Страйер Л. Биохимия. Том 3
Подождите немного. Документ загружается.

Рабдовирусы -
вирусы, имеющие форму пули. К ним относятся вирусы везикулярного стома-
тита и бешенства. Название происходит от греческого слова rhabdo, что озна-
чает «палочка».
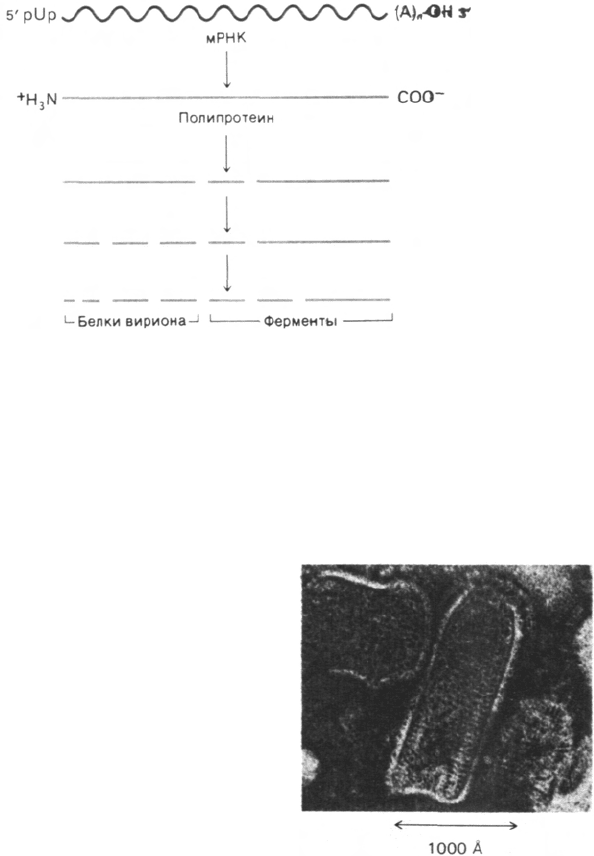
Рис. 30.22. Синтез белков вируса полио-
миелита путем множественно-
го расщепления гигантского
полипептидного предшествен-
ника.
капсида N массой 50 кДа - основного белка
вириона. ВВС заключен в липидную дву-
слойную мембрану, которую он захваты-
вает из плазматической мембраны в процес-
се отпочковывания от клетки (рис. 30.23).
Белок G (от англ. glycoprotein - гликопро-
теин) массой 65 кДа, кодируемый вирусом,
образует шипы, выступающие из этой мем-
браны. Белок матрикса М массой 29 кДа
располагается между оболочкой и нуклео-
капсидом. Эти пять белков ВВС образуются
при трансляции пяти (+)мРНК, а не пу-
тем расщепления одного полипротеина. Та
же самая РНК-репликаза синтезирует
и длинную (+)РНК, содержащую всю гене-
тическую информацию вируса. Эта полная
(+)РНК в свою очередь служит матрицей
для синтеза (—)РНК, которая упаковывает-
ся и дает новые вирионы.
30.13. Геном реовируса состоит из десяти
различных молекул двухцепочечной РНК
Реовирус, содержащий двухцепочечную
РНК, поражает клетки млекопитающих
и представляет третий тип вирусных генети-
ческих систем. Сердцевина вириона содер-
жит десять различных двухцепочечных мо-
лекул (±)РНК, ассоциированных с белка-
ми. Проникнув в клетку-хозяина, вирион
теряет наружную икосаэдрическую оболоч-
ку, состоящую из трех видов белков. Удале-
ние этой оболочки активирует РНК-поли-
меразу, содержащуюся в сердцевине вирио-
на. Эта РНК-зависимая полимераза по-
лностью транскрибирует 10 молекул
(+)РНК, так что образующиеся (+)мРНК
имеют такую же длину, как фрагменты ге-
Рис. 30.23. Электронная микрофотогра-
фия вируса везикулярного сто-
матита. (Williams R. С., Fisher
Н. W., Electron Micrographic
Atlas of Viruses, С. С. Thomas,
1974. Печатается с любезного
разрешения издателя.)
30. Вирусы
181

Реовирус -
вирус, содержащий двухцепочечную РНК, выделен из дыхательных путей и же-
лудочно-кишечного тракта людей и других млекопитающих; насколько извест-
но, он не вызывает заболевания. Приставка рео составлена из первых букв ан-
глийских слов respiratory enteric orphan, что означает энтеро-респираторный
вирус-сирота (сирота, поскольку бродит «не пристроенный» ни к какой
болезни).
Рис. 30.24. Электронная микрофотогра-
фия реовируса. (Williams R. С.,
Fisher H. W., An electron
micrographic atlac of viruses,
С. С. Thomas, 1974. Печатается
с любезного разрешения изда-
теля.)
Рис. 30.25. Синтез мРНК в сердцевине ре-
овируса. Молекулы мРНК вы-
глядят нитями, отходящими от
темных телец-сердцевин.
[Bartlett N. М., Gillies S. G.,
Bullivant S., Bellamy A. R.,
J. Virol., 14, 324 (1974).]
182
Часть IV.
