Санагурський Д.І. Об’єкти біофізики: монографія
Подождите немного. Документ загружается.


101
деполяризуючого впливу адреналіну може ґрунтуватися на змінах
функціонування йонтранспортних систем. Добре відомо про вплив
інсуліну на ці системи у диференційованих клітинах (С.И. Кусень,
Р.С. Стойка, 1985; M.P. Czech, 1977). В огляді Мура (R.D. Moore,
1983) узагальнено дані про різноманітні транспортні системи, які
можуть опосередковувати вплив інсуліну на перерозподіл іонів у
клітині. Гормон збільшує активність Na
+
, K
+
-ATФ-ази, та ця дія
незалежна від білкового синтезу і відбувається при фізіологічних
концентраціях інсуліну. Він також впливає на транспортування
йонів кальцію, інгібуючи Ca
2+
, Mg
2+
-АТФ-азу, що веде до
збільшення обміну йонів натрію на кальцій (R.D. Moore, 1983;
N.E. Owen, M.L. Villereal, 1983).
Виявлено, що при гальмуванні мітозів цитостатиком
колхіцином бракує параболічного росту рівня ТМП, характерного
для контрольного досліду, але зберігаються його періодичні
коливання з видовженим періодом та значно збільшеною
амплітудою ніж у нормі (Е.А. Гойда, 1993; Е.А. Гойда и др., 1981).
Відомі також повідомлення, що хвилі скорочення кортекса за цих
умов відбуваються з тим самим ритмом, що й у нормі, а
ізольований кортекс містить кальмодулін (M. Ishikawa, T. Kohno,
1984)
Наведені факти засвідчують те, що динамічні зміни
біоелектричних процесів у період раннього ембріогенезу тварин
тісно спряжені з метаболічними процесами, що відбуваються в
цитоплазмі, і теж характеризуються періодичністю. Ця
спряженість, вочевидь, підтримується різними регуляторними
факторами.
102
2.11. ijÿ êîëõ³öèíó íà âåëè÷èíó
³ õàðàêòåð êîëèâàíü ÒÌÏ
çàðîäê³â â’þíà
Як описано попередньо, температурний
фактор має великий вплив на темп дроблення бластомерів у в’юна.
Було цікаво прослідкувати, як позначиться гальмування утворення
нових мембран ембріональних клітин на генеруванні ТМП. Для
цієї мети було використано колхіцин, який є хорошим
цитостатиком (А.И. Зотин, 1961). Він інгібує процес розбіжності
хромосом і утворення борозен і мембран нових клітин, не порушує
при цьому перебігу інших основних ланок метаболізму в клітинах
(А.И. Зотин, 1961).
У наших дослідах при концентраціях 0,9·10
-3
і 4,5·10
-3
М
(рис. 2.22, криві 3, 2) колхіцин спричиняє початкову швидку
гіперполяризацію мембрани, що утримується потім упродовж
перших двох–двох з половиною год розвитку зародків. Разом з тим
періодичні коливання ТМП, які простежувалися, мають більш
розтягнутий за тривалістю період і збільшену амплітуду коливань,
порівняно з контролем.
Потрібно відзначити і те, що в разі впливу колхіцину у
зазначених концентраціях немає характерного для контролю спаду
ТМП на шостій годині розвитку зародка. Мабуть, зародки в’юна не
встигають виконати ввесь генетично запрограмований часовий
план, який включає, очевидно, i певні метаболічні перетворення,
впливають на потенціалгенеруючу систему клітин.
