Pugnaire F.I. Valladares F. Functional Plant Ecology
Подождите немного. Документ загружается.

there is little evidence for this and the depression could easily represent increased PR as in
higher plants. Many mosses show negative NP at higher temperatures, over about 208C
(Gannutz 1970; Longton 1988; Kappen 1989; Noekes and Longton 1989a; Wilson 1990;
Convey 1994, Davey and Rothery 1997). At 08C NP can still be substantial but is always
very low by 58C and ceases at around 78C (Kappen et al. 1989, Pannewitz et al. 2005).
Data on the photosynthetic response to WC for Antarctic mosses are not extensive but it
seems unlikely that they will differ greatly from temperate mosses. Most data are for species in
the northern, maritime Antarctic (Davey 1997b) but little difference was found between these
plants and others studied in temperate or northern tundra areas (Fowbert 1996). At Cape
Hallett (728 S) maximal WC for B. subrotundifolium and B. pseudotriquetrum were around
750% and 600% d.wt., respectively; there was a depression in NP at WC above that optimal
for NP (about 400% and 300%, respectively. Thallus water contents for half maximal NP for
the two species were 150% and 120%. These values for B. pseudotriquetrum fall in the mid-
range of those found for the same species in wet and dry sites at Windmill Islands (668 17
0
S,
Robinson et al. 2000). In their study on Grimmia. antarctici, B. pseudotriquetrum, and
C. purpureus, Robinson et al. (2000) found correlations between water loss rate under
standard conditions, maximal water content, WC for half maximal photosynthetic efficiency,
desiccation tolerance, and water status of the habitat. Samples and species from the drier
habitat lost water less rapidly, had a lower WCmax, and remained photosynthetically active
to lower WC than those from wetter sites.
A strong relationship is reported between moss species distribution and water flow for
several species (Schwarz et al. 1992, Okitsu et al. 2004, Robinson et al. 2003), and Kappen et al.
(1989) found photosynthetic differences between mesic and xeric ecodemes of Schistidium
(Grimmia) antarctici Card.in the Windmill Islands. However, more detailed investigations
are needed because the moss species appear to vary in this characteristic as well. At Granite
Harbour, C. purpureus occupies the wetter sites, the exact opposite as that found for Windmill
Islands. Additionally, drying rates, water contents, and NP are commonly expressed on a dry
weight basis and it is an unfortunate fact that a large proportion of the samples can be
inorganic, for example, Pottia heimii samples that came from an apparently clean moss
hummock in the Dry Valleys had an inorganic content of a surprising 66% (unpublished
results). Response to CO
2
concentration has been little investigated for Antarctic mosses but
appears to be similar to that found for other C
3
species with saturation occurring at levels well
above ambient (Pannewitz et al. 2005). In studies at Cape Hallett at 208C, B. pseudotriquetrum
was not CO
2
saturated at 2000 ppm (Figure 13.4) compared with B. subrotundifolium that was
saturated at about 1000 ppm. The actual CO
2
concentration around the mosses is a matter of
debate but very high values, several times normal ambient levels, have been found in Grimmia
antarctici in continental Antarctica, (Tarnawski et al. 1992) and in B. subrotundifolium at Cape
Hallett (Green et al. 2000). The source of the high CO
2
, its seasonal change and its effect on
overall productivity are not yet known. B. subrotundifolium measured at normal (360 ppm) and
2000 ppm CO
2
under identical conditions showed a 60%–80% increase in daily carbon gain so
carbon budgets modeled on normal ambient levels may be in error (Pannewitz et al. 2005).
Lichens
Considerably more work has been done on Antarctic lichens than the mosses (Kappen 1993a,
2000). However, as they are poikilohydric the lichen and moss photosynthetic responses are
similar in form although differing in detail (see Longton 1988a). PPFD required for satur-
ation can be very high, often around 1000 mmol m
2
s
1
, and light compensation values
are dependent on the thallus temperature, for example: saturation at 1300 mmol m
2
s
1
above 18C and compensation from 5 mmol m
2
s
1
at 28Cto128mmol m
2
s
1
at 208C
for Leptogium puberulum Hue (Figure 13.5 and see Schlensog et al. 1997a). Sun and shade
Francisco Pugnaire/Functional Plant Ecology 7488_C013 Final Proof page 400 16.4.2007 2:34pm Compositor Name: BMani
400 Functional Plant Ecology

forms of Usnea sphacelata R. Br. differed little in the form of their photosynthetic responses
to PPFD at identical temperatures although the sun form had considerably larger NP at
higher temperatures (Kappen 1983). Topt tends to be lower and more consistent than for
mosses and values around 58C158C are common (Longton 1988a, Kappen 1993a).
