Ответы по всему курсу Физиологии человека
Подождите немного. Документ загружается.

Закон раздражения Дюбуа-Реймона (аккомодации): раздражающее действие
постоянного тока зависит не только от абсолютной величины силы тока или его
плотности, но и от скорости нарастания тока во времени. При действии медленно
нарастающего раздражителя возбуждение не возникает, так как происходит
приспосабливание возбудимой ткани к действию этого раздражителя, что получило
название аккомодации. Аккомодация обусловлена тем, что при действии медленно
нарастающего раздражителя в мембране возбудимой ткани происходит повышение
критического уровня деполяризации. При снижении скорости нарастания силы
раздражителя до некоторого минимального значения потенциал действия вообще
не возникает. Причина заключается в том, что деполяризация мембраны является
пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению
натриевой проницаемости, и тем самым обусловливающего возникновение
потенциала действия, и медленного, приводящего к инактивации натриевой
проницаемости и как следствие этого - окончанию потенциала действия. При
быстром нарастании стимула повышение натриевой проницаемости успевает
достичь значительной величины прежде, чем наступит инактивация натриевой
проницаемости. При медленном нарастании тока на первый план выступают
процессы инактивации, приводящие к повышению порога или ликвидации
возможности генерировать ПД вообще. Способность к аккомодации различных
структур неодинакова. Наиболее .высокая она у двигательных нервных волокон, а
наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка.
Закон силы-длительности: раздражающее действие постоянного тока зависит не
только от его величины, но и от времени, в течение которого он действует. Чем
больше ток, тем меньше времени он должен действовать для возникновения
возбуждения.
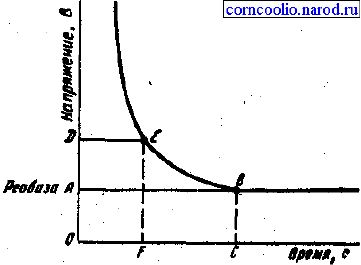
Исследования зависимости силы-длительности показали, что последняя имеет
гиперболический характер (рис. 3). Из этого следует, что ток ниже некоторой
минимальной величины не вызывает возбуждение, как бы длительно он не
действовал, и чем короче импульсы тока, тем меньшую раздражающую способность
они имеют. Причиной такой' зависимости является мембранная емкость. Очень
"короткие" токи просто не успевают разрядить эту емкость до критического уровня
деполяризации. Минимальная величина тока, способная вызвать возбуждение при
неограниченно длительном его действии, называется реобазой. реобазе, и вызывает
возбуждение, называется полезным временем.
Рис.3. Графическое выражение закона силы-длительности.
В связи с тем, что определение этого времени затруднено, было введено понятие
хронаксия - минимальное время, в течение которого ток, равный двум реобазам,
должен действовать на ткань, чтобы вызвать ответную реакцию. Определение
11
хронаксии - хронаксимет-рия - находит применение в клинике. Электрический ток,
приложенный к мышце, проходит через как мышечные, так и нервные волокна и их
окончания, находящиеся в этой мышце. Так как хронаксия нервных волокон
значительно меньше хронаксии мышечных волокон, то при исследовании хронаксии
мышцы практически получают хронаксию нервных волокон. Если нерв поврежден
или произошла гибель соответствующих мотонейронов спинного мозга (это имеет
место при полимиелите и некоторых других заболеваниях), то происходит
перерождение нервных волокон и тогда определяется хронаксия уже мышечных
волокон, которая имеет большую величину, чем нервных волокон.
Закон полярного действия постоянного тока: при замыкании тока возбуждение
возникает под катодом, а при размыкании - под анодом. Прохождение постоянного
электрического тока через нервное или мышечное волокно вызывает изменение
мембранного потенциала покоя. Так, в области приложения к возбудимой ткани
катода положительный потенциал на наружной стороне мембраны уменьшается,
возникает деполяризация, которая быстро достигает критического уровня и
вызывает возбуждение. В области же приложения анода положительный потенциал
на наружной стороне мембраны возрастает, происходит гиперполяризация
мембраны и возбуждение не возникает. Но при этом под анодом критический
уровень деполяризации смещается к уровню потенциала покоя. Поэтому при
размыкании цепи тока гиперполяризация на мембране исчезает и потенциал покоя,
возвращаясь к исходной величине, достигает смещенного критического уровнями
возникает возбуждение.