Информация
нома. (±)РНК матрица транскрибируется
асимметрично и консервативно, т.е. обра-
зуются только (+)РНК, а исходные
(±)РНК разрушаются. В сердцевине вирио-
на к 5'-концам этих мРНК присоединяются
«колпачки» под действием ферментов. За-
тем эти концы выходят через каналы в серд-
цевине (рис. 30.25). Следовательно, сердце-
вина - высокоорганизованная система син-
теза мРНК. Каждая из этих десяти мРНК
дает при трансляции один белок. Затем
весь набор десяти (+)РНК соединяется
с некоторыми вирусными белками и об-
разует предшественник сердцевины («пре-
кор»), в котором синтезируется десять
(—)цепей.
Для чего геном реовируса разделен на
фрагменты? Как уже было указано выше,
вирусы животных не могут иметь полици-
стронных мРНК. Вирус полиомиелита ре-
шает эту проблему путем расщепления ги-
гантского белка-предшественника, а вирус
везикулярного стоматита транскрибирует
РНК вириона в виде коротких мРНК, ка-
ждая из которых соответствует одному бел-
ку. Стратегия реовируса состоит в том,
чтобы иметь отдельную «хромосому» для
каждого синтезируемого белка.
Четвертый путь выражения генетической
информации РНК-содержащих вирусов -
использование ДНК-посредника, интегри-
рующей с геном клетки-хозяина. Это более
сложная генетическая система, которую ис-
пользуют ретровирусы (РНК-содержащие
опухолеродные вирусы). Мы обсудим ее ни-
же в этой главе (разд. 30.19).
30.14. Мелкие РНК-содержащие фаги
содержат перекрывающиеся гены
Такие РНК-содержащие фаги, как R17 (MS2,
F2) и Q
β
, относятся к простейшим вирусам.
Они имеют форму правильного многогран-
ника и диаметр около 200 А. Капсид этих
близкородственных фагов содержит 180 мо-
лекул белка оболочки массой 14 кДа и одну
молекулу белка А (созревания) массой
38 кДа. Кроме того, одноцепочечная
(+)РНК кодирует одну из субъединиц ре-
пликазы. До сих пор считалось, что эти мел-
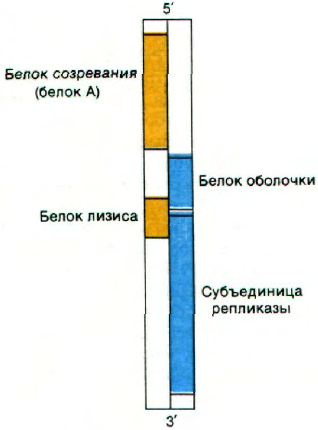
кие РНК-содержащие вирусы содержат
только три гена. Однако обнаружение му-
танта фага, образующего нормальные ви-
рионы, но не способного при этом лизиро-
вать клетку-хозяина, повлекло за собой
поиск еще одного кодируемого вирусом
белка. Действительно, у РНК-содержаших
фагов имеется четвертый ген, кодирующий
белок, необходимый для лизиса бактерии-
хозяина. Ген этого белка лизиса перекры-
вается с генами белка оболочки и субъеди-
ницы репликазы (рис. 30.26). Эти мелкие
РНК-содержащие фаги, подобно маленько-
му ДНК-содержащему фагу φX174
(разд. 26.11), используют перекрывающиеся
гены для того, чтобы вместить больше ин-
формации в свои маленькие геномы. Молеку-
ла (+)РНК вириона служит матрицей и для
синтеза четырех белков, и для синтеза
(—)РНК. Затем (—)РНК используется в ка-
честве матрицы для образования множества
копий (+)РНК. Таким образом, по генети-
ческой системе эти фаги напоминают вирус
полиомиелита.
Репликаза, синтезирующая (+)- и (—)це-
пи фаговой РНК,- очень интересный фер-
мент. Он проявляет высокую специфич-
ность к гомологичной фаговой РНК. Поэто-
му молекулы РНК клетки-хозяина не конку-
рируют с фаговой РНК при репликации.
Qβ-репликаза состоит из четырех субъеди-
ниц, из которых только одна кодируется фа-
говой РНК. Три другие субъединицы репли-
казы - белки клетки-хозяина, которые фаг
приспособил для собственных нужд. Два из
них - факторы элонгации синтеза белка
EF-Tu и EF-Ts, а третий - один из компонен-
тов 30S-субчастицы рибосомы. Так фаг Qβ
создает весьма специфичный фермент мак-
симально экономичным способом.
Регуляторные механизмы обеспечивают
правильную временную последовательность
трансляции и репликации. (+)РНК служит
одновременно матрицей для синтеза белка
и для синтеза (—)РНК. Было бы нежела-
тельно, чтобы оба процесса происходили
одновременно на одной и той же (+)РНК,
поскольку рибосомы, движущиеся в на-
правлении 5'—>3', сталкивались бы с репли-
казой, движущейся в направлении 3'—>5'.
Этого не происходит, так как Qβ-репликаза
сильно ингибирует связывание рибосом с
(+)РНК до тех пор, пока не синтезируется
достаточное число молекул (—)РНК.
Четыре фаговых белка синтезируются
в различных количествах. Белок оболочки,
который синтезируется на протяжении всего
Рис. 30.26. Перекрывающиеся гены в РНК
вирусов R17 и Qβ. Одна рамка
считывания показана желтым
цветом, другая - синим.
