Caballero B. (ed.) Encyclopaedia of Food Science, Food Technology and Nutrition. Ten-Volume Set
Подождите немного. Документ загружается.

to their respective receptors along the intestinal wall
reducing absorption. The results of a recent meta-
analysis using several different clinical studies showed
a statistically significant lowering of plasma triacyl-
glycerides and cholesterol in subjects fed for at least
14 days with a diet supplemented with oats, psyllium,
pectin or guar gum, but the effect was modest: a
decrease of approximately 1 mg per deciliter per
gram of soluble fiber was observed. If we consider
that an individual with moderately higher blood
cholesterol has a total serum cholesterol value of
approximately 300 mg dl
1
, the change brought
about from consumption of viscous, soluble fibers is
on the order of 0.3%. Considering that oat bran
contains about 20% fiber by weight and only half of
this is soluble, one can calculate that 10 g of oat bran
per day is needed for each 0.3% change in total
cholesterol.
0014 Insoluble dietary fiber is, by definition, the fraction
of total dietary fiber (TDF) that is not soluble in hot
buffer solution. However, the measured proportion of
insoluble fiber can vary depending on the dietary fiber
methodology. Insoluble dietary fiber consists primar-
ily of hemicellulose, cellulose, and lignin (See Dietary
Fiber: Determination). It is often incorrectly believed
that insoluble fiber is not fermented to any great
extent by the bacteria present in the large bowel.
However, actual measurements of fiber fermentabil-
ity have shown that a significant proportion of insol-
uble dietary fiber is fermented. This is true even for
wheat bran, which has a relatively lower insoluble
fiber fermentability of between 30 and 40% (ap-
proximately 90% of the wheat bran TDF is insoluble
dietary fiber). The fermentability of oat bran is even
greater – up to 80% of the insoluble fiber (approxi-
mately half of the TDF) is fermented in the human
large intestine. The degree of fermentability is a prop-
erty unique to each dietary fiber and depends largely
on the nature and the structural arrangement of the
fiber components, and also on other physical charac-
teristics such as particle size. For example, the struc-
ture of coarse wheat bran favors a high water-holding
capacity (see below) and a slow rate of fermentation.
This permits the microtrapping of slowly released
fermentation gasses, increasing fecal bulk and stimu-
lating defecation.
Water-holding Capacity
0015 Dietary fiber holds water by adsorption and absorp-
tion. Some water is also retained outside the fiber
matrix (free water). The particle size, chemical com-
position, and structure of dietary fiber influence the
water-holding capacity. The in vitro water-holding
capacity cannot be used to predict the impact of
highly fermentable fiber on colonic function.
0016Although lettuce fiber isolates retain much water,
the water-holding capacity expressed in grams of
water held by fiber per gram of edible portion of
food has been found to be higher for wheat bran
fiber (4.5 g) and carrot fiber (2.1 g) than for that of
lettuce and various other foods (0.3–1.3 g). The
water-holding capacity of fiber from the brans of
ready-to-eat breakfast cereals is positively correlated
(r 0.85, P < 0.05) with its mean particle size (MPS):
the water-holding capacity of 160-mm MPS wheat
bran fiber is 59% of that of 800-mm MPS wheat bran
fiber (Table 2). A large portion of the water held by
wheat bran fiber appears to be free water.
0017The water-holding capacity has a significant effect
on fecal output and stool hardness, as discussed
above. These factors are important physiological
effects of wheat bran and contribute significantly to
its laxative effects.
Bile Salt Binding and Serum Cholesterol Reduction
0018There is considerable interest in the cholesterol-
reducing properties of dietary fiber. This is because
the influence of serum cholesterol (low-density lipo-
protein (LDL) and high-density lipoprotein (HDL)
cholesterol, in particular) on cardiovascular disease
is considerable. Three different mechanisms have
been proposed to account for the serum cholesterol-
reducing properties of dietary fiber. As discussed
above, a certain soluble viscous fiber may modestly
lower cholesterol by increasing luminal viscosity and
preventing cholesterol (re)absorption. Acetic and pro-
pionic acids, produced by the fermentation of dietary
fibers in the gut, are thought to inhibit liver choles-
terol synthesis at the metabolic level. Dietary fibers
also have the ability to bind cholesterol directly:
deconjugated bile salts are bound to pectic substances
by hydrogen bonding, and lignin appears to bind bile
salts through hydrophobic interactions. The actual
tbl0002Table 2 Effect of grinding on the properties of wheat bran
insoluble fiber
a
Mesh sieve Aperture
b
(mm) MPS (mm)
c
WHC
d
Glycocholate
binding
e
As is 800 9.5 + 0.1 26.5 + 1.5
20 840 420 8.1 + 0.1 24.9 + 1.2
40 420 280 6.6 + 0.2 23.8 + 0.8
60 250 180 5.8 + 0.1 22.1 + 1.0
80 175 160 5.6 + 0.1 21.5 + 0.3
a
Residue after neutral detergent and porcine pancreatic a-amylase
treatments.
b
Aperture size of sieve used with a Wiley mill, intermediate model.
c
MPS, geometric mean particle size.
d
WHC, water-holding capacity, grams of water per gram of insoluble fiber,
using the centrifugation method: mean + SEM of three measurements.
e
Grams of glycocholate bound per 0.2 g of insoluble fiber; mean + SEM of
four measurements.
1816 DIETARY FIBER/Properties and Sources

in vivo mechanism is unknown, and it is likely that
different fibers operate through different combin-
ations of all three mechanisms to influence blood
cholesterol levels.
0019 In vitro evidence suggests that the physical form of
the fiber may also play an important role in binding
bile salts. In commonly ingested ready-to-eat cereals,
glycocholate (r ¼0.90, P < 0.001) and taurocholate
(r ¼0.86, P < 0.05) binding have been positively cor-
related with the MPS of the neutral detergent fiber
(NDF; see Determination). Reducing the particle size
of wheat bran NDF from 800 to 160 mm reduces its
glycocholate binding by 19% (Table 2). The bile salt
binding capacity of fiber isolates, although method-
dependent, may reflect events in the terminal small
intestine. Purified fiber fractions may be poor
(cellulose) or strong (lignin) binders. Some rice
brans appear to have a high capacity to bind bile salts.