Закон физиологического электротона: действие постоянного тока на ткань
сопровождается изменением ее возбудимости. При прохождении постоянного тока
через нерв или мышцу порог раздражения под катодом и соседних с ним участках
понижается вследствие деполяризации мембраны - возбудимость повышается. В
области приложения анода происходит повышение порога раздражения, т. е.
снижение возбудимости вследствие гиперполяризации мембраны. Эти изменения
возбудимости под катодом и анодом получили название электротона
(электротоническое изменение возбудимости). Повышение возбудимости под
катодом называется катэлектротоном, а снижение возбудимости под анодом -
анэлектротоном.
При дальнейшем действии постоянного тока первоначальное повышение
возбудимости под катодом сменяется ее понижением, развивается так называемая
катодическая депрессия. Первоначальное же снижение возбудимости под анодом
сменяется ее повышением - анодная экзальтация. При этом в области приложения
катода происходит инактивация натриевых каналов, а в области действия анода
происходит снижение калиевой проницаемости и ослабление исходной инактивации
натриевой проницаемости.
Физиология нервов и нервных волокон.
Нервные волокна (отростки нервных клеток) выполняют специализированную
функцию - проведение нервных импульсов. По морфологическому признаку нервные
волокна делятся на миелиновые (покрытые миелиновой оболочкой) и
безмиелиновые. Нервные волокна формируют нерв или нервный ствол, состоящий
из большого числа нервных волокон, заключенных в общую соединительноткан-ную
оболочку. В состав нерва входят миелиновые и безмиелиновые волокна.
12
Нервные волокна, проводящие возбуждение от рецепторов в ЦНС называются
афферентными, а волокна, проводящие возбуждение от ЦНС к исполнительным
органам, называются эфферентными. Нервы состоят из афферентных и
эфферентных волокон.
Нервное волокно обладает следующими физиологическими свойствами:
возбудимостью, проводимостью, лабильностью.
Проведение возбуждения по нервным волокнам осуществляется по определенным
законам.
Закон двустороннего проведения возбуждения по нервному волокну.
Возбуждение по нервному волокну распространяется в обе стороны от места его
возникновения, т. е. центростремительно и центробежно. Это можно доказать, если
на нервное волокно наложить регистрирующие электроды на некотором расстоянии
друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют
электроды по обе стороны от места раздражения.
Закон анатомической и физиологической целостности нервного волокна.
Проведение возбуждения по нервному волокну возможно лишь в том случае, если
сохранена его анатомическая и физиологическая целостность. Различные факторы,
воздействующие на нервное волокно (наркотические вещества, охлаждение,
перевязка и т. д.) приводят к нарушению физиологической целостности, т. е. к
нарушению механизмов передачи возбуждения. Несмотря на сохранение его
анатомической целостности проведение возбуждения в таких условиях нарушается.
Закон изолированного проведения возбуждения по нервному волокну. В
составе нерва возбуждение по нервному волокну распространяется изолированно, т.
е. не переходя с одного волокна на другое. Изолированное проведение возбуждения
обусловлено тем, что сопротивление жидкости, заполняющей межклеточные
пространства, значительно ниже сопротивления мембраны нервных волокон.
Поэтому основная часть тока, возникающего между возбужденным и
невозбужденным участками нервного волокна, проходит по межклеточным щелям,
не действуя на рядом расположенные нервные волокна.
Изолированное проведение возбуждения имеет важное значение. Нервсодержит
большое количество нервных волокон (чувствительных, двигательных,
вегетативных), которые иннервируют различные по структуре и функциям
эффекторы (клетки, ткани, органы). Если бы возбуждение внутри нерва
распространялось с одного нервного волокна на другое, то нормальное
функционирование органов было бы невозможно.
Нервные волокна по скорости проведения возбуждения, диаметру волок- на,
длительности различных фаз потенциала действия и строению принято
подразделять на три типа: А, В, С. Волокна типа А в свою очередь делятся на
подтипы: А-альфа, А-бета, А-гамма, А-дельта.
Волокна типа А покрыты миелиновой оболочкой. Наиболее толстые из них А -
альфа, они имеют диаметр 12-22 мкм и высокую скорость проведения возбуждения -
70-120 м/с. Эти волокна проводят возбуждение от моторных нервных центров
спинного мозга к скелетным мышцам (двигательные волокна) и от определенных
рецепторов мышц к соответствующим нервным центрам.