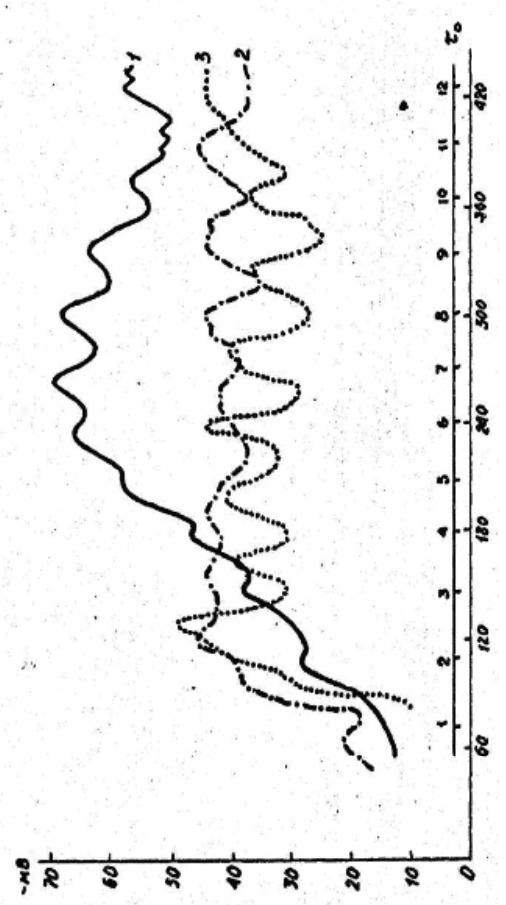
103
Рис. 2.22. Вплив різних концентрацій колхіцину на динаміку ТМП зародків в’юна, що розвиваються
хв

104
Цікавим є те, що колхіцин усуває параболічне наростання
величин ТМП, характерне для контролю. Це, швидше за все,
пов’язано з постійною площею клітинних мембран, оскільки
клітинне ділення гальмується.
Двогорбі ж піки при нижчих концентраціях (рис. 2.22,
крива 2) колхіцину є частковим феноменом, якому немає
переконливого пояснення. Щоправда, можна припустити, що в
останньому прикладі певне значення має час поміщення зародків у
середовище, що містить цитостатик.
Отже, колхіцин, не усуваючи періодичних коливань,
обмежує ТМП у максимальних величинах тих, що не перевищують
-35÷10 мВ. Ураховуючи обставину, що основні обмінні реакції
ембріональних клітин мало порушуються колхіцином (А.И. Зотин,
1961), можна припустити, що спостережувані при його дії
коливання ТМП є своєрідним віддзеркаленням запущених актом
запліднення періодичних змін інтенсивності синтетичних процесів
у цитоплазмі, які швидко змінюються один за одним відповідно до
мітотичних циклів бластомерів зародків в’юна, що дробляться в
нормі. Щоб упевнитися в цьому, нами була зроблена спроба
визначити динаміку ТМП у зародків в’юна при дії на них деяких
біологічно активних речовин з відомими точками впливу, що
призводить до пригнічення або активування окремих ланок
обміну речовин у клітині.

105
2.12. Õàðàêòåðèñòèêà ÒÌÏ
çàðîäê³â â'þíà çà íàÿâíîñò³
â ñåðåäîâèù³ ³íêóáàö³¿
ã³äðîêîðòèçîíó
Вплив гідрокортизону в концентрації
1·10 М на величину ТМП і його динаміку у зародків в’юна
зображено на рис. 2.23, крива 2. Визначати ТМП почали після
трьох годин після запліднення, а до цього зародки знаходилися в
розчині, що містив гормон. У цьому варіанті досліду змінюється не
тільки основний ритм періодичних коливань ТМП, порівняно з
контролем (рис. 2.23, крива 1), але й знижується його абсолютна
величина.
Відмічене нами раніше в контролі падіння величини ТМП
після п’яти годин ембріогенезу, при дії гідрокортизону, зрушується
за часовою шкалою і виражено як після шести годин розвитку. До
того ж, максимальне значення ТМП (–55 мВ) у разі дії
гідрокортизону досягається через 330 хвилин після запліднення,
тобто у момент відносно низького його значення при
нормальному розвитку зародків.
