene chain, is the carotenoid proto-
type; formally, it can be considered to be composed
of eight isoprene units joined head-to-tail, except
in the center, so that the molecule is symmetrical about
the center. The large number of conjugated carbon–
carbon double bonds is responsible for carotenoids’
high light-absorbing ability and intense red–yellow
colors (wavelengths of maximum light absorption
are typically 450–500 nm, molar extinction coeffi-
cients typically 132 000–185 000 mol
1
l
1
). Effect-
ive photoprotection requires at least nine conjugated
double bonds, and absorption of visible light requires
seven to 13 conjugated double bonds. Higher carote-
noids (C
45
and C
50
), of bacterial origin, are products
of addition of C
5
units to give a ring structure substi-
tuted at the 2-position.
0002 The two major carotenoid subgroups are the caro-
tenes (hydrocarbon carotenoids, including lycopene,
a-carotene, and b-carotene) and the oxycarotenoids,
also called xanthophyls (hydoxy-, oxy-, epoxy-, and
furanoxy derivatives, such as lutein, canthaxanthin,
and astaxanthin) (Figure 1). The prefix ‘neo-’ was
previously used to designate carotenoids with at
least one cis configuration in the polyisoprene
chain, and the prefix ‘pro-’ designates some poly-cis-
carotenoids. The prefix ‘apo-’ designates a chain-
shortened structure (e.g., b-apo-8
0
-carotenal). Be-
cause the basic structures can be further metabolized
by the introduction of hydroxyl, keto, or methoxyl
groups, and by hydrogenation, cyclization, isomeri-
zation, and oxidative degradation, a great variety of
structures exist; it has been estimated that more than
600 carotenoids have already been identified in
nature. Good reviews on the structures and physical
properties of carotenoids have been provided by Isler
et al. and by Bauernfeind, and in a series edited by
Britton, Liaaen-Jensen, and Pfander.
0003Carotenoids are synthesized by higher plants and
by certain fungi, algae, and bacteria; they are not
synthesized by animals but may be biochemically
modified by them. Carotenoids are ubiquitous
in the plant kingdom (no naturally occurring
green plant lacks carotenoids), which suggests that
their two functions are irreplaceable: as protectants
against reactive singlet oxygen and as accessory
photopigments. In photoprotection, chlorophyl and
other cellular components are protected against
irreversible destruction by preferential reaction of
reactive singlet-state oxygen (electronically excited)
with carotenoids. As accessory photopigments,
CH
O
β-Carotene
β-Cryptoxanthin
β-apo-8⬘-Carotenal
Zeaxanthin
Canthaxanthin Astaxanthin
Lutein
Lycopene
α-Carotene
HO
HO
HO
HO
OH
OH
OH
O
O
O
O
..
fig0001 Figure 1 Structures of some representative carotenoids.
CAROTENOIDS/Physiology 937

carotenoids increase the efficiency of photosynthesis
by absorbing light in regions of the visible spectrum
where chlorophyls do not absorb efficiently, then
transferring the captured energy to other photopig-
ments; carotenoids in photosystems are associated
with proteins, and the carotenoid–protein inter-
actions can shift the wavelength of maximum light
absorption. Carotenoids are particularly effective in
both processes, because their singlet electronic states
are higher in energy, and their triplet states are lower
in energy than those of chlorophyls, thus allowing
both photoprotection and light-harvesting to be ener-
getically favorable.
0004 The total carotenoid production in the biosphere
has been estimated at 10
8
tonnes per year; this is
mostly due to fucosanthanin (characteristic of many
marine algae) and the main carotenoids of green
leaves, namely lutein, violaxanthin, and neoxanthin.
b-Carotene, although quantitatively less important,
occurs widely and is of particular interest as a
major precursor of vitamin A for animal and human
nutrition.
Absorption and Bioavailability of
Carotenoids
0005 There is considerable species variability in the effi-
ciency of absorption and in metabolism of carote-
noids. Humans are apparently unusual in absorbing,
transporting, and storing both carotenes and xantho-
phyls well. Rats do not accumulate b-carotene, but
instead metabolize almost all dietary b-carotene to
vitamin A. In contrast, cattle show serum and milk
accumulation of b-carotene but not of xanthophyls.
Animals with yellow fat absorb and store carote-
noids, whereas animals with white fat do not. Ferrets,
gerbils, preruminant calves, and nonhuman primates
have been used as animal models for human caroten-
oid absorption; rats and mice are also useful for
studies of noncleavable carotenoids and for studies
of pharmacological doses of carotenoids used for
cancer prevention or treatment. Some fish (for
example, salmonids) and birds absorb xanthophyls
particularly well and use them as pigments.
