Алексеев В.В. Физическое и математическое моделирование экосистем
Подождите немного. Документ загружается.


Тогда G(p) преобразуется к виду
к* (
п
\ 8
и
УР) — 1 — 6*ехр(р + (х)а '
и
характеристическое уравнение приобретает форму
1-6*ехр[
—
а]
=
0.
(6.46)
Произведение Sg определяет взаимное влияние консумента и расте-
ний;
Sg= (влияние консумента на ресурс) X (влияние ресурса на
рождаемость консумента). Урав-
нение
(6.46)
имеет бесконечное Im.p
число корней (собственных зна-
чений)
для каждого значения Sg.
д
-
При
изменении Sg от нуля до
бесконечности,
каждый корень
описывает траекторию на комп-
лексной
плоскости. Локализация
Рис.
6.11. Общее расположение
корней
характеристического урав-
нения
(6.46) на комплексной пло-
скости.
корней
может служить хорошей иллюстрацией динамики линеари-
зованной
системы при разных режимах внешних условий.
Для случая Bg = О, уравнение
(6.46)
редуцируется к виду
) = 0. (6.47)
Уравнение
(6.47)
имеет следующие корни.
(6.48)
Уравнение
(6.48)
имеет один действительный корень р —lnft*/a —
—
ц = р и комплексные корни, определяемые из условия р = р ±
±
/со, откуда со = 2яп/а (п = 1, 2...).
Общее расположение корней показано на рис. 6.11 для случая
b(a) =
b*6{a
— а). Ветви начинаются при Bg — 0{l) и проведены
до Sg = + оо (2). Корни уравнения
(6.46)
при каждом фиксиро-
ванном
значении Sg находятся на ветвях, как отмечено, например,
точками Pi (i= I, 2, ...). Зная распределение корней для уравне-
ния
(6.46), можно получить частотные отклики на периодическое
воздействие u(t) для ресурса #(ко)/е7(ио) и общей плотности по-
221

пуляции Y(i(u)/U(m). Поскольку первая пара комплексно сопря-
женных корней лежит очень близко к реальной оси, результирую-
щая
резонансная частота может быть бесконечно малой. Из
рис.
6.11 видно, что
существует
область значений параметров,,
в
которой линеаризованная модель переходит от устойчивости
к
неустойчивости, т. е. при изменении параметра взаимодействия
Eg в определенной области, первая пара собственных значений
пересекает мнимую ось. В этой точке в нелинейной системе наблю-
даются колебания с малой амплитудой, которая в дальнейшем
растет при увеличении Eg.
Если
окно
репродуктивное™
у консумента имеет конечную
1
ширину
7. то уравнение
(6.46)
должно быть заменено на уравне-
ние
(6.44), и характеристическое уравнение приобретает вид
Корни
р = — А; р = — ц остаются на месте, как и в предыдущем
случае, исследование комплексных корней оказывается более
сложным, однако качественное поведение системы остается
прежним.
6.5.
Трофические системы
с
возрастным
распределением
Объединяя несколько популяций с возрастным распределением,,
можно сформировать систему с трофическими взаимодействиями.
Так,
простейшая система «хищник—жертва» (рис. 6.12) может
Хищник
Жертва
Рис. 6.12.
Система
«хищник—жертва»
при на-
личии
возрастной
структуры
популяций.
быть получена объединением
двух
уравнений типа (6.3), при этом
в
уравнении для жертвы коэффициент смертности заменяется на
фактор
смертности, зависящий от общей численности
другого
222