0020 Human cholesterol-lowering studies have linked
the consumption of dietary fiber with lower total
plasma cholesterol and increased LDL/HDL ratios
in addition to reductions in serum triglycerides. Stud-
ies such as these have suggested that cardiovascular
risk may be reduced by consuming particular types of
foods high in dietary fiber, most notably whole cereal
grains. However, it should be noted that coronary
heart disease (CHD) is a complex disease and is
affected by many factors. For example, the reduction
in CHD observed in a recent study was larger than
would be expected from the beneficial effects of sol-
uble dietary fiber on serum cholesterol levels only.
Thus, reductions in CHD are likely to be the result
of many factors, one of which is total dietary fiber
intake.
Cation-exchange Capacity
0021 Cation exchange is partly dependent on the presence
of uronic acid in the nonesterified form. The prepar-
ation of the fiber material may decrease the number
of nonesterified carboxyl groups and the apparent
cation exchange capacity. Wheat bran contains little
uronic acid, and its cation exchange capacity is
mainly due to diffusion within the dietary fiber net-
work. The cation exchange capacity of fiber from
different sources is difficult to compare, unless it is
expressed per edible portion. According to the affin-
ity of minerals for carboxylic acid groups, cabbage
and coarse wheat bran show a high cation-exchange
capacity compared with pectin.
0022 There has been some concern that the ability of
dietary fiber to bind minerals (as measured in vitro)
may lead to mineral deficiencies in individuals con-
suming high-fiber diets. In North American diets,
calculations show that mineral intakes far exceed
the potential binding capacity of dietary fiber so
that no need for concern arises. In addition, minerals
that are bound to fibers or that are trapped in the (as
yet, undigested) cell-wall matrix may not be absorbed
in the small intestine but could be partially released
and absorbed in the colon when the fiber is degraded
by bacteria. Absorption of minerals in the colon has
been suggested as a mechanism for accounting for
increased mineral absorption in rats fed fructooligo-
saccharides. This may explain why cation adsorption
has not been consistently related to mineral bioavail-
ability. However, wheat bran fibers can permanently
bind heavy metal ions to decrease their toxicity.
Viscosity and Gelling Properties
0023As mentioned above, certain soluble fibers, such as
oat b-d-glucans, are viscous when dissolved in water,
while others, such as pectins, show gelling properties.
These fibers influence gastric emptying and absorp-
tion rates in the small intestine. Direct measurement
of the viscosity produced by concentrations of fiber
likely to be used in diets has shown little effect of low
methoxy pectin, a slight increase with wheat bran,
and a significant increase with high methoxy pectin.
Some of the nonstructural dietary fiber components
can also increase viscosity.
Microbial Degradation (Fermentability)
0024Most dietary fiber remains undegraded until it reaches
the large intestine, where the extent of fermentation
depends on the source as well as several other factors,
including the physical structure of the fiber, the
presence of specific components in the fiber matrix,
the nitrogen source, bacterial adaptation, and transit
time. It is generally accepted that soluble fiber is
almost completely fermented in the large intestine,
but unfortunately, it is also generally believed that
insoluble fiber is not fermented. Actually, both
insoluble and soluble fiber are extensively fermented.
On average, 70–80% of the total dietary fiber from
mixed diets (e.g., fruits, vegetables, legumes, and
most cereals) is degraded by colonic bacteria. Since
insoluble dietary fiber is the major dietary fiber
fraction, representing approximately two-thirds of
total dietary fiber in Western diets, this means that
insoluble fiber is highly fermented. Indeed, as indi-
cated above, 70–80% of the insoluble fiber from oat
bran and up to 40% of the insoluble dietary fiber
from wheat bran are fermented.
0025There is some indication that fermentability
depends on particle size, with small particle sizes
being more readily fermented, but this has not been
consistently reported, and the effect appears modest.
In rats fed purified diets containing 15% hard red
wheat bran, NDF fermentability was largely un-
changed (34–36%) when the MPS varied from 1275
DIETARY FIBER/Properties and Sources 1817

to 394 mm. Similar trends were observed when Ameri-
can Association of Cereal Chemists soft and hard
wheat brans were fed at various MPS. In these experi-
ments, 37.3–37.6% and 30.0–32.8% of the NDF was
fermented, respectively.
Interaction between Structure and
Physiology
Food Processing
0026 The physicochemical properties of dietary fiber per se
are dependent on the chemical composition and the
structural characteristics of the fiber. However, as
noted above, knowing the former does not allow
one to predict the latter. When considering the poten-
tial in vivo effect of dietary fiber, one must also take
food-processing practices into consideration. Many
different types of food processing practices are
currently employed. Foods can be boiled, canned,
frozen, blanched, parboiled, extruded, adiabatically
extruded, and milled. All processes can have an effect
on dietary fiber content, as measured by current
methodologies. For example, heat treatment can
break glycosidic linkages in dietary fiber polysacchar-
ides. This can solubilize some of the insoluble fiber as
well as reduce the TDF content if the polymers are
broken down into small molecular mass fragments.
Heat treatment can also reduce the overall length of
the polysaccharide chains to lower viscosity and the
water-holding capacity of the fiber. The pentosans in
dietary fiber can react with amino acids such as lysine
(Maillard reaction) and form new polymers that do
not behave physiologically like dietary fiber. The
boiling of vegetables increases the apparent TDF con-
tent, but this seems to be due to the loss of nonfiber
components to the water.
0027 It is often difficult to predict before hand the effect
of a food-processing method on TDF and the distri-
bution between insoluble and soluble fractions. For
example, extrusion cooking of some cereal brans ap-
parently increases its water-holding capacity in vitro,
although this does not translate into a change in fecal
water holding capacity in vivo. Extrusion cooking of
wheat bran causes small decreases in TDF and insol-
uble dietary fiber but also gives rise to small increases
in soluble dietary fiber. Processing of rice (abrasion or
parboiling) removes the seed coat and the majority of
the dietary fiber. Some collapsing of the fiber struc-
ture also occurs during food preparation and/or mas-
tication due to the reduction of particle size, and the
fiber structure of sources such as spinach may be
modified by cooking.