In mosses, Topt declines with fall in PPFD below saturation (Figure 13.6) (Schroeter et al. 1995,
Thallus temperature (°C)
−5 0 5 10152025
LCP
(µmol m
−2
s
−1
)
0
100
200
300
400
500
600
Leptogium puberulum
Parmelia saxatilis
Umbilicaria aprina
Usnea antarctica
FIGURE 13.5 The typical rise in light compensation point with temperature for four lichens measured in
the field in Antarctica. The most continental species, Umbilicaria aprina, shows the least change. (Data
from Schlensog, M., Schroeter, B., Sancho, L.G., Pintado, A., and Kappen, L., Secc. Biol. 93, 107, 1997a
[Leptogium puberulum]; Sancho, L.G., Parmelia, A., Valladares, F., Schroeter, B., and Schlensog, M., Bibl.
Lichenol. 67, 197, 1997 [Parmelia saxatilis], Schroeter, B., Kappen, L., and Moldaenke, C., Lichenologist,23,
253, 1991 [Usnea antarctica], and Schroeter, B., Grundlagen der Stoffproduktion von Kryptogamen unter
besonderer Beru
¨
cksichtigung der Flechten—eine Synopse—Habilitationsschrift der Mathematisch—
Naturwissenschaftliche Fakulta
¨
t der Christian—Albrechts—Universita
¨
t zu Kiel, 1997 [U. aprina].)
PPFD (µmol m
−2
s
−1
)
0 250 500 750 1000 1250 1500
Topt
(°C)
−5
0
5
10
15
20
25
Leptogium puberulum
Parmelia saxatilis
Umbilicaria nylanderiana
Usnea antarctica
FIGURE 13.6 The typical change in Topt (optimal temperature for net photosynthesis) with increase
in irradiance (PPFD in mmol Photons m
2
s
1
) for four lichens measured in the field in Antarctica.
Sources as for Figure 13.5 except Umbilicaria nylanderiana. (From Sancho, L.G., Pintado, A.,
Valladares, F., Schroeter, B., and Schlensog, M., Bibl. Lichenol., 67, 197, 1997.)
Francisco Pugnaire/Functional Plant Ecology 7488_C013 Final Proof page 401 16.4.2007 2:34pm Compositor Name: BMani
Plant Life in Antarctica 401
Sancho et al. 1997). In complete contrast to mosses, positive NP has been found to
temperatures as low as 178C for U. aprina Nyl. measured in the field (Schroeter et al.
1994) although this ability is not confined to Antarctic lichens, for example: 248C for
Cladonia alcornis (Lightf.) Rabh. measured in the laboratory (Lange 1965).
Fewer studies of the effect of WC on photosynthesis exist but those that do show the
characteristic low WC range (200%–400% maximal WC) compared with mosses and often
with depressed NP at high WC due to increased CO
2
diffusion resistances (Kappen 1985b,
Harrisson et al. 1989, Kappen and Breuer 1991, Schroeter 1991). Lichens in the Antarctic
normally occupy much drier habitats than mosses and are not found in wet areas unless on
the drier tops of mosses. Response to CO
2
concentration seems to have not been measured but
would be expected to be similar to that of temperate lichens that is, not saturated at ambient
CO
2
levels. The increased ambient CO
2
reported around mosses would not be expected
around lichens because of their elevation and lack of substantial organic substrates below
them (Tarnawski et al. 1992).