0006 Intestinal absorption of carotenoids in animals re-
quires the presence of bile acids; human subjects with
impaired bile flow (biliary atresia) show low levels of
liver carotenoids compared with normal subjects. The
presence of other dietary lipid promotes expansion of
the mixed bile salt micelle and enhances carotenoid
absorption. Animal and human feeding studies have
shown that the biological matrix of food affects
absorption of dietary carotenoids, and dietary caro-
tenoids in plant tissues have a lower bioavailability
than those in oil solutions. Hence, the plant source
and method of preparation of foods affect bioavail-
ability of b-carotene and other carotenoids: processes
that break down the biological matrix (e.g. grinding
or chopping, mild cooking) improve carotenoid ab-
sorption. Castenmiller and West (1998) have charac-
terized the factors affecting carotenoid absorption as
Species of carotene; Molecular linkage (e.g., esterifi-
cation of xanthophyls); Amount of carotene con-
sumed in a meal; Matrix in which the carotenoid
is incorporated; Absorption modifiers (including
lipid content of the meal); Nutrient status of host (in
particular, vitamin A status); Genetic factors; Host-
related factors; and Interactions (e.g., competition
among carotenoids for absorption).
0007Unequivocal estimation of the efficiency of absorp-
tion of dietary carotenoids is very difficult because of
the ubiquitous presence of carotenoids in the diet
(and thus in plasma and other tissues), slow absorp-
tion kinetics, partial metabolism of carotenoids, and
considerable individual variation in the extent of
absorption and metabolism.
Transport, Distribution, and Storage of
Carotenoids
0008As with other lipids, freshly absorbed carotenoids are
transported in the lymph via chylomicra. It is believed
that the carotenoids, as with other lipid components
of the chylomicron remnants, are taken up by
the liver and then released into the blood stream
as components of very-low-density lipoproteins
(VLDL). The VLDL are taken up by extrahepatic
tissues, their components processed into low-density
lipoproteins (LDL), and eventually into high-density
lipoproteins (HDL). Early studies showed that
in human plasma, the hydrocarbon carotenoids b-
carotene and lycopene are predominantly associated
with LDL (75%) and HDL (25%); the xanthophyl
lutein is approximately equally distributed between
HDL and LDL. In cattle, where the predominant
plasma lipoprotein is HDL, more than 80% of
total plasma b-carotene is transported in this fraction.
The predominant human serum carotenoids in the
USA are b-carotene, a-carotene, lycopene, b-cryptox-
anthin, lutein, and zeaxanthin, and also the colorless
carotenoids phytoene and phytofluene; serum carot-
enoid profiles and concentrations are highly depend-
ent on dietary intake and so depend on food habits. It
is estimated that serum carotenoids represent about
1% of total body carotenoids in the human.
0009Adipose tissue (80–85% of total body carotenoids)
and liver (8–12%) are the major sites of carotenoid
deposition in humans when the total mass of tissue
carotenoid is considered. Typical total human carot-
enoid levels of 100–150 mg are reported from
938 CAROTENOIDS/Physiology

autopsy analyses. Human adipose carotenoid profiles
are similar to those of serum. Reported values for
total human liver carotenoids range between 0 and
97 mg per gram of liver (i.e., 0–0.18 mmol g
1
; total
carotenoids were expressed as b-carotene in older
studies), and include lutein, lycopene, a-carotene,
and b-carotene. Other tissues that are known to
contain high concentrations of carotenoids include
adrenals (20 mgg
1
(37 nmol g
1
) tissue, mostly as b-
carotene) and human macular pigment (containing
predominantly the xanthophyls lutein and zeaxan-
thin), as well as kidney, ovaries, pituitary, placenta,
testes, and thymus in cattle. Chick retinal oil droplets
accumulate astaxanthin, which can be formed from
dietary zeaxanthin or canthaxanthin. It has been sug-
gested that organs with high numbers of LDL recep-
tors and high rates of LDL uptake show higher tissue
levels of carotenoids.
0010 The specific accumulation of certain carotenoids in
particular tissues may furnish clues to distinctive
physiological roles of carotenoids. The macula of
the primate eye accumulates the xanthophyls lutein
and zeaxanthin, with lutein being found predomin-
antly in the center of the macula; loss of these xantho-
phyls is associated with macular degeneration, a
disease characteristic of aging. Bovine corpus luteum
amasses xanthophyls (as much as 60 mg of total car-
otenoids per gram, 105 nmol per gram of tissue), and
it has been suggested that carotenoids play some im-
portant role in bovine reproduction; however, porcine
corpus luteum does not accumulate carotenoids.
Bovine pineal gland has been found to store b-carotene
(as much as 1 mg per gram, 2 nmol per gram of wet
tissue), with perhaps small amounts of xanthophyls
also present; the absence of other carotenoids was
noteworthy. Levels of approximately 1 mg of total
carotenoid per gram of bovine pituitary gland have
been reported. The physiological importance of these
accumulations of carotenoids is, however, not clear.
0011 Carotenoid–protein interactions have been stud-
ied in plants, bacteria, and marine invertebrates.
The strength of specific protein–carotenoid binding
depends, not surprisingly, on carotenoid structure.
Distinctive interactions between a binding protein
and its carotenoid ligand are evident by the spectral
changes of the carotenoid on binding; these spectral
changes may reflect physiological functions as well
as producing color polymorphism among species. A
notable example of these carotenoid–protein inter-
actions is the binding of the xanthophyl astaxanthin
(red-colored in solution) to the protein a-crustacyanin
in the lobster, giving a pigment that is typically blue–
green; on boiling the animal, the carotenoid is
released from its binding protein to yield a bright
red color.