периода инфекции,- основной продукт
трансляции. Одна из причин этого состоит
в том. что рибосомы гораздо прочнее связы-
ваются с участком инициации соответ-
ствующего цистрона, чем с другими участ-
ками инициации в (+)РНК. Кроме того,
белок оболочки подавляет трансляцию гена
репликазы, блокируя его участок инициа-
ции. Таким образом, белок оболочки - спе-
цифический релрессор трансляции. Белок
А транслируется только с незаконченных
молекул (+)РНК, так как в полных молеку-
лах РНК его участок инициации блокирован
в результате спаривания оснований. Вторич-
ная структура полной молекулы РНК по-
зволяет транслировать лишь небольшое ко-
личество белка А.
30.15. Дарвиновская эволюция фаговой РНК
вне клетки
Очистка РНК фага Qβ и Qβ-репликазы от
примесей нуклеаз позволила Солу Спигел-
ману (Sol Spigelman) изучать эволюционные
события вне живой клетки. Один из вопро-
сов был сформулирован следующим обра-
зом: что произойдет с молекулами РНК, ес-
30. Вирусы
183
ли единственное предъявляемое к ним требо-
вание - это как можно быстрее размно-
жаться? Молекулы РНК и репликазы фага
Qβ и рибонуклеозидтрифосфаты инкубиро-
вали в течение 20 мин. Такое время инкуба-
ции способствует отбору мутантных моле-
кул РНК, которые быстро реплицируются.
Образец этой инкубационной смеси перено-
сили и разводили в свежей порции стандарт-
ной реакционной смеси, содержащей Qβ-ре-
пликазу и рибонуклеозидтрифосфаты. Про-
водили 75 переносов и затем анализировали
образовавшиеся РНК. Продолжительность
инкубации постепенно снижали, так как мо-
лекулы РНК на протяжении эксперимента
реплицировались все быстрее. Самым удиви-
тельным было то, что после 75 переносов
(«поколений») длина молекул РНК соста-
вляла всего 12% длины исходной РНК фага
Qβ. Нуклеотиды, ненужные для репликации,
были утрачены, так как укороченные моле-
кулы реплицировались быстрее. Основное
ограничение, которое накладывали условия
этого эксперимента,- сохранение в му-
тантных молекулах инициирующей после-
довательности, которую узнает Qβ-репли-
каза.
30.16. Лизогенные фаги могут включать
свою ДНК в состав ДНК клетки-хозяина
У некоторых бактериофагов существует два
возможных пути, по которым может пойти
их дальнейшее развитие после заражения
клетки-хозяина: они могут размножаться
и лизировать зараженную клетку (литиче-
ский путь развития) или же их ДНК может
включиться в ДНК зараженной клетки, не
проявляя способности к размножению и ли-
зису (лизогенный путь развития). Вирусы,
которые не всегда убивают клетку-хозяина,
называются умеренными. Лучше всего из
умеренных вирусов изучен фаг
λ (рис. 30.27); мы уже говорили о нем рань-
ше в связи с регуляцией транскрипции
(разд. 28.11). Напомним, что репрессор фага
λ связывается с двумя группами опера-
торных участков O
L
и O
R
и что он регули-
рует свой собственный синтез.
ДНК вириона λ - линейная двухцепочеч-
ная молекула длиной 48 kb. 5'-конец каждой
ее цепи представляет собой одноцепочеч-
ную последовательность из 12 нуклеотидов.
Эти последовательности называются липки-
184
Часть IV.
Информация
Рис. 30.27. Электронная микрофотогра-
фия фага λ. (Печатается с лю-
безного разрешения д-ра
A. Dale Kaiser.)
ми концами, так как они взаимно компле-
ментарны и могут спариваться друг с дру-
гом. На самом деле они соединяются сразу
после заражения. В результате 5'-фосфат ка-
ждой цепи оказывается рядом со своим
собственным 3'-гидроксильным концом.
ДНК-лигаза клетки-хозяина заделывает
разрывы, и в результате образуется кольце-
вая молекула ДНК фага λ (рис. 30.28).
Репликация этой кольцевой молекулы
ДНК фага λ происходит путем взаимодей-
ствия белков, кодируемых фагом λ, с репли-
кационными механизмами клетки-хозяина.