вида [117, 119, 272]:
«р + Ур
\
р
р
p(0,
t)= \
m
p
(a',
t,
H{t-x))p(a', t)da',
(6-49)
A(0,
t)= j
m
h
(a', t)h(a
r
, t)da',
a
h
CO
OO
H(t)=\h(a,
t)da, P(t)=\p(a, t)da,
где
h(a,
t)—плотность возрастного распределения популяции
жертвы; р(а, t)—плотность хищника.
В простейшем случае, когда рождаемость как
у
хищника, так
и
у
жертвы приходится на строго определенный возраст
т
р
=
т
р
Ь{а
—
а
р
),
(6.50)
m
h
=
m
h
b(a
— a
h
),
анализ,
аналогичный проведенному
в
предыдущем параграфе, дает
для отклонения общей численности хищника
в
частотном виде вы-
ражение
U tM')
+
jjgr(«). (6-51)
pw
s +
Цр
1— exp (— a
p
s)
pv
'
T
\i
p
:
где Up(s)—вклад окружающей среды
в
рождаемость хищника.
Для популяции жертвы имеем
Y
h
(s)=
}. -. \ г
U
h
(s)
-
4*2- Y
p
(s). (6.52)
s
+
M-A 1— exp
(—a^s)
"
v
' n^s
pv
'
x
'
Разрешая
алгебраические уравнения (6.50)
и
(6.51)
относительно
Y
p
(s)
и
Vft(s),
получим
1
1
s
2
I s +
jip
I
— exp (—sap)
fiftS
s +
t*p 1 — exp
(—sa
p
)
|ЛрА
1 1
ix
p
s
S
+
ЦЙ
I— exp
(—
11
I—
exp(—sa
h
)
где
(6.53)
223
Из
(6.53)
нетрудно видеть, что кроме резонансных членов
(1—ехр(—sap)) и (1 — ехр(—sa
ft
)), которые наблюдались в изо-
лированных популяциях, общий отклик системы «хищник-жертва»
характеризуется гармонической составляющей вида (s
2
+ Q
2
) с ча-
стотой й.
В динамике каждой популяции появляются, как минимум, две
синусоиды, приводящие к новому явлению — гармоническим «бие-
ниям» с частотой, пропорциональной разности
между
частотой
внешнего воздействия и резонансными частотами. Период биений
для частот fii и Q
2
равен
2n/(Qi — Q
2
). Таким обра-
зом,
явление «биений» ока-
зывается новым динамиче-
ским
свойством, присущим
связанным
системам.
Система
«паразит—хо-
зяин».
Практические задачи
борьбы с вредными насеко-
мыми
обусловили повышен-
ное
внимание к эксперимен-
тальному и теоретическому
изучению динамики попу-
ляционных
систем «пара-
зит—хозяин» (насекомое и
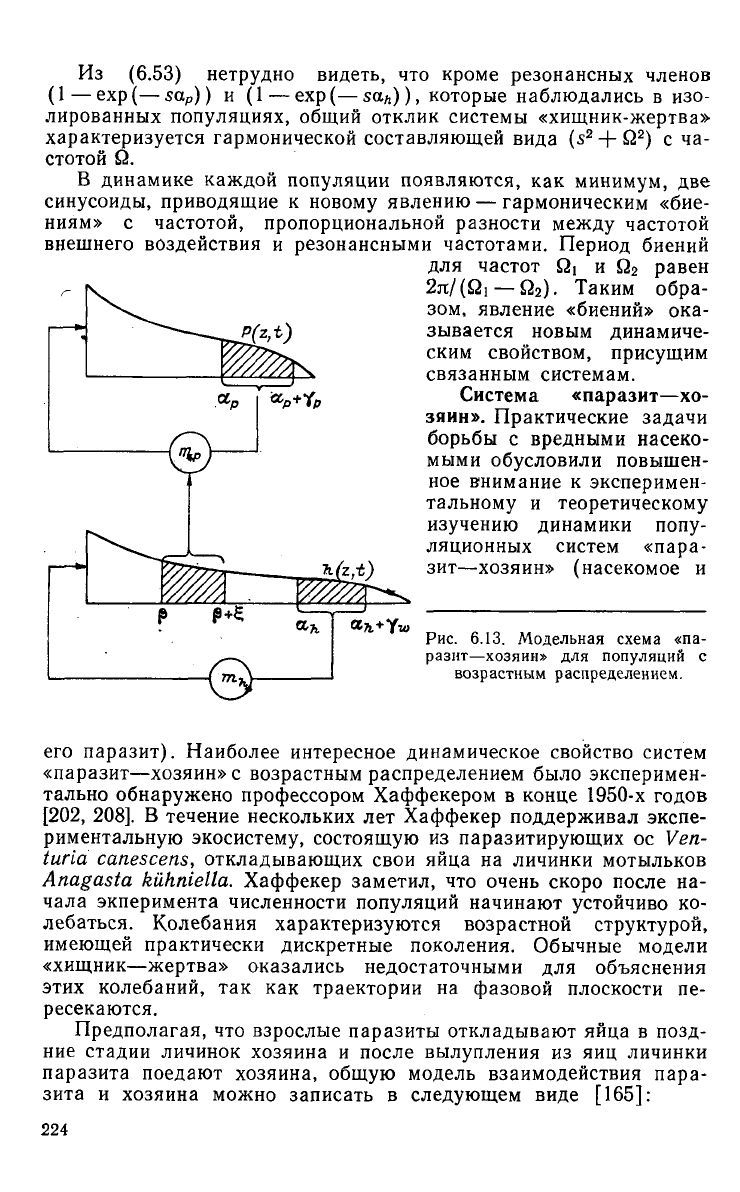
Рис.
6.13. Модельная схема «па-
разит—хозяин»
для популяций с
возрастным
распределением.
его паразит). Наиболее интересное динамическое свойство систем
«паразит—хозяин» с возрастным распределением было эксперимен-
тально обнаружено профессором Хаффекером в конце
1950-х
годов
[202, 208]. В течение нескольких лет Хаффекер поддерживал экспе-
риментальную экосистему, состоящую из паразитирующих ос Ven-
turia
canescens,
откладывающих свои яйца на личинки мотыльков
Anagasta
kiihniella.
Хаффекер заметил, что очень скоро после на-
чала экперимента численности популяций начинают устойчиво ко-
лебаться. Колебания характеризуются возрастной структурой,
имеющей практически дискретные поколения. Обычные модели
«хищник—жертва» оказались недостаточными для объяснения
этих колебаний, так как траектории на фазовой плоскости пе-
ресекаются.
Предполагая, что взрослые паразиты откладывают яйца в позд-
ние
стадии личинок хозяина и после вылупления из яиц личинки
паразита поедают хозяина, общую модель взаимодействия пара-
зита и хозяина можно записать в следующем виде [165]:
224