Retinol Equivalents of Different
Carotenoids
0012Of the more than 600 known carotenoids, only about
60 have been reported to be precursors of vitamin A.
In order to serve as a precursor of vitamin A, a carot-
enoid must have at least one unsubstituted ‘b-ionone’
ring (2,6,6-trimethyl-1-cyclohexen-1-yl) with a poly-
ene side chain of at least 11 carbon atoms. Thus,
a-carotene (with one b-ionone ring) has half the
biological activity of b-carotene, and canthaxanthin
(with keto substitutions on both rings; Figure 1) has
no provitamin A activity. Absolute vitamin A activity,
however, depends on a number of other factors, not
all of which are well understood. In general, it seems
that the efficiency of the conversion of carotenoids to
vitamin A depends on the vitamin A status: high
intakes of preformed vitamin A result in poor effi-
ciency of conversion to vitamin A, perhaps due to
metabolic control of the cleavage enzyme(s). Frank
vitamin A deficiency is also associated with impaired
conversion efficiency, perhaps because of damaged
intestinal epithelial function in vitamin A inadequacy.
Because conversion of b-carotene to vitamin A is
impaired in vitamin A deficiency, vitamin A defi-
ciency disease is more rapidly cured by providing
preformed vitamin A than by providing dietary
carotenoids. Excessive vitamin E intake may impair
carotenoid cleavage (or interferes with intestinal ab-
sorption); vitamin E deficiency decreases vitamin A
formation, perhaps because adequate vitamin E is
needed to protect carotenoids and vitamin A from
oxidation. Conversion efficiency of b-carotene and
other carotenoids to vitamin A is also decreased at
high carotenoid intakes, perhaps due to impaired
intestinal absorption or to metabolic control of the
cleavage enzyme(s). Protein deficiency impairs carot-
enoid cleavage, suggesting that protein malnutrition
in humans exacerbates vitamin A deficiency.
0013Common dietary sources of provitamin A carote-
noids are carrots, yellow squash, dark-green leafy
vegetables, yellow maize, tomatoes, papaya, and
oranges. Cereal grains and white maize contain little
or no provitamin A. Red palm oil is one of the richest
sources of provitamin A carotenoids, containing
approximately 0.5 mg of a-carotene þb-carotene per
milliliter; as little as 7 ml of palm oil per day may
provide adequate vitamin A for the preschool child.
On a global basis, b-carotene is the most important
vitamin A precursor, both because of its greater pro-
vitamin A activity and because of its wide distribution
in plant products. The recent incorporation of the
enzymatic pathway of carotenoid biosynthesis into
rice (‘Golden Rice’) promises to help alleviate vitamin
A deficiency in parts of the world where rice is
CAROTENOIDS/Physiology 939

eaten extensively. Other provitamin A carotenoids,
such as a-carotene and b-cryptoxanthin and
b-apo-carotenals, can be nutritionally important
sources of vitamin A from particular foods (Table 1).
Other carotenoids, such as lycopene (which has no b-
ionone rings) and the dihydroxycarotenoids lutein
and zeaxanthin (which are substituted on both ionone
rings), may be major carotenoids in particular foods
and may have other important physiological func-
tions but have no provitamin A activity.
0014 Estimation of provitamin A content of foods from
food composition tables is problematic: carotenoid
composition of raw fruits and vegetables varies with
species, growth conditions, and mode of storage. Fur-
thermore, older tables often express total carotenoids
as ‘b-carotene,’ ignoring the different provitamin A
activity of different carotenoids. Biological activity
of carotenoids in foodstuffs may differ from the
analyzed content because of the mode of preparation,
binding of carotenoids within the foodstuff, and the
nature of the meal (presence or absence of fat), as
discussed above. Conversion factors, recently revised
to account for apparent bioavailability of carotenoids
from foods, are given in Table 2.