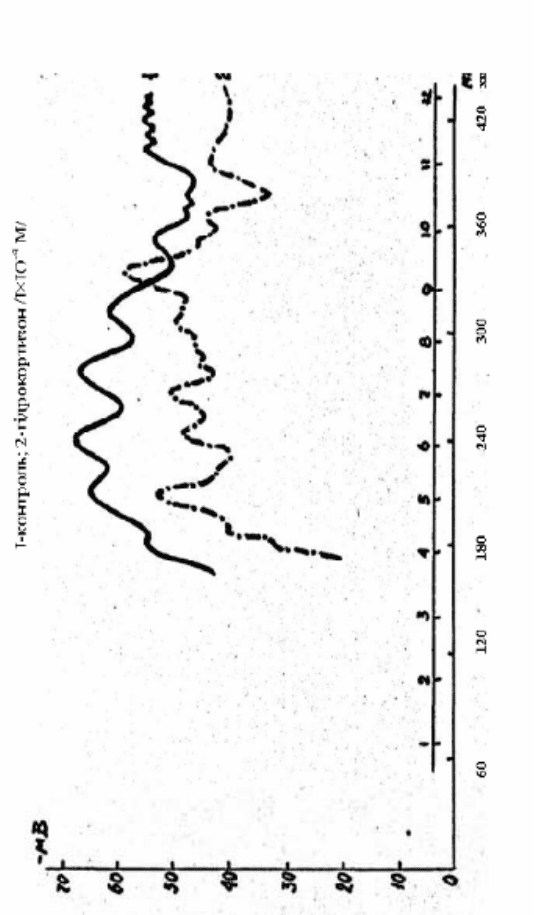
106
Рис. 2.23. Вплив гідрокортизону на ТМП зародків в’юна, що розвиваються

107
Таке зрушення коливань величин ТМП є, мабуть,
наслідком модифікації розвитку зародків в’юна гідрокортизоном.
Це засвідчують також дані про зменшення під впливом
гідрокортизону в зародках в’юна, активності деяких ферментів
(С.Й. Кусень і ін., 1974).
Крім того, при дії гідрокортизону спостерігається
аперіодична поява зубців на кривій зміни ТМП, що не виявляє
особливої закономірності. Створюється враження, що
гідрокортизон частково десинхронізує або затримує ділення
бластомерів зародків в’юна вже на початкових етапах розвитку, що
підтверджується візуальними і біохімічними спостереженнями
(С.И. Кусень, П.С. Олешко, 1974 ).
2.13. Âïëèâ ð³çíèõ
êîíöåíòðàö³é ³íñóë³íó
íà äèíàì³êó ÒÌÏ
ó ðàííüîìó ðîçâèòêó â'þíà
Дані, одержані в серіях дослідів з
інсуліном (рис. 2.24, криві 2, 3, 4), свідчать про специфічний вплив
інсуліну на величину ТМП, що залежить від концентрацій гормону
в середовищі. Так, при концентрації 0,1 МО/мл (рис. 2.24, крива 2)
інсулін спричиняє значну гіперполяризацію мембрани, чітко
виражену до кінця четвертої години розвитку зародків. Водночас
зберігається ритміка коливань ТМП, аналогічна тривалості періоду
мітотичного циклу синхронних дроблень бластомерів, проте
спостерігається деяке зрушення в настанні мінімумів і максимумів
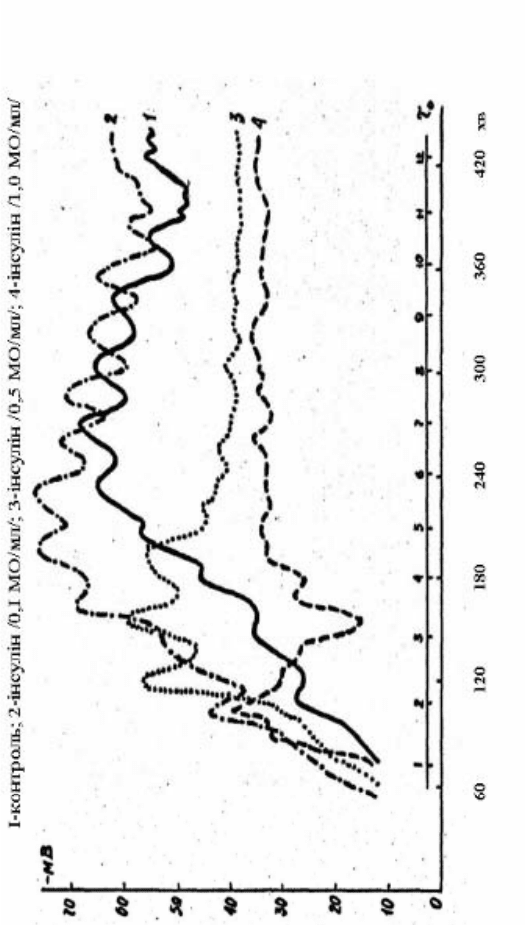
108
Рис. 2.24. Вплив різних концектрацій інсуліну на динаміку ТМП у ранньому розвитку в’юна
109
коливань, хоча періоди й амплітуди наближаються до таких у
нормі.