1) для паразита
(6.54)
р{0, /) = ] т
р
\а, t, H
0
)p(a, t)da,
2) для хозяина
dh , dh
=
—йл^а, *, Я, Я
о
, Ро)Л,
А
(0, t)= \ m
h
(a, t, H
0
{t—x))hda,
(6.55)
p
+ 6
p
+ Vp
=
J
p(a,t)da,P(t)
=
где p(a, t); h(a, t) — плотности популяций паразита и хозяина;
P(t),
H(t) — общие количества особей паразитов и хозяев; #о(О —
число личинок хозяина, подверженных возможности заражения;
Po(t)— число взрослых особей паразита.
Модельная схема системы «паразит—хозяин» представлена на
рис.
6.13.
Центральное биологическое свойство модели — специфика взаи-
модействия
двух
популяций. Простейший вид взаимодействия мо-
жет быть получен из предположения о случайном поиске каждым
паразитом личинок хозяина, считая площадь «обследования» S
o
для одного паразита либо постоянной, либо зависящей от возра-
ста встречающихся личинок хозяина
S
0
(a).
Если личинки хозяина
распределены по плоскости равномерно, то вероятность поражения
паразитом является пуассоновским процессом. Взаимодействие
между
паразитом и хозяином принимает вид
[
Число
зараженных личинок ]
\--=Ыг{а)[\
— ехр(—S
0
(a)P
e
)].
хозяина возраста а \
у
'
l v v v
'
п
(6.56)
Зараженный
хозяин считается мертвым и заменяется яйцами в по-
пуляции
паразита. Смертность хозяина имеет также некоторую
зависимость от плотности. Плодовитость хозяина включает запа-
здывающий эффект, который определяется его питанием в про-
шлом.
15
Заказ
№ 57 225