MAXIMAL RATES OF PHOTOSYNTHESIS
Longton (1988a) gives an extensive list of maximal NP and Topt for a wide range of polar
bryophytes and lichens although only a few are from the areas covered in this article. Kappen
(1988, 1993a) gives additional, similar data for lichens. The brevity of both lists indicates the
lack of knowledge that exists at present. Considerable variation in rates certainly exists for
lichens, from 0:08 mg CO
2
g
1
h
1
for Rhizoplaca melanophthalma (Ram.) Leuck and Poelt,
to 0:8mgCO
2
g
1
h
1
for U. aprina. Although it is occasionally suggested that the abun-
dance of particular lichen species may be related to photosynthetic performance we do not
really yet have the data to be certain. Lechowicz (1982) has analyzed the relationship between
several photosynthetic parameters and latitude for the Northern Hemisphere. From his
analysis maximal NP are certainly substantially lower at latitudes greater than 658 N and
these far-northern values are similar to those found in the Antarctic. Kappen (1988) states
that maximal rates of NP are lower for lichens in continental Antarctic than in the maritime
zone. No similar analysis exists for mosses but, in general, the rates found seem to be close to
the higher rates reported elsewhere (1---2 mg CO
2
g:d:wt:
1
h
1
, Rastorfer 1972, Longton
1988a, Kappen et al. 1989) and no depression is obvious.
DIEL AND LONG-TERM PHOTOSYNTHETIC PERFORMANCE
Continuous measurement of photosynthetic performance over a day, or several days, has only
recently become common in Antarctica and a few extensive studies have been made in the
northern maritime (e.g., Kappen et al. 1991, Schroeter 1991, Schroeter et al. 1991, Sancho
et al. 1997 for lichens, Collins 1977 for mosses). A more common approach to obtain an
estimate of seasonal production is to log microclimate parameters and to interpolate plant
performance from photosynthetic response to PPFD and temperature (e.g., Kappen et al.
1991, Schroeter 1991, Schroeter et al. 1995). The present situation is probably a reflection of
the difficulties of working in Antarctica with large and expensive equipment. Kappen et al.
(1991) obtained eight diurnal records for U. sphacelata near Casey Station and recorded
positive photosynthesis through the entire light period of the day even though temperatures
were mostly below 08C. They were able to demonstrate relatively good agreement between
their model, based on PPFD and temperature, and in situ rates of photosynthesis. Sancho
et al. (1997) obtained diurnal courses for three cosmopolitan lichen species, Parmelia saxatilis
(L.) Ach., Pseudophebe pubescens (L.) M. Choisy, and Umbilicaria nylanderiana (Zahlbr.)
H. Magn., over three months of the summer, 1995, in the maritime Antarctic (Livingston
Island) and found activity only for 182 h. Typical examples of diel patterns for two lichens
Francisco Pugnaire/Functional Plant Ecology 7488_C013 Final Proof page 402 16.4.2007 2:34pm Compositor Name: BMani
402 Functional Plant Ecology
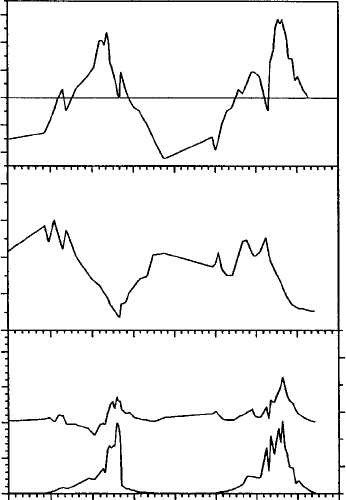
(P. saxatilis and U. aprina) and one moss (B. subrotundifolium) are given in Figure 13.7,
Figure 13.8, and Figure 13.9. The two examples from continental Antarctica (Figure 13.8 and
Figure 13.9) show positive NP 24 h a day, whereas the maritime example (P. saxatilis, Figure
13.7) had negative NP overnight and the daily carbon balance was often negative as, also, was
the cumulative balance calculated over 3 months (Sancho et al. 1997).