13
Три другие группы волокон тина А (бета, гамма, дельта) имеют меньший диаметр от
8 до 1 мкм и меньшую скорость проведения возбуждения от 5 до 70 м/с. Волокна
этих групп относятся преимущественно к чувствительным, проводящим
возбуждение от различных рецепторов (тактильных, температурных, некоторых
болевых, рецепторов внутренних органов) в ЦНС. Исключение составляют лишь
гамма-волокна, значительная часть которых проводит возбуждение от клеток
спинного мозга к интрафузальным мышечным волокнам.
К волокнам типа В относятся миелинизированные преганглио-нарные волокна
вегетативной нервной системы. Их диаметр - 1-3,5 мкм, а скорость проведения
возбуждения - 3-18 м/с.
К волокнам типа С относятся безмиелиновые нервные волокна малого диаметра -
0,5-2,0 мкм. Скорость проведения возбуждения в этих волокнах не более 3 м/с (0,5-
3,0 м/с) . Большинство волокон типа С - это постганглионарные волокна
симпатического отдела вегетативной нервной системы, а также нервные волокна,
которые проводят возбуждение от болевых рецепторов, некоторых терморецепторов
и рецепторов давления.
Механизм проведения возбуждения по нервным волокнам. Механизм
распространения возбуждения у различных нервных волокон неодинаков. По
современным представлениям распространение возбуждения по нервным волокнам
осуществляется на основе ионных механизмов генерации потенциала действия.
При распространении возбуждения по безмиелиновому нервному волокну местные
электрические токи,, которые возникают между его возбужденным участком,
заряженным отрицательно, и невозбужденным, заряженным положительно,
вызывают деполяризацию мембраны до критического уровня с последующей
генерацией ПД в ближайшей точке невозбужденного участка мембраны. Этот
процесс повторяется многократно. На всем протяжении нервного волокна
происходит процесс репродукции нового ПД в каждой точке мембраны волокна.
Такое проведение возбуждения называете и непрерывным.
Наличие у миелиновых волокон оболочки, обладающей высоким электрическим
сопротивлением, а также участков волокна, лишенных оболочки (перехватов
Ранвье) создают условия для качественно нового типа проведения возбуждения по
миелиновым нервным волокнам. Местные электрические токи возникают между
соседними перехватами Ранвье, т. к. мембрана возбужденного перехвата становится
заряженной отрицательно по отношению к поверхности соседнего невозбужденного
перехвата. Эти местные токи деполярязуют мембрану невозбужденного перехвата
до критического уровня и в нем возникает ПД (рис. 4). Следовательно, возбуждение
как бы "перепрыгивает" через участки нервного волокна, покрытые миелином, от
одного перехвата к другому. Такой механизм распространения возбуждения
называется сальтаторным или скачкообразным. Скорость такого способа
проведения возбуждения значительно выше и он более экономичен по сравнению с
непрерывным проведением возбуждения, поскольку в состояние активности
вовлекается не вся мембрана, а только ее небольшие участки в области перехватов.
14
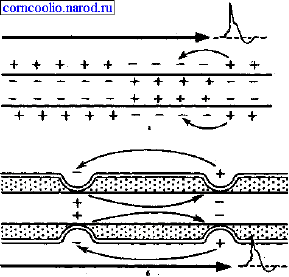
Рис. 4. Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б)
нервных волокнах.
"Перепрыгивание" потенциала действия через участок между перехватами
возможно потому, что амплитуда ПД в 5-6 раз превышает пороговую величину,
необходимую для возбуждения соседнего перехвата. ПД может "перепрыгивать" не
только через один, но и через два межперехватных промежутка. Это явление может
наблюдаться при снижении возбудимости соседнего перехвата под действием
какого-либо фармакологического вещества, например, новокаина, кокаина и др.
Нервные волокна обладают лабильностью - способностью воспроизводить
определенное количество циклов возбуждения в единицу времени в соответствии с
ритмом действующих раздражителей. Мерой лабильности является максимальное
количество циклов возбуждения, которое способно воспроизвести нервное волокно в
единицу времени без трансформации ритма раздражения. Лабильность
определяется длительностью пика потенциала действия, т. е. фазой абсолютной
рефрактерности. Так как длительность абсолютной ре-фрактерности у спайкового
потенциала нервного волокна самая короткая, то лабильность его самая высокая.
Нервное волокно способно воспроизвести до 1000 импульсов в секунду.
Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е.
воздействию повреждающего агента) посредством, например, отравления или
повреждения, то лабильность такого участка резко снижается. Восстановление
исходного состояния нервного волокна после каждого потенциала действия в
поврежденном участке происходит медленно. При действии на этот участок частых
раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и
поэтому проведение импульсов блокируется. Такое состояние пониженной
лабильности было названо Н. Е. Введенским парабиозом. В развитии состояния
парабиоза можно отметить три, последовательно сменяющих друг друга, фазы:
уравнительную, парадоксальную, тормозную.