0028 Milling is known to significantly affect TDF fer-
mentation in the human colon. Cereal grains have a
tough outer seed coat that would be completely resist-
ant to digestion if the cereals were not milled prior to
consumption. After milling, the thin-walled dietary
fiber structures from the endosperm are partly dis-
rupted and, thus, easily broken during fermentation.
Milling, therefore, makes the endosperm accessible to
the digestive enzymes in the upper digestive tract but
also makes the dietary fiber more accessible to co-
lonic bacteria. However, milling can also negatively
affect the physiological properties of the dietary fiber
in grains. While coarse grinding and adequate masti-
cation appear to be sufficient for digestion of cereals,
fine grinding disrupts much of the cell-wall architec-
ture, and further milling can even interfere with its
determination by gravimetric methods. Table 3 shows
the effect of milling on the dry bulk volume of wheat
bran, as measured in a volumetric cylinder. The
volume of the coarse bran was reduced from
5mlg
1
to less than 2 ml after grinding in a M5
Wiley mill using a 0.5-mm screen. Fiber from differ-
ent sources ground under the same conditions may
behave differently during milling, so that their
resulting particle sizes may be different.
0029As discussed above, the particle size of unfer-
mented wheat bran fiber may be related to colonic
function and fecal characteristics in humans and
laboratory animals. Finely ground cellulose or even
wheat bran, for example, may have a constipating
effect, whereas coarsely ground wheat bran prevents
constipation. The influence of particle size on fecal
bulking has been measured directly in rats. When rats
were fed diets containing 12% hard wheat bran, the
fecal wet density was 0.796 + 0.010 (mean + SEM,
n ¼12) in the coarse bran group (geometric MPS:
850 mm) but increased to 0.888 + 0.013 in the fine
bran group (MPS ¼308 mm). This increased density
was not due to a change in fecal weight but was due to
a decrease in fecal volume, paralleling the effect dem-
onstrated for bran in Table 3. In ready-to-eat break-
fast cereals, the fiber MPS has been found to vary
between 350 and 2000 mm. Increased particle size
normalizes gastrointestinal transit time, delays gastric
tbl0003Table 3 Effect of grinding on dry bulk of wheat brans (ml g
1
):
duplicate measurements
Sieve aperture
a
Bran A
b
Bran B Bran C Bran D
As is 3.1 3.4 5.1 3.5
2.0 mm (10 mesh) 2.7 2.4 2.8 2.1
1.0 mm (18 mesh) 2.2 2.1 2.4 2.0
0.5 mm (35 mesh) 1.8 1.8 1.9 1.7
a
Using a MS Wiley mill. Mesh sieve equivalence is indicated in
parentheses.
b
Bran A was American Association of Cereal Chemists-certified soft white
wheat bran; brans B–D were soft or hard wheat brans from other sources.
1818 DIETARY FIBER/Properties and Sources

emptying, and may also affect the extent of insoluble
fiber fermentation. Although dietary fiber has many
positive effects on the digestive tract, excessive
milling of dietary fiber to produce small particles in
the range of 7–70 mm may have negative effects.
Smaller particles do not exhibit the transit time nor-
malization associated with larger bran particles, and
the persorption of a small amount of very fine fiber
particles may occur in some regions of the intestinal
wall. Contrary to persorbed starch granules, per-
sorbed fiber particles are not degraded in the blood
and may impose an increased workload on the
kidneys. Data on persorption are scarce, and much
work is needed on this subject.
0030 Studies with apples, apple pure
´
e, and apple juice
have highlighted the link between the physical integ-
rity of the cell wall and the beneficial effects of un-
processed dietary fiber, in this case, a lowering of the
glycemic index of a food (the rise in blood glucose
that accompanies ingestion and digestion of a carbo-
hydrate-containing food). Only the unprocessed apple
had a reduced glycemic index; the apple pure
´
e, con-
taining the physically degraded dietary fiber was simi-
lar to apple juice in response. This demonstrates the
importance of the physical encapsulation of available
macronutrients within the fiber matrix. This effect
has also been observed with rolled oats where higher
glycemic indices were observed with more finely cut
oats. Studies such as these show that the integrity of
the cell wall is important in bringing about beneficial
physiological effects, even in the presence of viscous,
soluble fiber types (b-glucans in the cell wall; see
section Bile Salt Binding and Serum Cholesterol
Reduction).
In vitro
Measurements
0031 In vitro properties of fiber isolates have been used
to estimate their potential in vivo effect. However, in
vitro data should be used with caution. The measure-
ments are influenced by the methods used to follow
digestion in vitro as well as by the methods used to
prepare the fiber. Other confounding influences are
also present: the fiber residue obtained in vitro may
have been altered by heat or chemicals or may still
contain residual digestible materials. For these
reasons, in vivo conditions generally result in a more
extensive digestion than that measured in vitro.
Digestive Events
0032 An important consideration when examining the
physiological effect of dietary fiber is the fact that
dietary fiber is concentrated and denuded only in
the terminal small intestine. This means that the diet-
ary fiber that is partially associated with digestible
materials in the jejunum and duodenum may have
different physical properties from those of an entirely
denuded fiber matrix at the terminal ileum. In the
large intestine, most of the soluble fiber and a large
but variable portion of insoluble dietary fiber are
degraded by the colonic microflora. As already dis-
cussed, fermentation plays an important role in medi-
ating many of the physiological effects associated
with dietary fiber.
Definition (Based on Function and
Structure)
0033In the early 1970s, it became evident that cellulose
represents only a small part of the total dietary fiber
content of foods. Prior to this, there was a tendency to
equate dietary fiber with cellulose because that is
what was measured using the old crude fiber method.
While this helped reinforce the use of the term dietary
fiber (cellulose has a fibrillar structure), cellulose rep-
resents only a small part of the total dietary fiber
content of foods. Thus, at that time, new methodolo-
gies were emerging, and it was clear that there was a
need for a dietary fiber definition that was linked to
the actual physiochemical composition of dietary
fiber.