Schroeter et al. (1991) demonstrated that basal chlorophyll a fluorescence could be used
to monitor photosynthetic activity of Usnea antarctica Du Rietz. Photosynthetic carbon
fixation could be estimated using microclimate records of temperature and PPFD (Schroeter
1991, Schroeter et al., 1997a,b). Schroeter et al. (1992) demonstrated that the photosynthetic
activity of the crustose lichen, B. frigida (Darb.) Dodge at Granite Harbour could easily be
monitored with a chlorophyll a fluorescence system and this has now become a common
investigative method. Concurrent measurements of CO
2
exchange and chlorophyll a fluores-
cence revealed complex relationships between ETR (relative electron transport rate calculated
from the fluorescence signal) and photosynthetic rate for lichens, and simpler but still not
linear relationships for mosses (Green et al. 1998). Schroeter et al. (1997c) measured fluores-
cence activity of B. frigida over several days at Granite Harbour. The studies revealed the
extremely erratic nature of thallus moistening, which depended on small-scale topography,
proximity to snow, degree of snow melt, and snow fall (Figure 13.10). Even though air
0
0
500
1000
1500
2000
0
0
1
2
3
100
200
300
400
−2
−1
612180
Local time (Feb. 8/9, 1995)
Parmelia saxatilis
PPFD (µmol m
−2
s
−1
)
WC (% d.wt.)
TT (°C)
NP (µmol CO
2
kg
−1
s
−1
)
612180
0
4
8
FIGURE 13.7 Diel pattern of net photosynthesis (upper panel, mmol CO
2
kg
1
s
1
), water content
(middle panel in % d.wt.) and thallus temperature and irradiance (lower panel, PPFD in
mmol Photons m
2
s
1
) for Parmelia saxatilis on February 8 and 9, 1995 measured at Livingston Island.
Because this is a maritime site there is a brief period of darkness each day leading to a period of negative
NP. Measurements were made in the field at a natural growth site using a porometer system and the
lichen was rewetted on the 8th by rainfall. (From Sancho, L.G., Pintado, A., Valladares, F., Schroeter, B.,
and Schlensog, M., Bib. Lichenol, 67, 197, 1997.)
Francisco Pugnaire/Functional Plant Ecology 7488_C013 Final Proof page 403 16.4.2007 2:34pm Compositor Name: BMani
Plant Life in Antarctica 403
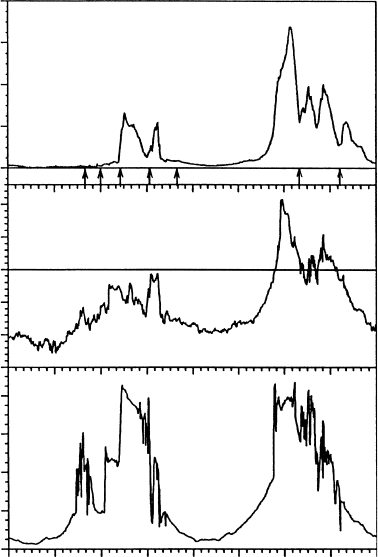
temperatures remained between 7.58C and 12.08C lichen thallus temperatures were
much higher, often around 158C higher, even when wet. Thus, most of the photosynthetic
activity actually occurred at positive temperatures, but was detectable down to 88C, and no
saturation was evident at 1500 mmol m
2
s
1
. These results, which were made in November
at 778 S, indicate that the growing season could be much longer than previously thought, even
at these high latitudes. The introduction of imaging systems for chlorophyll a fluorescence has
allowed even finer analysis of the responses to water and drying (Barta
´
k et al. 2005).
A feature of all the diurnal studies is the elevation of thallus temperatures above ambient
air temperatures because of the absorbed irradiation. Kappen (1993a) noted that thallus
temperatures of wet lichens were often about 88C–108C at many maritime and continental
sites as a result of shelter and sun exposure, although B. frigida at Granite Harbour could
reach 158C (Schroeter et al. 1997c, Kappen et al. 1998b). Monitoring of the photosynthetic
activity of P. heimii on the Canada Glacier flush (Taylor Valley) showed that, in late
0
0
−15
−10
−5
0
5
10
0.0
0.5
1.0
1.5
2.0
Umbilicaria aprina
400
800
1200
1600
61218
Local time (Nov. 19/20, 1994)
PPFD (µmol m
−2
s
−1
)
TT (°C)
0 6 12 18 0
NP (µmol CO
2
m
−1
s
−1
)
FIGURE 13.8 Diel pattern of net photosynthesis (upper panel, mmol CO
2
m
2
s
1
), thallus temperature
(middle panel, 8C), and irradiance (lower panel, PPFD in mmol Photons m
2
s
1
) for Umbilicaria aprina
on November 19 and 20, 1994 at Botany Bay, Granite Harbour. The lichen was rewetted at the points
marked with arrows in the upper panel. Although this is early in the season at a continental site (778 S)
the irradiance reaches high values each day and there is no period of complete darkness. Hence positive
NP can occur through the full 24 h and also, on the 19th at subzero thallus temperatures. (Data from
Schroeter, B., Grundlagen der Stoffproduktion von Kryptogamen unter besonderer Beru
¨
cksichtigung
der Flechten—eine Synopse—Habilitationsschrift der Mathematisch—Naturwissenschaftliche Fakulta
¨
t
der Christian—Albrechts—Universita
¨
t zu Kiel, 1997.)