В уравнительную фазу происходит уравнивание величины ответной реакции на
частые и редкие раздражители. В нормальных условиях функционирования
нервного волокна величина ответной реакции иннервируемых им мышечных
волокон подчиняется закону силы: на редкие раздражители ответная реакция
меньше, а на частые раздражители - больше. При действии парабиотического
агента и при редком ритме раздражении (например, 25 Гц) все импульсы
возбуждения проводятся через парабиотический участок, так как возбудимость
после предыдущего импульса успевает восстановиться. При высоком ритме
раздражении (100 Гц) последующие импульсы могут поступать в тот момент, когда
нервное волокно еще находится в состоянии относительной рефрактерности,
15
вызванной предыдущим потенциалом действия. Поэтому часть импульсов не
проводится. Если проводится только каждое четвертое возбуждение (т. е. 25
импульсов из 100) , то амплитуда ответной реакции становится такой же, как на
редкие раздражители (25 Гц) - происходит уравнивание ответной реакции.
В парадоксальную-фазу происходит дальнейшее снижение лабильности. При этом
на редкие и частые раздражители ответная реакция возникает, но на частые
раздражители она значительно меньше, т. к. частые раздражители еще больше
снижают лабильность, удлиняя фазу абсолютной рефрактерности. Следовательно,
наблюдается парадокс - на редкие раздражители ответная реакция больше, чем на
частые.
В тормозную фазу лабильность снижается до такой степени, что и редкие, и частые
раздражители не вызывают ответной реакции. При этом мембрана нервного волокна
деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается
ее исходное состояние.
Явление парабиоза лежит в основе медикаментозного локального обезболивания.
Влияние анестезирующих веществ также связано с понижением лабильности и
нарушением механизма проведения возбуждения по нервным волокнам.
Парабиоз - явление обратимое. Если парабиотическое вещество действует недолго,
то после прекращения его действия нерв выходит из состояния парабиоза через те
же фазы, но в обратной последовательности.
Механизм развития парабиотического состояния сводится к следующему. При
воздействии на нервное волокно парабиотического фактора нарушается способность
мембраны увеличивать натриевую проницаемость в ответ на раздражение. В
участке альтерации инак-тивация натриевых каналов, вызванная повреждающим
агентом, суммируется с инактивацией, вызываемой нервным импульсом, и
возбудимость снижается настолько, что проведение следующего импульса
блокируется.
Физиология мышц.
У человека различают три вида мышц:
• поперечно-полосатые скелетные мышцы;
• поперечно-полосатая сердечная мышца;
• гладкие мышцы внутренних органов, кожи, сосудов.
Мышцы обладают физическими и физиологическими свойствами. Рассмотрим те
свойства, которые характерны для скилетных мышц.
Физические свойства скелетных мышц.
1. Растяжимость - способность мышцы изменять свою длину под действием
растягивающей ее силы.
16
2. Эластичность - способность мышцы принимать свою первоначальную длину
после прекращения действия растягивающей или деформирующей силы. Живая
мышца обладает малой, но совершенной эластичностью: уже небольшая сила
способна вызвать относительно большое удлинение мышцы, а возвращение ее к
первоначальным размерам является полным. Эти свойства очень важны для
осуществления нормальных функций скелетных мышц.
3. Сила мышцы. Она определяется максимальным грузом, который мышца в
состоянии поднять. Для сравнения силы различных мышц определяют их удельную
силу - максимальный груз, который мышца в состоянии поднять, делят на число
квадратных сантиметров ее физиологического поперечного сечения.
4. Способность мышцы совершать работу. Работа мышцы определяется
произведением величины поднятого груза на высоту подъема. Работа мышцы
постепенно увеличивается с увеличением груза, но до определенного предела,
после которого увеличение груза приводит к уменьшению работы, т. к. снижается
высота подъема груза. Следовательно, максимальная работа мышцей производится
при средних величинах нагрузок (закон средних нагрузок).
Физиологические свойства мышц.
• Возбудимость - способность приходить в состояние возбуждения при действии
раздражителей.
• Проводимость - способность проводить возбуждение.
• Сократимость - способность мышцы изменять свою длину или напряжение в
ответ на действие раздражителя.
• Лабильность - лабильность мышцы равна 200-300 Гц.