0034It is in this context that the dietary fiber hypothesis
appeared, a few years after Cleave’s hypothesis in
1956, which related refined foods and Western dis-
eases. The dietary fiber hypothesis thus appeared as a
refocusing of Cleave’s original ideas, reversing the
original emphasis to become: a diet high in unrefined
edible plant material is protective against Western
diseases of the bowel. The dietary fiber hypothesis is
important in understanding the ‘dietary fiber con-
cept,’ since it links botanical structure to digestive
processes and health outcomes – mainly noninfective
colonic diseases such as constipation, diverticular dis-
ease, irritable colon, ulcerative colitis, appendicitis,
hemorrhoids, polyps, and cancer of the large bowel. It
is this link to these and other health outcomes that
has characterized much of the work in the dietary
fiber field over the last 30 years. The first dietary
fiber definition in 1972 sought to describe the plant
structures and components that were responsible for
mediating these health benefits: ‘‘that portion of food
which is derived from the cellular walls of plants and
is digested very poorly by human beings . . . Fibre is
composed largely of cellulose, . . . hemicelluloses, pen-
tosans, pectin and lignin.’’
0035There are many different definitions of dietary fiber.
However, they all include certain specific and critical
aspects. First and foremost, a definition must include
the fact that dietary fiber corresponds mostly to the
structural material in the edible part of plant foods
in the usual human diet. This has been consistently
DIETARY FIBER/Properties and Sources 1819

reflected throughout decades by the inclusion of the
terms ‘plant cell wall’ when referring to dietary fiber
(Table 4). The term ‘dietary fiber’ itself is something of
a misnomer, since dietary fiber is not fibrillar (except
for cellulose). To reflect this fact, many other terms
have been proposed over the years, including unavail-
able carbohydrate and plantix. None of these terms has
succeeded in replacing ‘dietary fiber.’
0036 Another important aspect of any dietary fiber def-
inition is that it be resistant to human digestive secre-
tions (including enzymes) and not absorbed in the
small intestine. The observation that dietary fiber
increased fecal weight initially gave the impression
that dietary fiber was resistant to both human digest-
ive secretions and to bacterial digestion in the lower
gut (fermentation). While it is true that dietary fiber is
not digested and absorbed in the small intestine, a
large proportion of dietary fiber is fermented in the
colon. The extent of this fermentation is a function of
several different factors (see Microbial Degradation
(Fermentability) Section), meaning that the extent of
fermentation cannot be use as a criterion to define
dietary fiber.
0037Using ‘resistance to endogenous digestive secre-
tions’ alone as a criterion may be confusing. For
example, some dietary fiber may be partly degraded
by acids in the stomach, while some small intestine-
digestible material can reach the colon where bacter-
ial fermentation occurs. In addition, a definition
based on the resistance to digestibility by endogenous
digestive secretions could be problematic if the mater-
ial is not specified as coming from the edible part of
plant foods. Indeed, such a definition could be erro-
neously interpreted as meaning that dietary fiber is
any material resistant to in vivo digestion or, even
as any material measured as dietary fiber by a specific
dietary fiber method. Among the materials to be ex-
cluded from the dietary fiber definition are ‘ruminant
fiber’ and the products formed during processing and/
or cooking. There have been requests to include some
other plant components as well as manufactured
polymers, and this is a subject of much debate.
Among the criteria that could be used to accept or
reject a material as dietary fiber, only ‘edible plant
cell wall’ is specific enough to allow an absolute
definition. In addition, it links current dietary fiber
tbl0004 Table 4 Use of the term ‘plant cell wall’ in defining dietary fiber over time
Year Text Reference
1885 The ‘cell membranes’ of Rubner Trowell HC (1975) In: Burkitt DP and Trowell HC (eds) Refined
carbohydrate foods and disease, p. 43. Academic Press, London.
1929 The ‘skeletal framework of the plant’ McCance RA and Lawrence RD (1929) The carbohydrate content
of foods. Medical Research Council special report series
No. 135, pp. 1–173. HM Stationary Office, London.
1981 The ‘remnants of plant cells’ AOAC Fiber Consensus (1981) Association of Official Analytical
Chemists 95th Annual Meeting, October 1981, Washington, DC.
1985 ‘Derived from plant cell walls and not digested by human
alimentary enzymes’
Trowell H, Burkitt DP and Heaton K (eds) (1985) Dietary fibre,
fibre-depleted foods and disease, Academic Press, London.
1990 The ‘structural plant cell wall composed of
polysaccharides and lignin’
Eastwood M (1990) Function of dietary fibre in the large
intestine. In: Southgate DAT, Waldron K, Johnson IT and
Fenwick GR (eds) Dietary Fibre: Chemical and Biological
Aspects, pp. 211–219. Cambridge: Royal Society of
Chemistry.
1995 ‘The ‘‘cell wall material of plant foods’’ responsible for the
physical form and texture of unprocessed plant foods’
Heaton KW (1995) The dietary fibre concept – time for a
re-evaluation? In: Sørensen A, Bach Knudsen KE, Englyst HN,
Gudmand-Høyer E and Nyman M (eds) Metabolic and
Physiological Aspects of Dietary Fibre in Foods: Recent Progress in
the Analysis of Dietary Fibre, COST 92, p. 15. Luxembourg:
Commission of the European Commission.
1995 The ‘plant cell walls’ are the common characteristic of
plant foods that constitute a high-fiber diet associated
with beneficial effects.
Cummings JH, Hudson GJ, Quigley ME and Englyst HN (1995)
The classification and measurement of dietary carbohydrates.
In: Sørensen A, Bach Knudsen KE, Englyst HN, Gudmand-
Høyer E and Nyman M (eds) Metabolic and physiological aspects
of dietary fibre in food. Recent progress in the analysis of dietary
fibre in food, COST 92, pp. 17–36. Luxembourg: European
Commission.
1999 ‘the cell walls of edible plant tissues in the traditional
human diet’
Mongeau R, Scott FW and Brassard R (1999) Definition and
analysis of dietary fiber. In: Cho SS, Prosky L and
Dreher M (eds) Complex Carbohydrates in Foods,
pp. 305–316. New York & Basel: Marcel Dekker.