Enzymatic Conversion to Vitamin A
0015By comparison of the structure of b-carotene with
that of vitamin A, it would appear that b-carotene
could be cleaved in its center (between the 15 and 15
0
carbon atoms) to give two molecules of vitamin A per
molecule of carotenoid. Such a mechanism was first
proposed by Karrer. However, in addition to this
central (symmetric) cleavage, a random (asymmetric)
cleavage has been proposed. Glover first suggested
that asymmetric cleavage of the symmetric b-carotene
molecule could be followed by stepwise shortening
to vitamin A. In support of this hypothesis, small
tbl0001 Table 1 Vitamin A activity of common carotenoids relative to b-carotene
a
Carotenoid Activity (%) Occurrence (partiallist)
b-Carotene 100 Green plants, vegetables, carrots, yellow sweet potatoes,
squash, tomatoes, red and yellow fruits
a-Carotene 50–54 Green plants, carrots, squash, maize, green peppers
g-Carotene 42–50 Carrots, sweet potatoes, maize, tomatoes, algae, some
fruits
b-Carotene 5
0
,6
0
-monoepoxide 21 Plants, potatoes, red peppers
b-Carotene 5,6:5
0
,6
0
-diepoxide Active? Plants
b-Carotene 5
0
,8
0
-monofuranoxide (mutatochrome,
citroxanthin, flavacin)
50 Orange peel, red peppers, tomatoes, sweet potatoes,
cranberries
4-Keto-b-carotene (4-oxo-b-carotene, echinenone,
aphanin, myxoxanthin)
44–54 Algae, sea urchins, Daphnia, Hydra, red sponges, brine
shrimp, crustaceans
3-Keto-b-carotene (3-oxo-b-carotene) 52
3-Hydroxy-b-carotene (b-cryptoxanthin) 50–60 Yellow maize, green peppers, lichens, persimmons,
papayas, lemons, oranges
4-Hydroxy-b-carotene (isocryptoxanthin) 48 Brine shrimp
3,4-Dehydro-3
0
-hydroxy-b-carotene (anhydrolutein,
deoxylutein)
Active, c. 10 Alfalfa meal, acidulated soybean soapstock
3-Hydroxy-4-keto-b-carotene (hydroxyechinenone) Probably active Algae, bacteria, flowers
b-Apo-2
0
-carotenal Active Citrus fruit
b-Apo-8
0
-carotenal 36–72 Citrus fruit, green plants
b-Apo-10
0
-carotenal Active Citrus fruit, green plants, alfalfa meal
b-Apo-12
0
-carotenal 120 Alfalfa meal
b-Apo-8
0
-carotenoic acid Active Maize, animal tissue
b-Apo-8
0
-carotenoic acid ethyl ester 25–78
Lycopene Inactive Tomatoes, carrots, green peppers, pink citrus fruit
3,3
0
-Dihydroxy-b-carotene (zeaxanthin) Inactive Spinach, paprika, yellow maize, green peppers, fruits
3,3
0
-Dihydroxy-a-carotene (lutein) Inactive Green leaves, yellow maize, potatoes, spinach, green
peppers, carrots, tomatoes, fruits
3,3
0
-Dihydroxy-4,4
0
-diketo-b-carotene (astaxanthin) Inactive Oranges, crustaceans, lobster, fish, algae, Daphnia, trout,
salmon
4,4
0
-Diketo-b-carotene (canthaxanthin, aphanicin) Inactive Mushrooms, trout, Daphnia, Hydra, microorganisms,
algae, crustaceans, brine shrimp
Capsanthin Inactive Red peppers, paprika
Capsorubrin Inactive Red peppers, paprika
Bixin Inactive Annatto seeds
a
Adapted and shortened from Bauernfeind JC, Adams CR and Marusich WL (1981) Carotenes and vitamin A precursors in animal feed. In: Bauernfeind JC
(ed.) Carotenoids as Colorants and Vitamin A Precursors, pp. 563–743. New York: Academic Press.
940 CAROTENOIDS/Physiology
amounts of labeled b-apo-8
0
-, b-apo-10
0
-, and b-apo-
12
0
-carotenal (and their alcohols and acids) have been
found in rat tissues after feeding radioactive b-carot-
ene. These apo-carotenals can be formed by chemical
reaction of oxidizing agents (hydrogen peroxide, po-
tassium permanganate) with b-carotene in vitro, and
they are found in small amounts in plants and in
animal-feeding experiments. Enzymatic asymmetric
cleavage activity has been demonstrated in vitro, but
its relative importance has not been established.
However, in vitro experiments (initially carried out
by Olson and Hayaishi and by Goodman and Huang)
using partially purified intestinal homogenates (from
rat, rabbit, or guinea-pig) have demonstrated sym-
metric cleavage of b-carotene, with retinal (vitamin
A aldehyde) as the only detectable product. This en-
zymatic activity requires the presence of O
2
, and has
been named ‘15,15
0
- b-carotenoid dioxygenase’ (EC
1.13.11.21). The enzyme has recently been cloned
from mammalian sources, and its further character-
ization promises to explain much about the factors
affecting provitamin A activity of b-carotene and re-
lated carotenoids. This enzymatic activity has been
demonstrated in intestine, liver, kidney, and several
other tissues in vitro. b-Carotene is not a source of
vitamin A in the cat, and it has been reported that
15,15
0
-dioxygenase activity is not present in cat intes-
tine, consistent with the ‘central cleavage’ hypothesis.
The possiblity remains that excentric cleavage is a
source of retinoic acid, which is an important modu-
lator of gene expression.
001 6 Some fish and birds can also convert the xantho-
phyls astaxanthin, canthaxanthin, and isozeaxanthin
to vitamin A; some freshwater fish can convert
lutein to 3,4-didehydroretinol (vitamin A
2
). Keto
carotenoids can be reduced to the corresponding alco-
hols and esterified with long-chain fatty acids.