Зі зростанням концентрації гормону в середовищі інкубації
зародків в’юна також має місце гіперполяризація мембрани
протягом двох–трьох з половиною годин початкового етапу
їхнього розвитку. Надалі рівень ТМП знижується, коливання
зменшуються за амплітудою, а їхня чітка періодичність
порушується. При концентрації гормону 0,5 МО/мл (рис. 2.24,
крива 3) наростання ТМП є різкішим, ніж у нормі, але
продовжується до двох з половиною годин після запліднення (у
нормі до 4 год). Інсулін у концентрації 1,0 МО/мл (рис. 2.24, крива
4) ще різкіше змінює характер динаміки цього показника. Так,
гіперполяризація мембрани через 1 годину і 40 хвилин від початку
ембріогенезу змінювалася різкою деполяризацією, і в змінах рівня
ТМП не виявляється характерної для норми ритміки коливань.
Привертає увагу і той факт, що зі збільшенням
концентрації гормону в середовищі, період тривалості
гіперполяризації клітинної мембрани скорочується. Ступінь
гіперполяризації клітинної мембрани, залежно від кількості
гормону в середовищі, при фіксованому часі можна ілюструвати
таким чином (рис. 2.25). Сімейство кривих одержано шляхом
трансформування результатів, які визначають вплив різних
концентрацій інсуліну на динаміку ТМП зародків в’юна (рис. 2.24),
що розвиваються.
З рис. 2.24 очевидно також, що зі збільшенням концентрації
гормону в середовищі зменшується ступінь періодичності функцій,
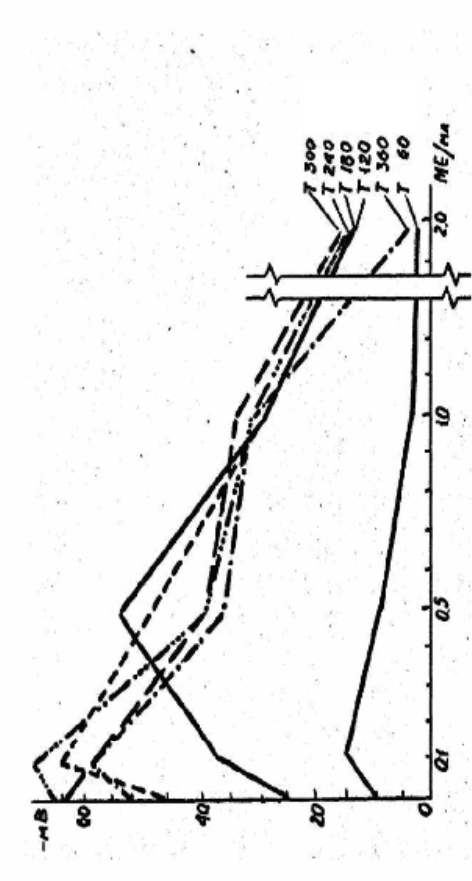
110
Рис. 2.25. Ступінь гіперполяризації клітинних мембран залежно від концентрації інсуліну в середовищі
хв
хв
хв
хв