В другом случае кольцевая ДНК фага λ мо-
жет включиться в бактериальную хромосо-
му с помощью одного акта реципрокной ре-
комбинации между специфическими участ-
ками ДНК фага λ и E.coli длиной по 15 пар
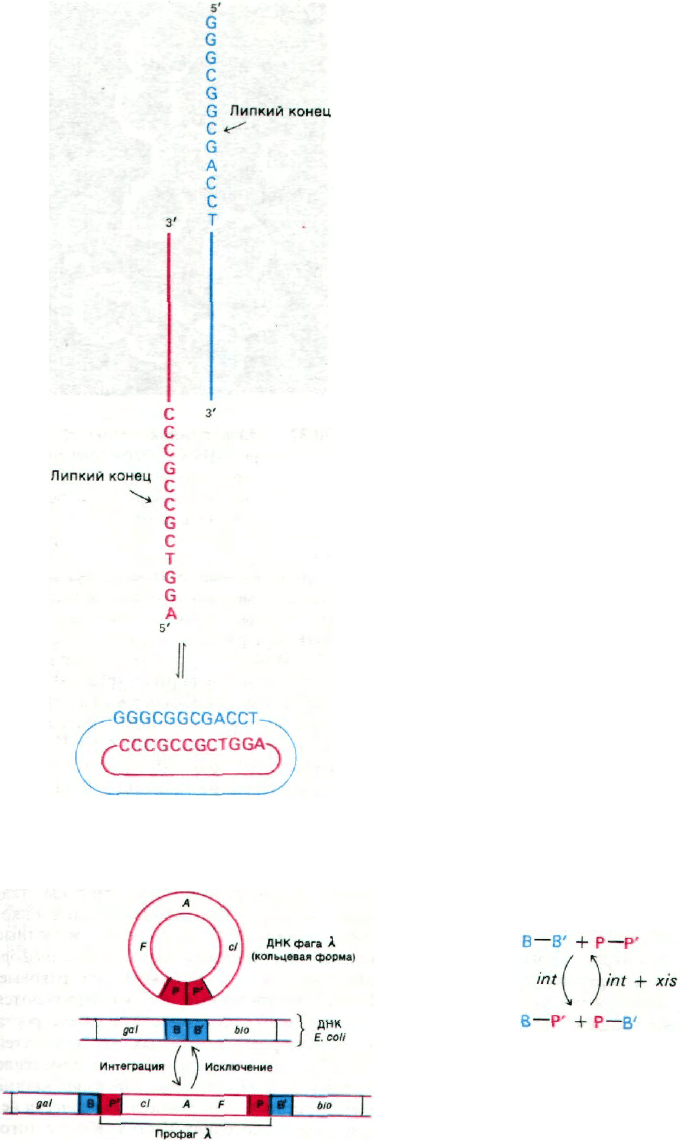
Рис. 30.28. Превращение линейной ДНК
фага λ в кольцевую форму.
Рис. 30.29. Схема реципрокной рекомби-
нации ДНК фага λ и ДНК Е.
coli.
оснований. Сайт (участок) прикрепления фа-
га λ в ДНК E.coli обозначается attλ и распо-
лагается между генами галактозного и био-
тинового оперонов galE и biоА. Последова-
тельность оснований attλ можно символи-
чески обозначать В-В' (от англ.
bacterial - бактериальный). Специфический
сайт прикрепления в фаге λ называется att
и локализован рядом с генами int (от англ.
integrate - интегрировать) и xis (от англ.
excise - вырезать). Последовательность ос-
нований att обозначается Р-Р' (от англ.
phage - фаг). Белок int узнает последователь-
ность Р-Р' в фаговой ДНК и последователь-
ность В-В' в ДНК E.coli. Затем происходит
взаимный перенос: Р соединяется с В', а В - с
P'. Эта схема (рис. 30.29 и 30.30) была перво-
начально предложена Алланом Кэмпбел-
лом (Allan Campbell) на основании генетиче-
ских данных.
Теперь ДНК фага λ составляет часть мо-
лекулы ДНК E.coli. Эта форма называется
профагом, а клетка E.coli, содержащая про-
фаг,- лизогенной бактерией. Профаг стаби-
лен в отсутствие белка xis. Транскрипция ге-
на xis блокируется репрессором фага
λ (разд. 28.11). Когда репрессия снимается.
белки xis и int совместно катализируют раз-
рыв последовательностей В—Р' и Р—В',
и снова происходит взаимный перенос
(рис. 30.30): Р соединяется с Р', а В с В'; при
этом снова получаются кольцевая молекула
ДНК фага λ и нелизогенная хромосома
E.coli. Главная особенность этой системы
рекомбинации заключается в том. что белок
int сам по себе не может узнавать две новые
последовательности на концах профага
(В-Р' и Р-В'), поэтому они устойчивы. Та-
ким образом, внедрение фага происходит
в присутствии одного белка int, тогда как
вырезание профага - только в присутствии
обоих белков - int и xis.
Рис. 30.30. Интеграция и исключение
ДНК фага λ. Это неполная схе-
ма, так как некоторые факторы,
участвующие в этих процессах,
пока не идентифицированы.