1820 DIETARY FIBER/Properties and Sources

definitions to the original ‘dietary fiber concept,’
which recognized that a diet high in minimally pro-
cessed edible plant foods provided many health bene-
fits. Confusion often arises because of the overlapping
characteristics and physiological effects of the differ-
ent ‘candidate’ materials with those of a dietary fiber
‘reference.’ Examples of ‘candidates’ are polydex-
trose (a randomly bonded synthetic glucose polymer),
inulin and fructooligosaccharides (naturally occur-
ring glucose-fructose oligomers and polymers), and
resistant starch (formed by food-processing methods
and largely undigested in the small intestine). This
overlapping means that it becomes virtually impos-
sible to distinguish between fiber and nonfiber mater-
ial based on resistance to digestion alone. Debates
such as this started shortly after dietary fiber became
popular in the 1970s. They are still ongoing, as dem-
onstrated by the publication of a recent international
survey of 147 professionals. While a complete discus-
sion of the merits of either side is beyond the scope of
this chapter, it is important to remember the origins of
the dietary fiber definition and the dietary fiber hy-
pothesis that firmly linked fiber to physiological out-
come.
0038 The definition of dietary fiber was first centered on
the ‘original dietary fiber concept’ that linked the
consumption of ‘plant cell walls’ in unrefined, trad-
itional foods to positive physiological outcomes.
Table 5 shows two definitions that were proposed
after the turn of the millennium. These two defin-
itions have taken the original ideas of the dietary
fiber hypothesis into account as well as included a
reference to some specific physiological effects in
humans.
0047 While the newer definitions of dietary fiber specif-
ically mention the fact that dietary fiber has physio-
logical benefits, the list is rather specific and deals
primarily with effects observed with viscous fibers
(lowering post-prandial serum glucose and lowering
serum cholesterol) or effects brought about from
partial fermentation and water holding capacity
(laxation). These effects are well supported by clinical
experiments with laboratory animals and with
humans. However, the original dietary fiber concept
related dietary fiber intake to the diseases of Western
man, including heart disease, cancer and obesity. As
mentioned above, the link between viscous dietary
fiber and heart disease is supported in the literature
through its effect on serum cholesterol and triglycer-
ides but the link with other Western diseases has been
much harder to demonstrate. It has been well estab-
lished that eating patterns that favor foods high in
dietary fiber and low in red meat consumption signifi-
cantly reduce the risk of cancer, heart disease and
obesity. However, it has been relatively difficult to
isolate this effect to dietary fiber consumption per
se. Part of the problem comes from the almost exclu-
sive reliance on epidemiological studies to demon-
strate any association. There are several reasons
why epidemiological studies are generally favored.
First, these studies provide data on free-living humans
rather than in-bred laboratory animals. Second, these
studies follow the long-term effect of dietary fiber
consumption on health outcomes. Third, the sample
size can vary from a few hundred (case control stud-
ies) to many thousands of individuals (prospective
cohort studies). This gives the studies good statistical
power and lends weight to the conclusions. However,
epidemiological studies also suffer from a number of
shortcomings including: problems with assessing
food intake for the past 2–3 years (case control stud-
ies), or with monitoring food intake 2–3 times over a
period of 5 years and using this to predict disease
outcome 15 years later (prospective cohort studies),
problems that arise from the use of incomplete dietary
tables to calculate dietary fiber intake, and problems
arising from the use of inappropriate methods for
correcting for energy intake.
0048The most striking conclusion from epidemiological
studies is a failure to provide support for an inverse
correlation between dietary fiber intake and colon
cancer. This concept has long been entrenched in the
literature largely because of studies with laboratory
animals that have provided much data in favor of
tbl0005 Table 5 Recent proposed definitions of dietary fiber
Year Text Reference
2001 ‘ . . . the edible parts of plant or analogous carbohydrates that
are resistant to digestion and absorption in the human small
intestine . . . promotes beneficial physiological effects including
laxation, and/or blood cholesterol attenuation, and/or glucose
attenuation.’
American Association of Cereal Chemists (2001) Report of
the Dietary Fiber Definition Committee to the Board of
Directors of the American Association of Cereal
Chemists. 2001. The definition of Dietary Fibre. Cereal
Foods World 46: 112–129.
2002 ‘Dietary Fiber nondigestible carbohydrates and lignin that are
intrinsic and intact in plants. Functional Fiber consists of
isolated, nondigestible carbohydrates that have beneficial
physiological effects in humans.’
National Academy of Sciences (2002) Food and Nutrition
Board. 2002. Dietary Reference Intakes. Proposed Definition
of Dietary Fiber. National Academy Press, Washington DC.
DIETARY FIBER/Properties and Sources 1821

a protective effect of dietary fiber. The animal studies
are supported by plausible physiological and bio-
chemical mechanisms that can account for the ob-
served results. However, the epidemiological data is
inconsistent in its support for an inverse relationship
between colon cancer and dietary fiber intake and
clinical trials that examined the effect of diet on the
re-appearance of colon polyps (colon cell growths
that have a 1/20 chance of forming a cancerous
tumor) found no protective effect of dietary fiber.
Even though the link between adenomatous polyps
and cancer is not straightforward, these studies show
that more research is needed to elucidate the relation-
ship between the known physiological benefits of
dietary fiber and cancer.
Sources of Dietary Fiber
0039 The total dietary fiber content of a food depends on
many factors, including (1) the plant variety, (2) the
stage of maturity when harvested, (3) the plant
growing conditions, and (4) the method of food
preparation. Although not all the methodological
issues have been completely resolved with respect
to measuring dietary fiber, there is now general con-
sensus on the total dietary fiber content for a majority
of foods.
0040 The names of 49 common dietary fiber sources
are provided in Table 6. The foods are classified
according to their TDF content expressed on an ‘as
is’ basis. Since some methodological variability is
inevitable, the letters ‘E,’‘M,’ and ‘P’ in the right-
hand column designate which methods would give
TDF values in the range shown. Other methods not
included in the comparison are likely to agree with
these values. The letter ‘E’ refers to NSP as measured
by the Englyst GLC procedure and includes a lignin
value measured separately with potassium perman-
ganate. The letter ‘M’ refers to the Mongeau rapid
method, and the letter ‘P’ refers to the Prosky method.
The last two methods are gravimetric and include
lignin in the final determination. Table 6 shows a
considerable agreement between methodologies: 33
of 49 foods are designated ‘EMP,’ meaning that
all three methods give similar values. In addition,
three foods are designated ‘MP’ and 13 foods as ‘M’
only because sample size limitations precluded
measurement with other methods. The TDF values
are based on winter and summer collections of up
to 20 individual foods over 30 months in different
locations.