Toxicity of Carotenoids
0017There is virtually no toxicity associated with high
intakes of carotenoids, in marked contrast to vita-
min A. Excessive human ingestion of b-carotene can
cause high serum carotenoid levels and yellowing of
light-colored skin, but such over-intake is very
unusual, and no harmful physiological effects from
even such high intakes of b-carotene have been
reported. The effects disappear when high carotenoid
intakes cease. Ingestion of large amounts of carote-
noids does not give rise to hypervitaminosis A, prob-
ably because of the decreased efficiency of conversion
of carotenoids to vitamin A at high intakes. Terato-
genicity due to carotenoids has never been suggested.
b-Carotene supplementation in humans does not
produce elevated serum triacylglycerols or choles-
terol. However, prolonged intake of high doses
(20 mg per day) of b-carotene has been associated in
epidemiologic studies with increased risk of lung
cancer in cigarette smokers, perhaps because of the
prooxidant effects of b-carotene at relatively higher
oxygen concentrations in the lungs. Although inges-
tion of 3–10mg of carotenoid per day from foods and
dietary supplements appears to be completely safe
and advisable, there are concerns that higher doses
of b-carotene (20–30 mg per day) may not be safe in
smokers.
0018The only carotenoid that has been clearly associated
with adverse effects is canthaxanthin (4,4
0
-diketo-b-
carotene), a carotenoid that has been used in treat-
ment of patients with erythropoietic protoporphyria
tbl00 02 Table 2 ConversionfactorsforestimatingvitaminAvaluefromcarotenoidcomposition
a
Factors
1retinolactivityequivalent(RAE)
¼1 mgall-transretinol
¼12 mgall-trans b-carotene
¼24 mgotherprovitaminAcarotenoids
¼3.33IUvitaminA
(Notethat1RAE¼1.15 mgofretinylacetate(MW328)¼1.83 mgofretinylpalmitate(MW524),i.e.,molarequivalence)
1InternationalUnitofvitaminA(IU
A
)
¼0.3 mgall-transretinol
¼0.3RAE
¼3.6 mgall-trans b-carotene
¼7.2 mgotherprovitaminAcarotenoids
Formulas
(1)Retinoland b-carotenegivenin mg,RAE¼ mgretinolþ(mg b-carotene/12)
(2) b-CaroteneandotherprovitaminAcarotenoidsgivenin mg,RAE¼(mg b-carotene/12)þ(mgotherprovitaminAcarotenoids/24)
a
AdaptedfromOlsonJA(1987)Recommendeddietaryintakes(RDI)ofvitaminAinhumans:Appendix.AmericanJournalofClinicalNutrition45:704–716
and from Panel on Micronutrients, Subcommittees on Upper Reference Levels of Nutrients and of Interpretation and Use of Dietary Reference Intakes,
and the Standing Committee on the Scientific Evaluation of Dietary Reference Intakes (2001) Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic,
Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon,Vanadium, and Zinc. Washington, DC: National Academy Press. Available
onlineathttp://books.nap.edu/.
CAROTENOIDS/Physiology 941

and related skin disorders. High daily doses
(50–100 mg) have resulted in canthaxanthin retino-
pathy (crystalline deposits in the retina leading to
impaired night vision); the effect is slowly reversible
on cessation of intake. Pharmacological doses of
canthaxanthin have depressed liver accumulation of
dietary vitamin A in rats. Human ingestion of large
amounts of canthaxanthin gives a yellowish color-
ation to light-colored skin; this is the basis of some
‘skin tanning’ pills.
Therapeutic Role of Carotenoids
0019 In addition to the important role of some carotenoids
as vitamin A precursors, there is evidence that carote-
noids can quench singlet oxygen and can serve as rad-
ical-trapping antioxidants; it has also been shown that
carotenoids may have a role (apart from being precur-
sors of retinoids) in enhancing the immune function in
animals. Singlet-oxygen trapping and antioxidant
capabilities are shown by carotenoids that have no
provitamin A activity (such as lycopene and astax-
anthin) as well as by provitamin A carotenoids.
0020 Both b-carotene and canthaxanthin have been used
in large dose regimens to ameliorate photosensitivity
associated with erythropoietic protoporphyria (a
genetic disease of porphyrin metabolism). b-Carotene
also prevents photosensitivity resulting from quini-
dine ingestion, but not photosensitivity to ultraviolet
light, although it has been reported that human serum
carotenoid levels decrease after repeated exposure to
ultraviolet light.
0021 Epidemiological evidence strongly suggests a
role for carotenoids in human cancer prevention,
although such studies may be confounded by other
factors. The strongest evidence is for a protective
effect of carotenoids against lung cancer; tomato
products (which provide lycopene) may be more
effective than carrots and squash (sources of
b-carotene). In particular, consumption of tomato
products (high in lycopene) has been associated epi-
demiologically with a reduced risk of prostate neo-
plasms. Protective effects of carotenoids against
laryngeal cancer, gastric cancer, invasive bladder
cancer, and cervical dysplasia and cervical cancer
have been suggested also; these effects are not
shown by vitamin A, suggesting a direct role of car-
otenoids. Neither carotenoids nor vitamin A protect
against cancers of the esophagous or gastrointestinal
tract, breast, head, or neck. A variety of cancers in
animal models are prevented or delayed by carote-
noids, including skin tumors and mammary tumors.