Francisco Pugnaire/Functional Plant Ecology 7488_C013 Final Proof page 404 16.4.2007 2:34pm Compositor Name: BMani
404 Functional Plant Ecology
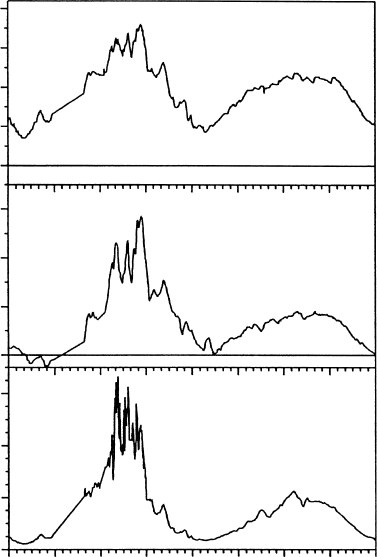
December, the plant was continuously active and did not face subzero temperatures. The wet
plants were buffered from the colder air by ice formation at zero degrees (Figure 13.12).
NUTRITION EFFECTS
The role of nutrients in the abundance and diversity of Antarctic plants is not well known
and requires more detailed studies. Elemental analyses for mosses (Smith 1996) show consider-
able variation depending on proximity to birds and species. Total nitrogen ranged from a low
0.55%–0.77% for P. alpestre to over 3% for Drepanocladus (Sanionia) uncinatus (Hedw.)
Loesk. Rastorfer (1972) found 2.95%, 2.75%, 1.50%, and 2.35% for Calliergidium austrostra-
mineum, Drepanocladus uncinatus, Polytrichum strictum Brid. (syn. P. alpestre), and Pohlia
nutans (Hedw.) Lindb., respectively. These values would not seem to be limiting and are not
likely to be the major control on production. Lichens, in contrast, can have very low
nitrogen contents, for example: 0.36% and 0.83% for U. sphacelata and Usnea aurantiaca-
atra (Jacq.) Bory, respectively (Kappen 1985b). Lichens with green algal photobionts seem
to almost always have below 1% total nitrogen (Green et al. 1980) and fruticose species are
the lowest (Green et al. 1997). However, chlorophyll content, which can be related to
0
0
0
4
8
12
0.0
0.2
0.4
0.6
0.8
400
800
1200
61218
Local time (h) (Jan. 19/20, 1992)
PPFD (µmol m
−2
s
−1
)
TT (°C)
0 6 12 18 0
NP (mg CO
2
g Chl
−1
h
−1
)
FIGURE 13.9 Diel pattern of net photosynthesis (upper panel mg CO
2
Chl
1
h
1
), thallus temperature
(middle panel, 8C), and irradiance (lower panel, PPFD in mmol Photons m
2
s
1
) for moist Bryum
subrotundifolium on January 19 and 20, 1992 at Botany Bay, Granite Harbour. This is just after mid-
summer and NP is positive through the entire period. Thallus temperatures are almost always positive and
are linked to the irradiance. (From Schroeter, B., Grundlagen der Stoffproduktion von Kryptogamen
unter besonderer Beru
¨
cksichtigung der Flechten—eine Synopse—Habilitationsschrift der Mathematisch—
Naturwissenschaftliche Fakulta
¨
t der Christian—Albrechts—Universita
¨
t zu Kiel, 1997.)
Francisco Pugnaire/Functional Plant Ecology 7488_C013 Final Proof page 405 16.4.2007 2:34pm Compositor Name: BMani
Plant Life in Antarctica 405

nitrogen content (Green et al. 1997), is much lower in Antarctic than in temperate species
(Sancho et al. 1997). In essence, the values are little different to those found for temperate
lichens and mosses.