0041 Most fruits and vegetables contain low amounts of
dietary fiber (1.0–2.2% TDF) on an ‘as is’ basis.
Legumes contain intermediate amounts of dietary
fiber (around 4% TDF), and cereals contain between
1–2.2% TDF (corn kernels) and 15.5–15.8% TDF
(oat bran). The dietary fiber content of foods is
greatly influenced by the moisture content. Thus,
cereal TDF values are high because these foods con-
tain little moisture (up to 10% at most), whereas
fruits and vegetables contain 80–90% moisture on
an ‘as is’ basis. The values of Table 6 are reported as
a percentage of weight, but when calculating dietary
fiber intake, it is important to factor in the serving
size (30 g for breakfast cereals, 50 g for lettuce and
raisins, and 125 g for vegetables and fruits). For
example, 1 serving of Bran flakes will provide ap-
proximately 4 g of dietary fiber, 1 serving of lettuce
will provide 0.35 g of TDF, 1 serving of raisins will
provide about 1.8 g TDF, and 1 serving of corn
will provide about 2 g of TDF.
tbl0006Table 6 Sources of dietary fiber
TDF
a
Foods Methods
b
0.1–0.5 White rice, cooked M
Orange juice from concentrate M
0.6–0.9 Lettuce M
Cucumbers EMP
Pineapple, melons (cantaloupe) EMP
1.0–2.2 White bead EMP
Corn kernel (canned), potatoes (boiled),
green beans (boiled), cabbage (raw),
celery (raw), onion (cooked),
green peppers (raw)
EMP
Cauliflower (cooked), tomatoes M
Apple, bananas, orange, pears,
strawberries, cherries, grapefruit,
plums
EMP
Apple pie, blueberry pie, pasta (cooked) M
2.3–3.0 Rye bread M
Mushroom (E ¼2.0) MP
Beets (canned), rutabaga (raw),
carrots (raw), broccoli (raw)
EMP
Blueberries (E ¼1.9) MP
3.1–4.3 Brown rice EMP
Raisins (seedless) M
4.4–6.4 Bran muffins, wheat cereal,
wholemeal bread
EMP
White beans (baked) EMP
Kidney beans (baked), peanut butter M
6.5–7.3 Peanuts EMP
Green peas (boiled) M
8.0–9.3 Oatmeal cereal EMP
9.4–10.3 Shredded wheat EMP
12.3–13.1 Bran flakes EMP
15.5–15.8 Oat bran EMP
40.6–40.9 AACC hard wheat bran (E ¼36.4) MP
a
TDF, total dietary fiber range in grams per 100 g on an ‘as is’ basis.
b
Methods: E, Englyst GLC method plus potassium permanganate lignin;
M, Mongeau rapid gravimetric method; P, Prosky gravimetric method.
From Mongeau R and Brassard R (1989) A comparison of three methods
for analyzing dietary fiber in 38 foods. Journal of Food Composition and
Analysis 2: 189–199; Mongeau R, Brassard R and Verdier P (1989)
Measurement of dietary fiber in a total diet study. Journal of Food
Composition and Analysis 2: 317–326.
1822 DIETARY FIBER/Properties and Sources

Glossary
.0042 Digestive secretions: enzymes and other secretions
produced by the digestive organs (e.g., salivary
glands and exocrine pancreas, stomach and small
intestinal mucosa, and liver); this excludes enzymes
produced by intestinal bacteria.
.
0043 Digestibility: degradation of ingested food com-
ponents by endogenous digestive secretions in the
upper gastrointestinal tract.
.
0044 Fermentability: degradation of undigested food
components (e.g., dietary fiber) by the intestinal
microflora, mostly in the large intestine.
.
0045 Pectic substances: mixtures of acidic and neutral
polysaccharides that can be extracted with water
from plant tissues and are rich in galacturonic acid;
together with hemicelluloses, they form the cell
wall matrix in which are enmeshed cellulose fibers.
.
0046 Persorption: the paracellular passage of small
amounts of solid food components from the lumen
of the intestine into the lymphatic and blood circula-
tion, this being without structural changes.
See also: Bile; Carbohydrates: Classification and
Properties; Cellulose; Dietary Fiber: Determination;
Physiological Effects; Effects of Fiber on Absorption;
Bran; Energy Value
Further Reading
Anderson JW, Allgood LD, Lawrence A et al. (2000) Chol-
esterol-lowering effects of psyllium intake adjunctive to
diet therapy in men and women with hypercholesterol-
emia: meta-analysis of 8 controlled trials. American
Journal of Clinical Nutrition 71: 472–479.
Baintner K (ed.) (1986) Intestinal Absorption of Macromol-
ecules and Immune Transmission from Mother to
Young. Boca Raton, FL: CRC Press.
Bolton RP, Heaton KW and Burroughs LF (1981) The role
of dietary fibre in satiety, glucose, and insulin: studies
with fruit and fruit juice. American Journal of Clinical
Nutrition 34: 211–217.
Brown L, Rosner B, Willett W and Sacks FM (1999) Chol-
esterol-lowering effects of dietary fiber: a meta-analysis.
American Journal of Clinical Nutrition 69: 30–42.
Carpita NC (1990) The chemical structure of the cell walls
of higher plants. In: Krichevsky D, Bonfield C and
Anderson JW (eds) Dietary Fiber: Chemistry, Physi-
ology, and Health Effects, pp. 15–30. New York:
Plenum Press.
Cummings JH (1986) The effect of dietary fiber on fecal
weight and composition. In: Spiller (ed.) Handbook of
Dietary Fiber in Human Nutrition, pp. 211–280. Boca
Raton, FL: CRC Press.
Dreher ML (ed.) (1987) Handbook of Dietary Fiber: An
Applied Approach. New York: Marcel Dekker.
Eastwood M (1990) Function of dietary fibre in the large
intestine. In: Southgate DAT, Waldron K, Johnson IT and
Fenwick GR (eds) Dietary Fibre: Chemical and Bio-
logical Aspects, pp. 211–219. Cambridge: Royal Society
of Chemistry.