30. Вирусы
185
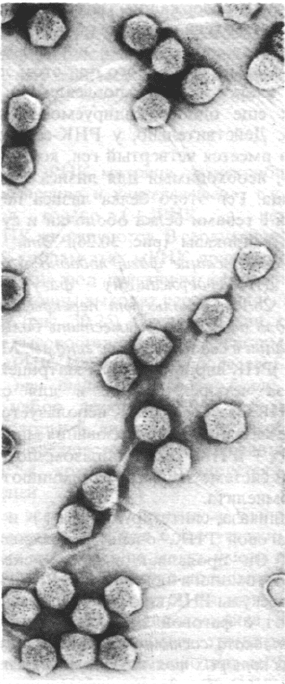
30.17. Ретровирусы и некоторые ДНК-
содержащие вирусы могут вызывать рак
у чувствительных клеток-хозяев
В 1911 г. Пейтон Раус (Peyton Rous) приго-
товил бесклеточный фильтрат из опухоли
соединительной ткани, которая спонтанно
возникла у курицы и ввел его нормальным
цыплятам. Как ни странно, у реципиентов
развились высокозлокачественные опухоли
такого же типа, которые называются сарко-
мами. Кроме того, Раус обнаружил, что опу-
холеродный фактор фильтрата, известный
в настоящее время под названием вируса
саркомы Рауса (RSV) или вируса саркомы
птиц (ASV) (рис. 30.31), можно размножить
путем последовательного пассирования
в курах. Вирус саркомы птиц относится
к группе РНК-содержащих опухолеродных
вирусов (онкогенных РНК-содержащих ви-
русов). Эти вирусы содержат в составе вири-
нов (+)РНК и размножаются с использова-
нием двухспиральной ДНК-посредника. По-
этому их называют ретровирусы. Ретрови-
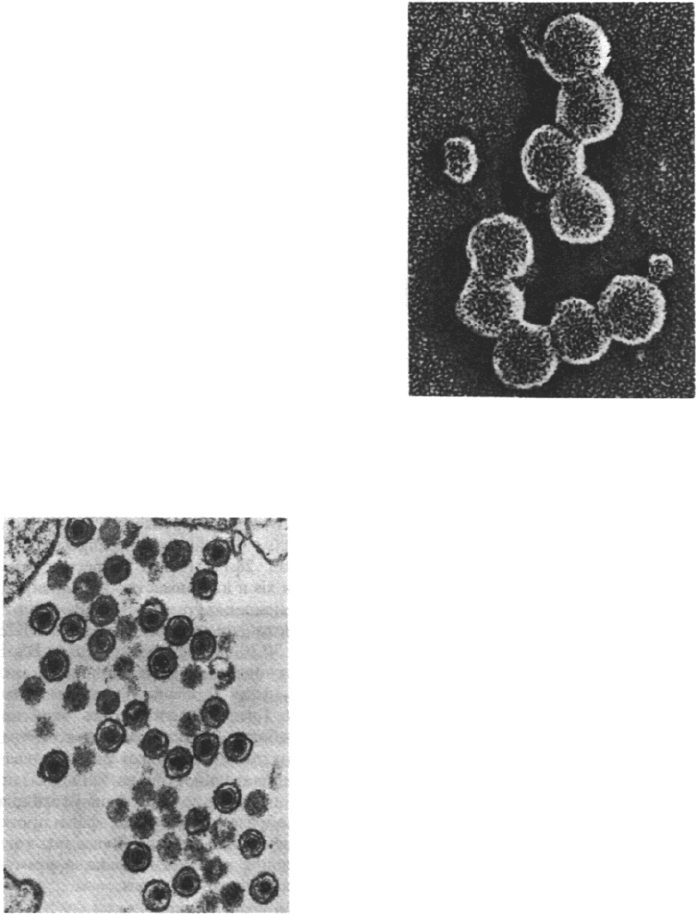
Рис. 30.31. Электронная микрофотогра-
фия вируса саркомы птиц (ви-
руса саркомы Рауса), относя-
щегося к ретровирусам. Силь-
ноокрашивающиеся вирионы
расположены вблизи поверх-
ности зараженной клетки цы-
пленка. (Печатается с любезно-
го разрешения д-ра Samuel
Dales.)
186
Часть IV.
Информация
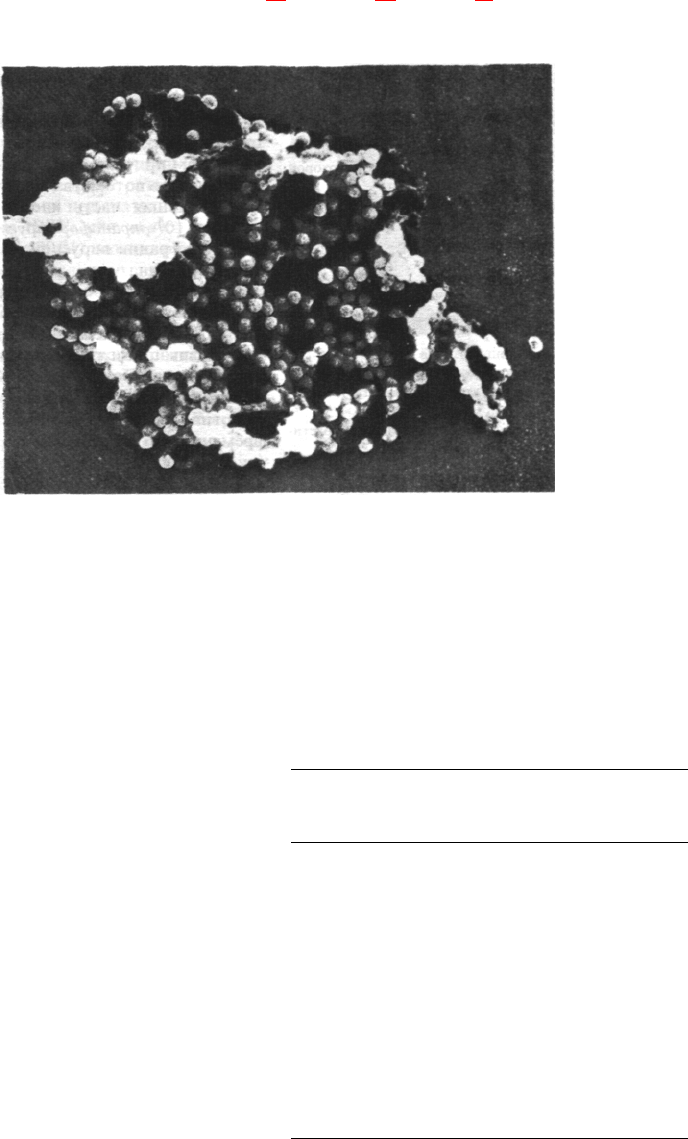
Рис. 30.32. Электронная микрофотогра-
фия ДНК-содержащего опухо-
леродного вируса SV-40. (Печа-
тается с любезного разрешения
д-ра Jack Griffith.)