In studies in cells in culture, carotenoids can prevent
malignant transformation, sister chromatid ex-
change, and mutagenic effects in bacterial systems.
0022A high consumption of carotenoid-containing
foods is associated with a reduced risk of coronary
heart disease, although there is a lack of consistency
in identifying the responsible compounds.
0023Carotenoids have been found to enhance both
specific and nonspecific immune functions, including
proliferation of T- and B-lymphocytes, induction of
specific effector cells capable of killing tumor cells,
and increased production of tumor necrosis factor,
and secretion of factors required for communication
between cells (prostaglandins and leukotrienes). It
has been suggested that the mechanism(s) may
involve quenching singlet oxygen and trapping free
radicals (generated by neutrophils to kill invading
cells), or maintaining cell-membrane fluidity (import-
ant for the function of membrane receptors and for
the release of immunomodulating factors). Again,
some of the immunologic functions of carotenoids
seem to be independent of any provitamin A activity,
although retinoids are also important in immune
function.
0024The xanthophyls lutein and zeaxanthin are accu-
mulated specifically in the macula of the primate eye,
and high dietary intakes of these two carotenoids (and
elevated plasma concentrations) are associated with a
reduced risk of age-related macular degeneration.
0025Elucidation of these biochemical effects of carote-
noids and further explanation of the metabolism
of conversion of carotenoids to vitamin A remain
central issues in the study of carotenoid function.
See also: Carotenoids: Occurrence, Properties, and
Determination; Protein: Deficiency; Retinol: Properties
and Determination; Tocopherols: Properties and
Determination
Further Reading
Bauernfeind JC (ed.) (1981) Carotenoids as Colorants and
Vitamin A Precursors. New York: Academic Press.
Beyer P, Al-Babili S, Ye X et al. (2002) Golden Rice: Intro-
ducing the beta-carotene biosynthesis pathway into rice
endosperm by genetic engineering to defeat vitamin A
deficiency. Journal of Nutrition 132: 506S–510S.
Britton G, Liaaen-Jensen S and Pfander H (eds) (1995a)
Carotenoids. 1A. Isolation and Analysis. Basel: Birk-
hauser.
Britton G, Liaaen-Jensen S and Pfander H (eds) (1995b)
Carotenoids. 1B: Spectroscopy. Basel: Birkhauser.
Castenmiller JJ and West CE (1998) Bioavailability and
bioconversion of carotenoids. Annual Review of Nutri-
tion 18: 19–38.
Isler O, Gutmann H and Solms U (eds) (1971) Carotenoids.
Basel: Birkhauser Verlag.
Lee CM, Boileau AC, Boileau TW et al. (1999) Review of
animal models in carotenoid research. Journal of Nutri-
tion 129: 2271–2277.
942 CAROTENOIDS/Physiology

Straub O (1987) In: Pfander H (ed.) Key to Carotenoids.
Basel: Birkhauser.
Subcommittees on Upper Reference Levels of Nutrients and
Interpretation and Uses of Dietary Reference Intakes,
and the Standing Committee on Scientific Evaluation
of Dietary Reference Intakes (2000) Dietary Reference
Intakes for Vitamin C, Vitamin E, Selenium and
Carotenoids: A Report of the Panel on Dietary Antioxi-
dants and Related Compounds. National Academy
Press, Washington, D.C.
Von Lintig J and Wyss A (2001) Molecular analysis of
vitamin A formation: cloning and characterization
of beta-carotene 15,15
0
-dioxygenase. Archives of Bio-
chemistry and Biophysics 385: 47–52.
CASEIN AND CASEINATES
Contents
Methods of Manufacture
Uses in the Food Industry
Methods of Manufacture
C R Southward, Fonterra Research Centre,
Palmerston North, New Zealand
This article is reproduced from Encyclopedia of Dairy Sciences,
Copyright 2002, Academic Press.
Introduction
0001 The commercial production of casein, the principal
protein in cows’ milk, has occurred for most of the
20th century. During the period to 1960, the major
proportion of casein was used in technical (or non-
food) applications. More recently, however, there has
been a significant change from technical to edible uses
for casein products and this has been reflected in the
introduction of requirements for pasteurization of
milk intended for casein manufacture (c. 1970) and
the greater number of specifications for microbial
quality and freedom from impurities. At the present
time, the major producers of casein include the Euro-
pean Community (in particular, Denmark, France,
Germany, the Irish Republic, and The Netherlands)
and New Zealand, with Poland, Australia, and India
producing smaller quantities. World production of
casein products is currently estimated at 220 000–
250 000 tonnes. This article describes the manufac-
ture of the main casein products.