Теоретический анализ показывает, что в системе «паразит—
хозяин», так же, как и в других системах с возраст-специфиче-
скими
взаимодействиями, можно наблюдать резонансные явления.
Рассмотрим линеаризованный вариант уравнений (6.55). Пусть
Х
р
(а, s) и Xh(a, s)—преобразования Лапласа для плотностей па-
разита и хозяина р(а, t), h(a, t). Определим преобразования для
взаимодействующих популяционных сегментов как
р
+ б
H
0
(s)= \ X
h
da,
Р
H(s)= \ X
h
da,
a
h
«p + Vp
P(s)= 5 X
p
da,
pda, (6.57)
Y
h
(s)=\x
h
da.
о
Преобразованная
по Лапласу система (6.55) приобретает вид
da
X
p
(0, s) = i
(6.58)
X
h
(0, s) = C
3
#,
где С
и
С
2
, С
3
— константы линеаризации; y(a) = v в «окне» воз-
растов репродуктивности (р, Р + б) и равно нулю вне этого ин-
тервала.
Решая
уравнения (6.58) для паразита, после интегрирования
получим
Р (s) = G
l
(s)H
0
(s),
(6.59)
Y
p
(s) = G
2
(s)H
0
(s).
226

Решая
уравнения
(6.58)
для хозяина и проинтегрировав по под-
ходящим интервалам,получим
=
G
5
(s)P(s),
(6.60)
где Gi(s) (г = 1, ... 7)— функции переноса.
Каждая из функций переноса Gi(s) (i= I, ..., 7) представляет
собой комплексную функцию оператора запаздывания
ехр[—(s
+
+
ц)а]. Для хозяина паразит Яо фактически является периодиче-
ским
внешним воздействием, его роль состоит в возбуждении по-
пуляционных волн, которые
дают
сложный резонансный спектр.
Фазовый
портрет общих численностей популяций имеет в общем
случае
вид замкнутой кривой с несколькими пересечениями. Кроме
простых резонансов, в системе возможны биения с периодом
больше резонансного периода. Модель
(6.54)
в целом хорошо соот-
ветствует
экспериментальным данным для систем «паразит—
хозяин». Так, Мэй выполнил сравнение расчетов по модели
(6.54)
с функцией поиска в форме
(6.56)
с экспериментальными дан-
ными
Барнетта по взаимодействию
между
оранжерейной белой
мушкой
Trialeurodes
vaporariorium
и ее паразитом
Encarsia
for-
mosa
[165]. Рисунок 6.14 показывает, что хорошее совпадение
экспериментальных и модельных значений наблюдалось в течение
22 генераций насекомых. Динамика системы представляла собой
усиливающиеся колебания, приводящие в конце концов к разру-
шению
системы.
Тщательное исследование проводилось для математической
модели, описывающей эксперименты Хаффекера (оса
Venturia
canescens
и ее паразит
Anagasta
kuhniella) [202, 326]. Один из
вариантов эксперимента и его модельного описания показан на
рис.
6.15. Как видно из рисунка, модель хорошо соответствует
опытным
данным. Динамические режимы модели исследовались
при
варьировании параметров v и Ь, отвечающих за поисковые воз-
можности паразита. В соответствии с экспериментальной процеду-
рой,
в качестве начального условия рассматривалась популяция,
состоящая только из взрослых самок. В отсутствие паразита по-
пуляция
хозяина постепенно эволюционировала к стабильному не-
прерывному распределению, как показано на рис. 6.16. Этот же
режим в ряде случаев наблюдается и при наличии паразита,
режим получил название «непрерывной распределенной моды ге-
нерации»— CGM (continuous generation mode). Более интерес-
ным
оказалось
другое
стабильное состояние, названное «дискрет-
ной
модой генерации» — DGM (discrete generation mode), кото-
рое и наблюдалось Хаффекером экспериментально. Если обе по-
пуляции
стартуют
с некоторого числа особей, имеющих ограничен-
ный
диапазон возрастов, то в последующем возрастной профиль
15* 227