русы - единственные РНК-содержащие ви-
русы, способные вызывать рак. Кроме того,
злокачественные опухоли может вызывать
ряд ДНК-содержащих вирусов. Обезьяний
вирус 40 (SV-40) и вирус полиомы принадле-
жат к группе паповавирусов (рис. 30.32); из
всех онкогенных ДНК-содержащих вирусов
они изучаются наиболее интенсивно.
Особый интерес к этим РНК- и ДНК-содер-
жащим вирусам привлекает то, что они со-
держат всего четыре-пять генов. В индукции
рака участвует всего один или два вирусных
гена, и потому исследователи питают на-
дежду выяснить механизм их действия.
Для изучения рака на молекулярном
уровне были разработаны системы тка-
невых культур. При проникновении онко-
генного вируса в подходящие животные
клетки они становятся постоянно трансфор-
мированными, т.е. похожими на раковые.
Трансформированные клетки отличаются
от нормальных особенностями своего роста
и характером клеточных поверхностей
(табл. 30.3). Самое разительное изменение
состоит в том, что трансформированные
клетки растут непрерывно и хаотически не-
зависимо от соседних клеток. Кроме того,
трансформированные клетки содержат ви-
русоспецифическую ДНК, интегрированную
с геномом клетки-хозяина. Этим объясняет-

Таблица 30.3. Изменения свойств клеток при трансфор-
мации ДНК- или РНК-содержащими опухолероднымн ви-
русами (Tooze J., ed.. The molecular biology of tumour
viruses. Cold Spring Harbor Laboratory, 1973, p. 351)
Характер роста
При введении чувствительным животным образуют
опухоли
Растут до гораздо более высокой плотности
Рост становится не ориентированным в пространстве
и клетки отделяются от поверхности, на которой
растут
В среду выделяются активаторы протеаз, что повы-
шает инвазивность клеток
Снижается потребность в факторах роста сыворотки
Свойства поверхности
Появляются новые вирусоспецифические антигены
Один из белков наружной поверхности клеток - фи-
бронектин - исчезает
Падает содержание ганглиозидов
На поверхности клетки появляются антигены плода
Скорость транспорта питательных веществ увеличи-
вается
Способность к агглютинации под действием расти-
тельных лектинов увеличивается
Признаки присутствия опухолеродного вируса
Имеются последовательности вирусной ДНК
Имеются вирусоспецифические мРНК
Выявляются вирусоспецифические антигены
ся тот факт, что трансформация является
наследуемым изменением фенотипа. Куль-
туры, полученные из трансформированных
колоний, навсегда сохраняют аномальные
свойства трансформированных клеток. Кро-
ме того, некоторые трансформированные
клетки, полученные из культуры ткани, при
введении в достаточном количестве в подхо-
дящего хозяина образуют раковую опухоль.
30.18. Вирусы SV-40 и полиомы могут
вызывать продуктивную инфекцию
или трансформацию клеток-хозяев
Вирусы SV-40 и полиомы содержат внутри
икосаэдрической оболочки маленькую коль-
цевую двухспиральную ДНК. В некоторых
клетках (они называются пермиссивными
клетками-хозяевами) развитие этих вирусов
идет по пути литического цикла, что приво-
дит к образованию множества новых вирио-
нов (рис. 30.33).
При продуктивной инфекции эти вирусы
убивают клетки. В клетках других типов (не-
пермиссивных клеток-хозяев) некоторые
стадии экспрессии вирусного генома по не-
понятным причинам блокируются. Никако-
го вирусного потомства в них не образуется.
но небольшая часть клеток - порядка од-
ной на 10
5
- трансформируется в резуль-
тате интеграции вирусной ДНК с геномом
клетки-хозяина.
К настоящему времени расшифрована
полная последовательность 5243 пар осно-
ваний ДНК вируса SV-40, а многие аспекты
его репликации и транскрипции интенсивно
исследуются. Половина ДНК транскриби-
руется на ранней стадии инфекции, другая
половина ДНК - на поздней стадии, одно-
временно с синтезом вирусной ДНК
(рис. 30.34).
Точка начала репликации находится в той
же области, что и начало транскрипции ран-
ней и поздней областей. Ранняя область
транскрибируется в направлении против ча-
совой стрелки и кодирует Т-антиген (белок
А), необходимый для инициирования репли-
кации ДНК. Другой, иммунологически от-
личный белок - малый t-антиген - также ко-
дируется ранней областью. При синтезе
мРНК для Т-антигена происходит выре-
зание вставочной последовательности из
первичного транскрипта.
Таким образом, вирус SV-40 использует
аппарат сплайсинга клеточного ядра. Заслу-
живает внимания также последовательность
оснований в точке начала репликации. Здесь
имеются две последовательности длиной по
13 пар оснований, симметричных относи-
тельно оси второго порядка, а рядом нахо-
дится АТ-богатая область:
30. Вирусы
187
Паповавирусы -
группа ДНК-содержащих вирусов. Название составлено по названиям трех
представителей группы: вирусов папилломы, полиомы и вакуолизирующего ви-
руса (SV-40).
Рис. 30.33. Электронная микрофотогра-
фия фрагмента ядерной мем-
браны клетки, зараженной ви-
русом SV-40. Видны ядерные
поры и множество вирионов.
(Печатается с любезного разре-
шения д-ра Jack Griffith.)