General
0002 The proteins that exist in milk can be broadly divided
into two groups – casein and whey proteins. Caseins
may be considered as those proteins that are precipi-
tated when unheated (raw) milk is acidified to pH 4.6
(the isoelectric point of casein) whereas whey proteins
remain in solution. Commercial casein is a mixture of
four different caseins – a
s1
-, a
s2
-, b- and k-casein, and
may sometimes be referred to as whole casein. As a
phosphoprotein, casein belongs to a relatively rare
class of proteins. It contains 0.7–0.9% phosphorus,
covalently bound to the casein by a serine ester
linkage. Casein exists in milk in combination with
calcium, inorganic phosphate, and citrate as a col-
loidal suspension of complex micelles and accounts
for 2.6–2.9% by weight of whole milk.
Manufacture
0003Casein may be precipitated from skimmed milk to
produce several products such as acid casein, rennet
casein, or coprecipitate. All these products are
insoluble in water after precipitation. However,
addition of alkali to acid casein yields water-soluble
caseinate.
Acid Casein
Acidification
0004When milk (pH 6.6) is acidified, the calcium and
inorganic phosphate are removed from the casein
micelles, the net charge on the micelles decreases,
and the micelles become less and less stable until the
casein precipitates. Complete precipitation of the
casein occurs at the isoelectric point, pH 4.6. Acidifi-
cation of the milk may be carried out by one of the
following processes:
1.
0005Inoculation of milk with lactic acid-producing
bacteria such as Lactococcus lactis subsp. lactis
CASEIN AND CASEINATES/Methods of Manufacture 943

or cremoris. These bacteria, commonly known as
‘starters,’ convert some of the lactose in the milk
to lactic acid during the period of incubation
(about 16–18 h). This is the most commonly
employed method of manufacture in New Zealand
(Figure 1).
2.
0006 Direct addition of dilute acid to skimmed milk.
Hydrochloric acid, sulfuric acid, phosphoric acid,
or lactic acid (or, occasionally, other organic acids)
may be used for this purpose. The most common
method of producing acid casein in countries other
than New Zealand is by means of hydrochloric
acid.
3.
0007 Indirect acidification of skim milk. A number of
alternative processes have been patented in which
skimmed milk is acidified by one or more of the
following:
.
0008 contact by means of ion exchange resins in the
acid (cation) form
.
0009 addition of acidified whey
.
0010 electrodialysis
It is believed that these processes have relatively
minor commercial significance.
Cooking/Acidulation
0011 In the manufacture of acid casein, acidification of the
milk is followed, or occasionally preceded, by heating
of the mixture. Heating promotes agglomeration of
the casein curd particles which subsequently shrink
(syneresis) to expel whey. At the same time, the curds
become firmer and are able to withstand the mechan-
ical processing that follows. Heating of the acidified
milk is sometimes termed ‘cooking’ (as used in cheese
manufacture) and may be carried out, usually in the
temperature range 45–55
C, by:
1.
0012 injection of steam into the pipeline carrying the
acidified milk
2.
0013 indirect heating by means of a heat exchanger
3.
0014 a combination of both – preheating through a heat
exchanger with steam injection to complete the
heating process (Figure 1)
In each case, the cooked curds and whey are held in a
‘cooking pipe’ for a period of about 10–20 s before
they emerge into an acidulation vat. The curds and
whey may remain here for a period varying from 30 s
to about 15 min, during which time the curds are
agitated gently in the whey until equilibrium between
the calcium in the curds and that in the whey is
attained.
0015 Alternative processes may use a syneresis tube; the
cooked curds and whey are held in a large-diameter
tube for a period of several minutes (also undergoing
acidulation or equilibration).
Dewheying and Washing
0016Following acidulation, the curds and whey are
discharged by gravity to a dewheying screen above
the first washing vat. The whey is removed and the
curds fall into the vat. Alternatively, the curds and
whey may be separated more completely using a hori-
zontal solid-bowl centrifuge (decanter) or a casein-
dewatering press before the curds are transferred to
the first wash. A combination of screening and decanter
dewheying can be used to reduce the hydraulic load on
the decanter. The purpose of washing is to remove
whey (containing mainly lactose) from the curds so
that the casein produced is relatively pure. The tem-
perature of the wash water may be varied, depending
upon particular requirements. Casein is usually sub-
jected to multiple washes and these are operated in a
counterflow to the direction of the curds, with the
purest curd meeting the cleanest water.
Dewatering
0017After washing, the curds are mechanically dewatered
to remove excess water before drying. As drying is a
relatively expensive operation, it is worthwhile re-
moving as much water as possible from the curd
before transferring the casein to the drier. The texture
of the curd is affected by temperature; as the tempera-
ture of the wash water is increased, the curd releases
more water during dewatering but becomes firm and
more plastic and is consequently harder to break up
and dry. Therefore, it is necessary to regulate carefully
the temperature of the last wash to optimize the con-
flicting requirements of minimum water content and
maximum friability of the curd.
0018Equipment for dewatering casein curd consists of
roller or belt presses, decanters, and screen-bowl cen-
trifuges. The roller press, used for many years, is
designed to reduce the moisture content of curd to
about 55%. The belt press will do a similar duty.
Solid-bowl decanters or screen-bowl centrifuges are
capable of reducing the moisture content of acid
casein curd to about 52%.