20
Рис.
6.14. Сопоставление расчетов по мо-
дели (6.54),
(6.56)
с эксперименталь-
ными
данными по взаимодействию оран-
жерейной мушки
Trialeurodes
vaporario-
т'шт (Г) с ее паразитом
Encarsia
for-
mosa
(II) [165].
/ —
экспериментальные данные,
2 —
расчет
по
модели.
10 15
Генерации
Рис.
6.15. Динамика эксперименталь-
ной
системы (/) «паразит — хозяин»
(оса
Venturia
canescens
(а) и ее па-
разит
Anagasta
kuhniella
(б)) и ре-
зультаты
расчетов (2) [202].
Рис.
6.16. Эволюция популяции хо-
зяина
к стабильному возрастному
распределению в отсутствие паразита.
Первоначальные волны индуцирова-
ны
периодическим добавлением зре-
лых самок [326].
Возраст
Рис.
6.17. Стробоскопическое изобра-
жение с периодом 51 сутки (время
генерации хозяина) возрастного про-
филя
популяции хозяина.
Первоначальный импульс переходит
в ста-
бильное периодическое распределение [326].
Режим «дискретная мода генерации».

поддерживает эту конфигурацию (рис.
6.17).
Импульс проходит
через генерацию, так что периодический осмотр каждой генерации
дает
стабильное конечное возрастное распределение. Паразит,
имеющий
время генерации примерно в два раза меньше, чем хо-
зяин,
дает
также стабильное рас-
пределение типа DGM, состоящее
из
движущихся в противополож-
ных направлениях волновых про-
филей.
Таким образом,
хотя
вся
система имеет период 51 день,
каждый 25-й день возрастной про-
филь
паразитов конденсируется
в
бимодальный движущийся вол- '
новой
профиль, как показано на
Рис.
6.18. Стробоскопическое изображе-
ние
с периодом 25 сут (время генерации
паразита) возрастного профиля генера-
ции
паразита.
Первоначальный импульс переходит
в ста-
бильное периодическое распределение. Режим
Режим «дискретная мода генерации:».
Возраст
рис.
6.18. Такая бимодальная
структура
наблюдалась эксперимен-
тально Хаффекером и была им названа термином «когенерация».
Результирующий предельный цикл на фазовой плоскости
10
th
Н
Популяция
хозяина
Рис.
6.19. Динамика общих численностей паразита и хозяина (а) и
изображение процесса на фазовой плоскости (б) [202, 326].
(рис.
6.19) имеет одно пересечение из-за присутствия
двух
волн
паразита на одну генерацию хозяина. Таким образом, в фазовом
пространстве
существуют
два состояния — одно стационарное, дру-
гое в виде движущихся волн. Как показано в работе [165], эти
два состояния
обладают
противоположной чувствительностью
к
изменениям параметров.
229

Чувствительность решений к малым вариациям параметров за-
дается коэффициентами чувствительности, определенными как
[280, 318]
a*(/
>z
) =
-^-(/,
г, я),
(6.61).
где
я —
любой исследуемый параметр (например,
v или Ь).
Набор
уравнений
для
эволюции коэффициентов чувствительно-
сти можно получить, дифференцируя уравнения модели
(6.55)
no-
параметрам
я.
Полученные уравнения имеют
вид
~dt
«o + V
o
p
n(0,t)= j m
p
o
p
n
da+ J
-^-pda,
(6.62)-
К , К
Л ^ аа
•о*
(0,/)=
J
Чувствительность популяций к изменению параметров можно полу-
чить, интегрируя по всем возрастам и затем дифференцируя по-
параметрам:
<*л
(t) = §(т
р
—
\1
Р
)
a
p
n
da — \ —^- р da,
(6.63)
оо
оо
а* (0= J im
h
-li
h
)aUa+\-^-h
da.
о
о
В целом уравнения чувствительности имеют ту же форму, что и
линеаризованные
уравнения для анализа устойчивости, но с реше-
нием
первоначальной модели в качестве вынуждающей силы.
Исследование чувствительности модели к вариациям параметра
поиска
v у паразита
дает
достаточно неожиданные результаты —
в
режиме дискретных мод с увеличением стационарной плотности
хозяина плотность паразида уменьшается, в режиме непрерывных
мод — наоборот. Таким образом, коэффициенты чувствительности
оь и оь меняют знак при переходе от одного режима к
другому.
Экспериментальное подтверждение изменения знака чувствитель-
ности
при смене динамических режимов дано в работе [326].
230