Nutrient supply, especially nitrogen and phosphorous, is strongly influenced by the
presence of birds and other animals. Leishman and Wild (2001) showed that soil nutrient
were enriched near to sea bird nests and declined rapidly with increasing distance. The
number of lichen species and mean lichen species abundance increased with soil nutrient
levels, with total P having a stronger influence than total N. In contrast, moss diversity and
abundance showed little correlation with soil nutrient levels but was positively correlated with
soil water content. Although nutrient deficiency has rarely been shown (Kappen and Schroeter
PPFD (µmol m
−2
s
−1
) Temperature (°C) ETR (∆F/F
m'
⫻ PPFD)
16
0
500
1000
1500
2000
−15
−10
−5
0
5
10
15
0
200
400
600
17 18 19 20 21
November 1994
22 23 24 25 26
FIGURE 13.10 Diel pattern of relative electron transport rate through Photosystem II, ETR, (upper
panel, mmol e
m
2
s
1
), thallus and air temperatures (middle panel, 8C), and irradiance (lower panel,
PPFD in mmol Photons m
2
s
1
) for a Buellia frigida thallus over a 10 day period, November 16–25, 1994,
at Botany Bay, Granite Harbour. Although air temperatures remain around 108C through the whole
period the thallus temperature is positive at times of high irradiance and several degrees above air
temperature during the night because of heat storage in the rock surface. ETR depends on both irradiance
and moistening of the thalli. Initially the lichen received water from melting snow on the rock but, as
the snow patch retreated away from the lichen, it was active only after snow falls (indicated by the stars in
the upper panel). ETR activity is least or zero in the late morning when the lichen has dried out, highest in the
afternoon, after wetting by melt, and continues overnight at subzero temperatures—the so-called reverse
diel pattern (see text). (From Schroeter, B., Grundlagen der Stoffproduktion von Kryptogamen
unter besonderer Beru
¨
cksichtigung der Flechten—eine Synopse—Habilitationsschrift der Mathematisch—
Naturwissenschaftliche Fakulta
¨
t der Christian—Albrechts—Universita
¨
t zu Kiel, 1997.)
Francisco Pugnaire/Functional Plant Ecology 7488_C013 Final Proof page 406 16.4.2007 2:34pm Compositor Name: BMani
406 Functional Plant Ecology
2002), addition of nutrients causes increased photosynthetic rates, ETR (relative electron
transport rate from chlorophyll fluorescence), and chlorophyll content (Smith 1993, Wasley
et al. 2006). Although it is normally claimed that this indicates nutrient limitation, it may be
that the plants are not carbon limited and the enhancement may represent utilization of storage
materials over several years. Longer-term increases in nutrient supply lead to changes in
community structure and biodiversity rather than just increased growth. This can be clearly
seen for mosses in the Ross Sea region. At low nutrient sites such as Taylor Valley, the moss
patches contain only two main species. At a more nutrient rich site, Granite Harbour or
Edmonson Point, four or five species can be common. Finally, where there is an overwhelming
presence of birds, Cape Hallett or Beaufort Island, then only B. subrotundifolium is found.
Overall, the major determinant for the occurrence of mosses, in particular, and lichens is the
availability of liquid water. Extra water may not enhance photosynthesis as much as when
nutrients are also present (Wasley et al. 2006). Finally, there are also suggestions that salt can
control distributions close to the coast (Broady 1989).
SURVIVING ANTARCTICA’S EXTREMES
There are few articles, and possibly even fewer grant applications, written that do not mention
some extreme aspects of Antarctica, usually the climate, and the accepted fact that the
organisms show adaptations to the stresses that they face. Of course the Antarctic climate
is extreme to us, but is it extreme to the organisms that survive there, and do they show special
adaptations; these are important questions in our understanding of potential responses to
change.
DESICCATION
All Antarctic terrestrial biota face a long period of cold, darkness, and desiccation over the
winter. Conditions are particularly extreme in continental Antarctica (Table 13.1) and there
can be little doubt that this limits the vegetation to mainly species of lichens and bryophytes.
Both groups are poikilohydric, meaning that their thallus hydration tends to equilibration
with the water status of the environment (Green and Lange 1994). Water uptake tends, with
the exception of the endohydric mosses like Polytrichum, to occur over the entire surface.