Drying
0019Drying of casein curd is most commonly carried out
using horizontal vibrating fluid-bed driers. These
driers have two or more perforated stainless-steel
decks (Figure 1). The combined effect of vibration
of the decks and the flow of hot air (typical tempera-
ture range 75–115
C) up through the holes in the
decks causes the casein curd to become fluidized and
materially helps in the removal of moisture from the
particles. Most of the water is removed during
the early stages of drying of the casein as it is evapor-
ated from the surface of the particles. The later stages
944 CASEIN AND CASEINATES/Methods of Manufacture
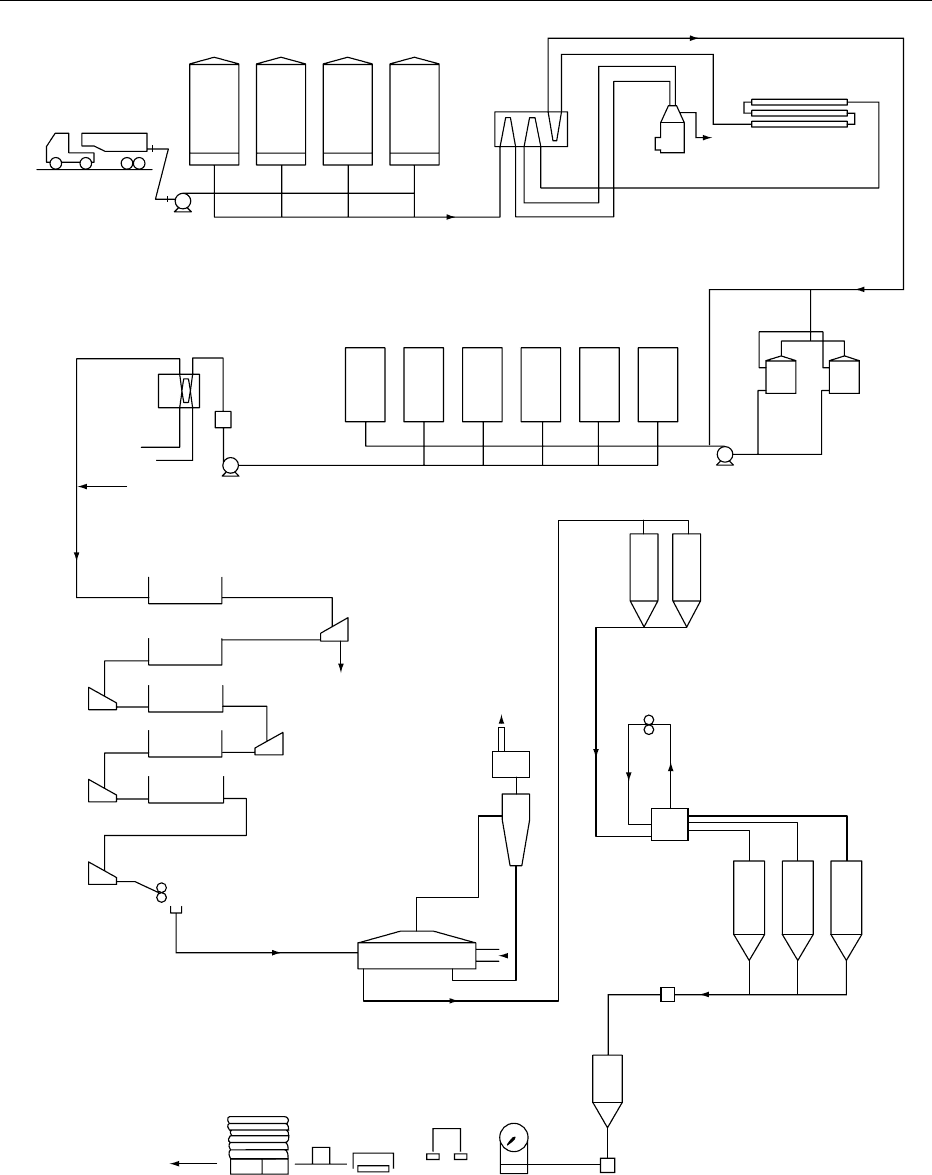
Tanker
reception
Whole milk storage
Wm
Unloading Pasteurizer
Preheater
Steam injector
Cas
Screen
Acidulation
Washing and
separation
by screening
Cas
Cas
Press
Casein drier
Fines recovery
cyclone
Whey
Exhaust
Tempering
Roller mill
Oversize
particles
Cas
Blending
80 mesh
60 mesh
30 mesh
Sifter
Metal
removal
Storage silos
Bag-filling
hopper
Bag
sewing
Metal
detector
Palletized bags
to store
Bag
flattening
Weighing
Coagulation silos
Separator
Sm
Sm
Sm
Co
Co
Flow
control
Starter
vessels
Cream
Wm
Holding
tubes
fig0001 Figure 1 Outline of the manufacturing steps involved in producing lactic acid casein from skim milk. Wm, whole milk; Sm, skimmed
milk; Co, coagulum; Cas, casein.
CASEIN AND CASEINATES/Methods of Manufacture 945