С этим участком связывается Т-антиген.
Транскрипция поздней области происхо-
дит по направлению часовой стрелки от на-
чала репликации (рис. 30.34) и приводит
к синтезу трех белков капсида: VP1, VP2
и VP3. И в этом случае происходит удаление
вставочных последовательностей из первич-
ного транскрипта. Три мРНК, по-видимо-
му, образуются в результате различных ре-
акций сплайсинга. N-концевая последова-
тельность аминокислот VP3 перекрывает
70% С-концевой последовательности VP2.
Кроме того, перекрывающийся участок из
22 нуклеотидов читается в одной рамке
считывания при синтезе VP2 и VP3 и в дру-
гой рамке при синтезе VP1. Так, ограничен-
ное количество генетической информации
у вируса SV-40 используется с максимальной
188
Часть IV.
Информация
эффективностью. Еще один пример генети-
ческой экономии - то, что SV-40 не синтези-
рует собственных белков для упаковки
ДНК. Новосинтезированная ДНК свя-
зывается с гистонами клетки-хозяина
(рис. 30.35). Затем этот сверхспирализован-
ный комплекс упаковывается в капсид с по-
мощью белков VP1, VP2 и VP3. В конце кон-
цов вирионы потомства высвобождаются
Таблица 30.4. Белки, кодируемые вирусом SV-40
Белок
Т-антиген
(белок А)
t-Антиген
VP1
VP2
VP3
Масса,
кДа
94
21
40
39
27
мРНК
Ранняя
Ранняя
Поздняя
Поздняя
Поздняя
Роль
Необходим для
инициации, репли-
кации ДНК и для
трансформации
Неизвестна
Основной белок
капсида
Минорный белок
капсида
Минорный белок
капсида
при лизисе клетки-хозяина, которая в ре-
зультате гибнет.
В непермиссивных клетках происходит
экспрессия ранней области генома SV-40,
поздняя же область не реплицируется и не
транскрибируется. Небольшая часть этих
клеток трансформируется в результате ин-
теграции генома SV-40 с клеточной ДНК.
В отличие от интеграции ДНК фага λ в ме-
ханизме интеграции ДНК SV-40, по-види-
мому, не участвуют специфические участки
ни вирусной, ни клеточной ДНК. Для транс-
формации, а также, по всей вероятности, для
поддержания трансформированного состоя-
ния необходимы оба вирусных антигена (Т
и t). Введение таких трансформированных
клеток чувствительным животным приво-
дит к быстрому образованию опухолей. Ос-
новная цель ведущихся в настоящее время
исследований вируса SV-40 состоит в том,
чтобы выяснить, как экспрессия ранней
области интегрированной формы этого ви-
руса делает клетку раковой.
30.19. Ретровирусы содержат обратную
транскриптазу, которая синтезирует
двухспиральную ДНК, используя
в качестве матрицы (+)РНК
Еще один класс опухолеродных вирусов -
ретровирусы - содержит (+)РНК-геном в
икосаэдрическом «футляре». Это сфериче-
ское нуклеопротеиновое «ядро» окружено
оболочкой, состоящей из кодируемых виру-
сом молекул гликопротеина в двуслойной
липидной оболочке, происходящей из плаз-
матической мембраны клетки-хозяина.
Обычно диаметр ретровирусов составляет
1000 А (см. рис. 30.31).
В 1964 г. Говард Темин (Howard Temin)
наблюдал, что заражение такими РНК-со-
держащими опухолеродными вирусами, как
вирус саркомы птиц, блокируется ингибито-
рами синтеза ДНК. Ингибиторы, например
аметоптерин, 5-фтордезоксиуридин и цито-
зинарабинозид, эффективны в течение
первых двадцати часов после введения виру-
сов. В результате этого открытия было
высказано предположение, что для роста
РНК-содержащих опухолеродных вирусов
необходим синтез ДНК. К тому же образо-
вание частиц вирусного потомства подав-
ляется актиномицином D. Известно, что
этот антибиотик ингибирует синтез РНК по
ДНК-матрице (разд. 25.18). Так зародилось
предположение о том. что для размножения
РНК-содержащих опухолеродных вирусов
Рис. 30.34. Генетическая карта ДНК виру-
са SV-40, содержащая 5243
пары оснований. Ранняя
область (транскрибируется
в направлении против часовой
стрелки) показана желтым цве-
том, поздняя область (тран-
скрибируется по часовой стрел-
ке) - синим, место начала ре-
пликации (ori) - красным.
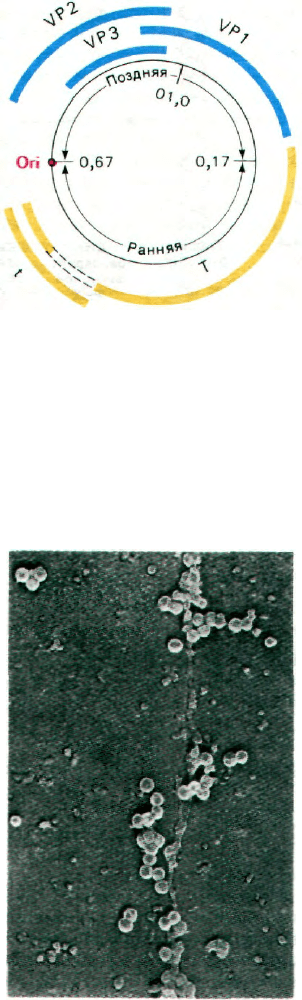
Рис. 30.35. Электронная микрофотогра-
фия формирующихся частиц
опухолеродного вируса SV-40,
ассоциированных с клеточной
хромосомой. (Печатается с лю-
безного разрешения д-ра Jack
Griffith.)