Heaton KW (1995) The dietary fibre concept – time for a
re-evaluation? In: Recent Progress in the Analysis of
Dietary Fibre, Metabolic and Physiological Aspects of
Dietary Fibre in Foods, COST 92, p. 15. Luxembourg:
Commission of the European Communities.
Lee SC and Prosky L (1995) International survey on dietary
fiber: definition, analysis, and reference materials. Jour-
nal of AOAC International 78: 22–36.
Mongeau R and Brassard R (1982) Insoluble dietary fiber
from breakfast cereals and brans: bile salt binding and
water-holding capacity in relation to particle size. Cereal
Chemistry 59: 413–417.
Mongeau R, Scott FW and Brassard R (1999) Definition
and analysis of dietary fiber. In: Cho SS, Prosky L
and Dreher M (eds) Complex Carbohydrates in Foods,
pp. 305–316. New York & Basel: Marcel Dekker.
Selvendran RR and Robertson JA (1990) The chemistry of
dietary fibre: an holistic view of the cell wall matrix. In:
Southgate DAT, Waldron K, Johnson IT and Fenwick
GA (eds) Dietary Fibre: Chemical and Biological
Aspects, pp. 27–43. Cambridge: Royal Society of
Chemistry.
Spiller GA (1986) Definition of dietary fiber. In: Spiller GA
(ed.) Handbook of Dietary Fiber in Human Nutrition,
pp. 15–18. Boca Raton, FL: CRC Press.
Trowell H (1972) Ischemic heart disease and dietary fiber.
American Journal of Clinical Nutrition 25: 926–932.
Wolk A, Manson JE, Stampfer MJ et al. (1999) Long-term
intake of dietary fibre and decreased risk of coronary
heart disease among women. Journal of the American
Medical Association 281: 1988–2004.
Determination
R Mongeau, Ottawa, Aylmer (Que
`
bec), Canada
S P J Brooks, Ottawa, Canada
Copyright 2003, Elsevier Science Ltd. All Rights Reserved.
Introduction
0001This chapter presents a brief overview and compari-
son of commonly utilized methods for quantifying
dietary fiber in food. There are, fundamentally, only
two types of methods. Both rely on the removal of
nonfiber components by a combination of physical,
chemical, and enzymatic procedures prior to meas-
urement of dietary fiber. The gravimetric methods
measure dietary fiber by weighing the residue and
the gas-liquid chromatographic (GLC) methods
measure dietary fiber by hydrolyzing a treated sample
with acid to sugar monomers prior to quantification
DIETARY FIBER/Determination 1823

by GLC. Although, on the surface, both types of
methods appear similar, there are fundamental prac-
tical differences that have come about because of
variations in the individual method steps and differ-
ences in the choice of reagents and enzymes. These
practical differences have significant consequences
since they determine the final composition of dietary
fiber as measured by each method.
0002 Procedures have been described for isolating and
analyzing dietary fiber in specific foods as well as
analyzing carbohydrate polymers that are not
digested in the small intestine and are not identified
as dietary fiber by the current Association of Official
Analytical Chemists (AOAC) methods. The present
article, however, will focus on practical methods that
are applicable to the routine analysis of a wide range
of foods. Most total dietary fiber (TDF) methods also
allow for the quantification of different fiber frac-
tions such as soluble fiber, insoluble fiber, hemicellu-
loses, cellulose, lignin, etc. In this article, only the
methods that measure TDF are reviewed. This is
partly because of space limitations and partly because
the individual fiber components have not yet been
positively associated with human physiological
effects. For example, soluble and insoluble fiber can
be readily measured but these designations have little
physiological meaning since the digestive properties
and physiological benefits associated with some sol-
uble fiber can be found in some insoluble fibers and
the physiological benefits associated with insoluble
fibers are also found with some soluble fibers. In
addition, some processing techniques can also partly
depolymerize insoluble fiber that may then be found
in the soluble fiber fraction.
0003 The early work of Van Soest focused on measuring
ruminant fiber (mainly hemicellulose, cellulose, and
lignin) but these procedures could not be used to
measure TDF because they would have underesti-
mated the TDF content of human foods by missing a
significant proportion of the soluble dietary fiber. All
the modern methods incorporate steps to measure
soluble fiber. This is important since it can represent
a significant proportion of the TDF of a food. The
chemical complexity and physical variability of the
dietary fiber fraction in various foodstuffs as well as
the various food-processing techniques have con-
tributed to the difficulties encountered in devising
adequate dietary fiber methods. Many of the present-
day TDF procedures are based on the pioneering
work of Southgate.
0004 Ideal reference standards do not exist for dietary
fiber analysis. Purified fiber components and unpuri-
fied standardized ‘reference samples’ are both used as
standards of analysis, but the former group rarely
represents the complex dietary fiber matrix found in
foods and the latter group remains variable in com-
position, structure, and properties.
Historical Perspectives on Dietary Fiber
Values
0005Several different measures of dietary fiber and dietary
fiber fractions have been reported for foods over the
last 150 years. It is important to put these values into
some sort of context to understand the differences
between these measurements and the implications
for human dietary fiber research.
0006The crude fiber method was standardized in the
late 19th century in Germany. It was initially de-
veloped to analyze ruminant feeds and silage. It is a
highly empirical method that uses sequential extrac-
tions with hot dilute acid and alkali. The crude fiber
residue contains no soluble fiber and retains only
about 15% of the total hemicelluloses, 50–100%
cellulose, and 10–50% lignin. The variability in the
method and the variable losses of various dietary fiber
fractions mean that crude fiber values are of little
value in human nutrition. (See Cellulose; Hemicellu-
loses; Lignin.)
0007Acid detergent fiber (ADF) represented the first
method that reproducibly maintained specific dietary
fiber fractions. ADF is obtained by refluxing in
1 N mol l
1
sulfuric acid containing cetylmethylam-
monium bromide detergent followed by filtering and
washing. This fraction contains mostly cellulose and
lignin.
0008Neutral detergent fiber (NDF) represents the insol-
uble fraction of TDF. It is obtained by refluxing in a
solution of sodium lauryl sulfate followed by filtra-
tion and washing. NDF includes most of the cell wall
material but excludes the soluble b-glucans and sol-
uble hemicelluloses. It includes the ADF fraction as
well as the insoluble hemicelluloses. It thus under-
estimates TDF because it does not measure soluble
dietary fiber.