Typically, these groups also show exceptional resistance to desiccation and can survive very
low water contents for long periods. Such resistance is, however, not inevitable and some
lichens, inhabitants of consistently moist rainforest, are very sensitive to desiccation (Green
et al. 1991). The Antarctic liverwort M. berteroana (see earlier Section ‘‘Liverworts’’) is also
very sensitive (Davey 1997a).
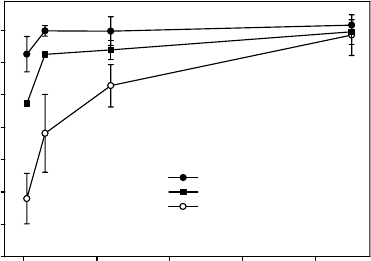
Schlensog et al. (2004) studied the ability of lichens and mosses to recover from desicca-
tion after an Antarctic winter at Granite Harbour (778 S). They found that, although all
species showed recovery starting within minutes as measured by chlorophyll fluorescence, the
lichen U. aprina, from open rock surfaces, reached full photosynthetic activity in just over an
hour, whereas the mosses, B. subrotundifolium and Hennediella heimii from running water
sites, took almost 24 h (Figure 13.11). This difference cannot be generalized to all bryophytes
as various mosses from xeric environments are known to reactivate within minutes (Proctor
2000). Moreover, the lichen P. caesia, characteristic of submerged water channel sides, took
considerably longer than U. aprina to recover (Figure 13.11). The results fit with the concept
from non-Antarctic bryophytes that the tolerance and activation of poikilohydric organisms
can be correlated to their normal active environment. That is, plants from xeric environments
are strongly desiccation-tolerant and activate rapidly, and the opposite occurs for plants from
consistently wet environments.
Francisco Pugnaire/Functional Plant Ecology 7488_C013 Final Proof page 407 16.4.2007 2:34pm Compositor Name: BMani
Plant Life in Antarctica 407
EXTREME COLD
In the dry state, poikilohydric organisms tend to be resistant to environmental extremes. Dry
lichens all survived liquid nitrogen temperatures (1968C, Kappen 1973) and moist thalli of
Antarctic species (Xanthoria candelaria (L.) Th. Fr., R. melanophthalma) fully tolerated slow
or rapid freezing to 1968C (Kappen and Lange 1972). Extended periods of cold and dryness
also had little effect, Alectoria ochroleuca (Hoffm.) Massal recovered totally after 3.5 years at
608C (Larson 1978) and the moss Schistidium antarctici withstood 18 months at 188C
(Kappen et al. 1989). Despite the impressive figures given above it is equally clear that
considerable differences do exist between species, both lichens and mosses, in their abilities
to withstand cold. Liverworts, based on the evidence from M. berteroana, seem to be excluded
from continental Antarctica, because of their lack of cold tolerance, as also are cyanobacterial
lichens (Lange 1965, Schroeter et al. 1994). Lichens also show a range of abilities to photo-
synthesize below 08C (Lange 1965).
There are, however, several reports of damage through subzero temperatures to young
shoot apices in mosses (Longton and Holdgate 1967, Collins 1976) and it does appear that
these highly hydrated shoots are at risk. Generally, however, it must be accepted that the
plants in Antarctica can survive the winter cold of their habitat.
FREEZE–THAW CYCLES
Freeze–thaw cycles can be extremely common in Antarctica with up to 110 in 1 year recorded
in the northern maritime (Longton 1988a). The cycles, although rarely falling to temperatures
more than a few degrees below freezing point, are thought to provide a severe stress to the plants
through intracellular freezing damaging tissue, extracellular freezing disrupting structure,
dehydration by withdrawal of water to external ice and phase changes in membranes leading
to loss of cell contents. Lovelock et al. (1995a,b) studied the effects of freeze=thaw cycles on the
photosynthesis of G. antarctici as monitored using chlorophyll a fluorescence. Every subzero
cycle caused decreased photosynthetic efficiency but recovery at low light was also rapid.