30. Вирусы
189
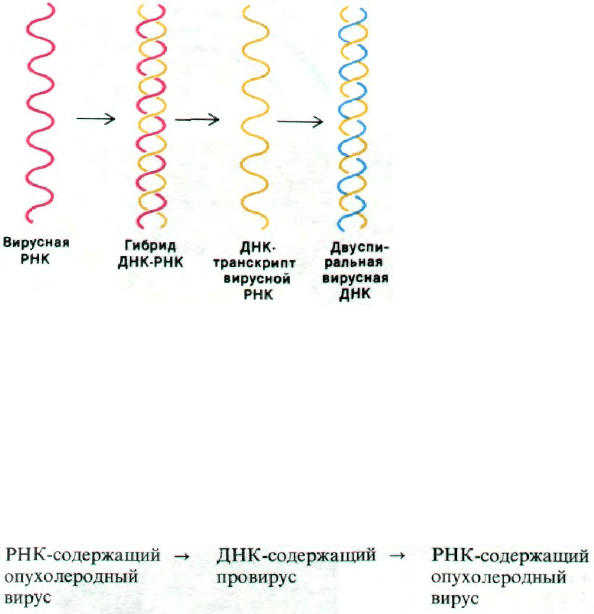
Рис. 30.36. Синтез ДНК по РНК-матрице
обратной транскриптазой. За-
травка не показана.
необходима транскрипция ДНК. Эти неожи-
данные данные привели Темина к мысли,
что ДНК-содержащий провирус служит
промежуточным продуктом в репликации
и в онкогенном действии этих вирусов:
Выдвинутая Темином гипотеза, что генети-
ческая информация может переходить от
РНК к ДНК, была вначале холодно встрече-
на большинством исследователей. Она тре-
бовала существования неизвестного тогда
еще фермента, способного синтезировать
ДНК по РНК-матрице (РНК-зависимой
ДНК-полимеразы). В 1970 г. Темин и Балти-
мор (Temin, Baltimore) независимо открыли
такой фермент, который называется обрат-
ной транскриптазой, в вирионах некоторых
РНК-зависимых опухолеродных вирусов.
Все вирусы этой группы, исследованные
в дальнейшем, содержали обратную тран-
скриптазу, поэтому их и называют ретрови-
русами (от англ. reverse transcriptase - обрат-
ная транскриптаза).
Жизненный цикл обычного ретровируса
начинается со связывания вирионов со спе-
цифическими рецепторами на поверхности
клетки-хозяина и проникновения в клетку.
В цитозоле вирусная (+)РНК скидывает
оболочку. Затем обратная транскриптаза,
190
Часть IV.
Информация
содержавшаяся в вирусной частице, синтези-
рует (—)цепь ДНК. Тот же фермент расще-
пляет цепь геномной РНК в составе гибрида
РНК—ДНК. Теперь обратная транскрипта-
за синтезирует (+)цепь ДНК, используя
в качестве матрицы (—)цепь. Таким обра-
зом, обратная транскриптаза осущест-
вляет три последовательные реакции: РНК-
зависимый синтез ДНК, гидролиз РНК
и ДНК-зависимый синтез ДНК (рис. 30.36).
Подобно другим ДНК-полимеразам,
обратная транскриптаза синтезирует ДНК
в направлении 5'—>3' и неспособна к инициа-
ции цепей de novo. Откуда же берется за-
травка для синтеза вирусной ДНК? Процесс
инициации весьма экономичен: (+)РНК ви-
русного генома содержит нековалентно свя-
занную транспортную РНК (в вирусе сар-
комы птиц это - триптофановая тРНК),
которая была захвачена в клетке-хозяине во
время предыдущего цикла заражения.
3'-ОН-группа этой тРНК, основания кото-
рой спарены с геномной РНК, действует
в качестве затравки синтеза ДНК. Как про-
исходит репликация полной цепи (+)РНК?
Напомним, что при репликации любой ли-
нейной ДНК возникает особая проблема за-
полнения 5'-концов (разд. 30.6). Ретрови-
русы нашли очень остроумный выход из
этого положения. Их геномы состоят не из
одной, а из двух молекул (+)РНК
(рис. 30.37). Эти молекулы связаны друг
с другом водородными связями вблизи
5'-концов. К тому же (+)РНК содержит од-
ну и ту же последовательность у 5'- и
у 3'-конца. Эта концевая избыточность, по-
видимому, необходима для процесса репли-
кации, как и в случае фага Т4 (разд. 30.6).
30.20. Ретровирусная ДНК
транскрибируется только в том случае, если
она интегрирована с геном клетки-хозяина
Двухспиральная вирусная ДНК переходит
в кольцевую форму и проникает в ядро.
Транскрипция ретровирусной ДНК проис-
ходит только после того, как она интегри-
рует с ДНК клетки-хозяина. Таким образом,
в жизненном цикле ретровирусов интегра-
ция — этап обязательный. В жизненном ци-
кле онкогенных ДНК-содержащих вирусов,
напротив, интеграция и продуктивная ин-