0009In some older tables, TDF values are reported as
the sum of NDF and pectin (estimated TDF) but, as
shown in Figure 1, this value is not always accurate.
Figure 1 also presents the relationship between insol-
uble dietary fiber (essentially NDF but using porcine
pancreatic a-amylase to remove starch; see below)
and TDF (as determined by the rapid HPB (Health
Protection Branch) method; see below) determined
for the same foods. The insoluble dietary fiber under-
estimates the TDF value by approximately 40% but
this is not constant. The differences between TDF and
insoluble dietary fiber represent actual differences
measured on the same food sample but the differences
between the estimated TDF and the actual TDF
values were performed at different times with
1824 DIETARY FIBER/Determination
different procedures and using different food samples
so that procedural variations may account for some
of the differences. For example, it is known that some
previously determined NDF values were measured
after incubation at lower (suboptimal) temperatures
that may have given higher and more variable values
due to incomplete starch digestion.
0010 In addition to the relationships described above,
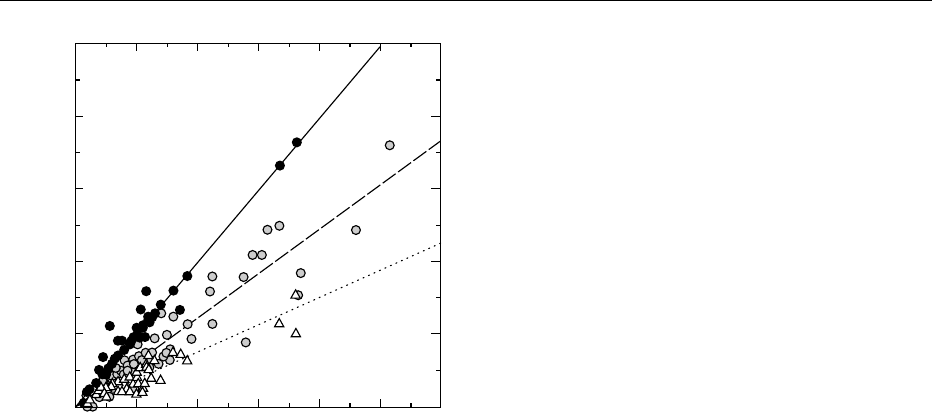
Figure 1 shows the relationship between crude fiber
(CRF) and TDF (as determined by the rapid HPB
method) for foods from the Canadian Nutrient File.
Like the insoluble dietary fiber values, the CRF values
significantly underestimate the TDF values; however,
in this case, the variability is largely due to the CRF
method. Figure 1 shows that no consistent relation-
ship between TDF and CRF or insoluble dietary fiber
(and thus NDF). Thus, it is not possible to predict
TDF values from either CRF or insoluble dietary
fiber: a complete TDF measure is required.
Gravimetric Methods
0011 Gravimetric methods use enzyme and/or chemical
treatments to remove material that is digestible in
the small intestine. Ideally, these treatments should
mimic the digestive processes to arrive at a sample
that is representative of the material entering the large
intestine of humans. Because dietary fiber represents
the structural components of plant cell wall material
(See Dietary Fiber: Properties and Sources), the
weight of the residue is corrected for the presence of
noncombustible materials (such as minerals) and cor-
rections may also be applied for components that are
incompletely digested by the procedure (such as pro-
tein). The remaining weight is considered to be diet-
ary fiber. In order to measure the remaining material
gravimetrically, it must be filtered through glass cru-
cibles of defined porosities. It is, therefore, important
when preparing the sample to insure that it is not
too finely ground. The use of a Wiley mill avoids
these problems and minimizes particle loss during
filtration. Many different gravimetric methods have
been published with many variations in reagents,
temperatures and incubation times. However, rela-
tively few of these methods have been approved by
the Association of Official Analytical Chemists
(AOAC) after rigorous testing and interlaboratory
verification. Three AOAC approved methods will be
discussed in detail in this section: the rapid Health
Protection Branch method (rapid HPB method), the
Prosky method, and the Lee method. Before present-
ing these methods, we will first discuss the detergent
system.
The Detergent System
0012When introduced in 1963, the detergent system rep-
resented a great improvement over the CRF method
and it is still in use today for measuring insoluble
dietary fiber. It has been widely used to measure silage
and ruminant feed fiber. The system includes the NDF
and the ADF procedures, which use hot detergent to
remove digestible material and soluble fiber. The de-
tergent system discriminates among the insoluble
fibers – hemicelluloses, cellulose, and lignin. A che-
lating agent in the neutral detergent solution permits
the solubilization of insoluble pectin. When NDF and
ADF procedures are performed sequentially, the dif-
ference in weight between the NDF and ADF extracts
is an estimation of the insoluble hemicellulose frac-
tion. The ADF residue contains lignin, cellulose, and
(if present) cutin. Lignin can be measured by gently
oxidizing the ADF residue with potassium perman-
ganate. A 72% sulfuric acid treatment hydrolyzes
cellulose leaving cutin, phlobaphenes, and Maillard
products.
NDF and Porcine a-Amylase
0013Since water-insoluble dietary fibers represent a major
fraction of total dietary fiber, the NDF procedure
represents a simple method that can form part of an
10
8
6
4
2
0
024681012
TDF (rapid HPB method)
(g g
−1
'as is')
Estimated TDF, IDF, or CRF
(g g
−1
'as is')
fig0001 Figure 1 Relationship between crude fiber (CRF; white tri-
angles), insoluble fiber (IDF; gray circles), estimated total dietary
fiber (TDF: total of pectin þ neutral detergent fiber (NDF), black
circles), and TDF values for different types of foods. The TDF and
CRF values are from the Canadian Nutrient File (Health Canada
1997) and Mongeau et al. (1989). TDF and IDF values were deter-
mined by the rapid HPB method (AOAC 992.16, see text). The solid
line represents a 1:1 correspondence. The dashed line (IDF) and
the dotted line (CRF) are regression lines with the intercept fixed at
the origin. Insoluble dietary fiber is essentially NDF but includes a
treatment with porcine pancreatic a-amylase. The NDF þ pectin
values are ultimately from the USDA Nutrient Database (1999).
DIETARY FIBER/Determination 1825