The most thorough analysis has been by Kennedy (1993a) for a northern maritime
species, P. alpestre, an endohydric moss in contrast to the entirely exohydric species of the
Elapsed time after sprayin
g
(h)
0 20 40 60 80
F
v
/F
m
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Bryum subrotundifolium
Umbilicaria aprina
Physcia caesia
FIGURE 13.11 Recovery of F
v
=F
m
(optimal quantum efficiency of Photosystem II measured after
darkening) for specimens that had been rehydrated after collection in a dehydrated state under
snow in early summer. Note the rapid recovery of Umbilicaria aprina (rock surface lichen), slow recovery
of Bryum subrotundifolium (moss from continually wet areas), and intermediate rate for Physcia caesia
(a lichen species that borders intermittent water flows and can be submerged for some time each day).
Francisco Pugnaire/Functional Plant Ecology 7488_C013 Final Proof page 408 16.4.2007 2:34pm Compositor Name: BMani
408 Functional Plant Ecology
continental Antarctic. The species proved to be sensitive to freeze=thaw cycles with reduction
in GP almost directly proportional to the temperature reached and with no recovery at 58C
or below. Almost all damage occurred on the first freeze cycle and plants with lower thallus
water contents were less sensitive. The mechanism of damage was not clarified but could well
have been membrane disruption since increased nitrogen loss from the plants followed the
freezing (Greenfield 1988). Kennedy (1993b) was of the opinion that freeze= thaw cycles could
easily limit species distribution, however, it must be questioned as to how severe this stress
actually is, especially to exohydric mosses. Continual recordings of photosynthetic activity of
P. heimii in Taylor Valley (Pannewitz et al. 2003a) showed that freezing of surrounding water
buffered the moss so that it faced temperatures only a few tenths of a degree below freezing
(Figure 13.12). Actual freezing temperature of the moss initiates about 1.88C (personal
research observations). It appears that, for much of the summer season, mosses in wet areas
are rarely exposed to subzero temperatures and that freeze–thaw events may be confined to
drier environments.
Wetting=drying cycles, also common in the Antarctic, are also thought to be disruptive
and release of carbohydrates has been detected (Davey 1997b, Greenfield 1993, Melick and
Seppelt 1992). Friedmann et al. (1993) estimated that examples of the cryptoendolithic
community in the Ross Desert would pass through an active=inactive cycle (either wet=dry
or freeze=thaw) a minimum of 120–150 days each year with substantial loss of metabolites
(Greenfield 1988). The stresses associated with these cycles contributed to the difference
between a modeled net production of 106 mg C m
2
year
1
and actual growth, estimated
using a variety of techniques, to be 3mgCm
2
year
1
.
Studies on U. aprina, going through a regular daily wetting=drying event near a retreating
snow patch showed no deleterious effects on the lichen (Schroeter et al. 1997c). A fuller
understanding of these transients is still needed before we can better gauge their effects on
2000/2001
16 18 20 22 24 26 28 30 1 3
MOSS temperature (°C)
–5
0
5
10
–10
–5
0
5
December January
AIR temperature (°C)
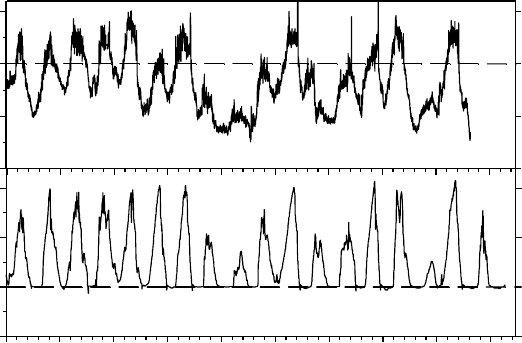
FIGURE 13.12 Uncoupling of thallus temperature (lower panel, 8C) from air temperature (upper panel,
8C) for Pottia heimii measured over an 18 day period in mid-summer at the Canada Glacier flush, Taylor
Valley, 778 S. Note how the air temperature is substantially negative each day, whereas the wet and
active moss never goes subzero. The temperature maxima and minima for air=moss were 4.0=10.0
and 7.0=0.18C, respectively. (Data modified from Pannewitz, S., Green, T.G.A., Scheidegger, C.,
Schlensog, M., and Schroeter, B., Pol. Biol., 26, 545, 2003a.)
Francisco Pugnaire/Functional Plant Ecology 7488_C013 Final Proof page 409 16.4.2007 2:34pm Compositor Name: BMani
Plant Life in Antarctica 409