При непосредственном раздражении мышцы (прямое раздражение) или
опосредовано через иннервирующий ее двигательный нерв (непрямое раздражение)
одиночным стимулом возникает одиночное мышечное сокращение, в котором
выделяют три фазы:
• латентный период - время от начала действия раздражителя до начала ответной
реакции;
• фаза сокращения (фаза укорочения);
• фаза расслабления.
В естественных условиях к скелетной мышце из ЦНС поступают не одиночные
импульсы, а серия импульсов, следующих друг за другом с определенными
интервалами, на которую мышца отвечает длительным сокращением. Такое
длительное сокращение мышцы, возникающее в ответ на ритмическое раздражение
получило название тетанического сокращения или тетануса. Различают два вида
тетануса: зубчатый и гладкий.
17
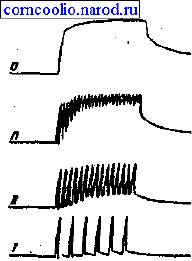
Если каждый последующий импульс возбуждения поступает к мышце в тот период,
когда она находится в фазе укорочения, то возникает гладкий тетанус, а если в
фазу расслабления - зубчатый тетанус (рис. 5).
Рис. 5. Различные виды тетануса при повышении частоты раздражения. I -
одиночные сокращения; II-III - зубчатый тетанус; VI - гладкий (сплошной) тетанус
Амплитуда тетанического сокращения превышает амплитуду одиночного
мышечного сокращения. Исходя из этого Гельмгольц объяснил процесс
тетанического сокращения простой суперпозицией, т. е. простой суммацией
амплитуды одного мышечного сокращения с амплитудой другого. Однако в
дальнейшем было показано, что при тетанусе имеет место не простое сложение
двух механических эффектов, т. к. эта сумма может быть то большей, то меньшей. Н.
Е. Введенский объяснил это явление с точки зрения состояния возбудимости
мышцы, введя понятие об оптимуме и пессимуме частоты раздражения.
Оптимальной называется такая частота раздражения, при которой каждое
последующее раздражение осуществляется в фазу повышенной возбудимости.
Тетанус при этом будет максимальным по амплитуде - оптимальным.
Пессимальной называется такая частота раздражения, при которой каждое
последующее раздражение осуществляется в фазу пониженной возбудимости.
Тетанус при этом будет минимальным по амплитуде - пессимальным.
Режимы мышечных сокращений. Различают изотонический, изометрический и
смешанный режимы сокращения мышц.
При изотоническом сокращении мышцы происходит изменение ее длины, а
напряжение остается постоянным. Такое сокращение происходит в том случае,
когда мышца не перемещает груз. В естественных условиях близкими к
изотоническому типу сокращений являются сокращения мышц языка.
При изометрическом сокращении длина мышечных волокон остается постоянной,
меняется напряжение мышцы. Такое сокращение мышцы можно получить при
попытке поднять непосильный груз.
В целом организме сокращения мышц никогда не бывают чисто изотоническим или
изометрическим, они всегда имеют смешанный характер, т. е. происходит
изменение и длины, и напряжения мышцы. Такой режим сокращения называется
ауксотоническим если преобладает напряжение мышцы, или ауксометрическим
если преобладает укорочение.
18
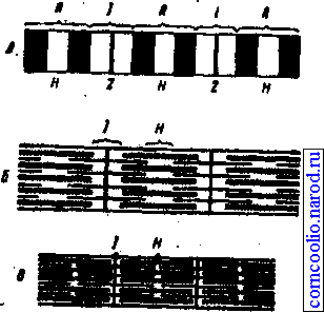
Механизм мышечного сокращения. Мышцы состоят из мышечных волокон,
которые состоят из множества тонких нитей - миофибрилл, расположенных
продольно. Каждая миофибрилла состоит из протофибрилл - нитей сократительных
белков актина и миозина. Перегородки, называемые 2-пластинами, разделяют
миофибриллы и, следовательно, мышечное волокно на участки - саркомеры. В
саркомере наблюдают правильно чередующиеся поперечные светлые и темные
полосы. Эта поперечная исчерченность миофибрилл обусловлена определенным
расположением нитей актина и миозина. В центральной части каждого саркомера
свободно расположены толстые нити миозина. На обоих концах саркомера
находятся тонкие нити актина, прикрепленные к Z-пластинам. Нити миозина
выглядят в световом микроскопе как светлая полоска (Н-зона) в темном диске,
который дает двойное лучепреломление, т. к. содержит нити миозина и актина и
называется анизотропным или А-диском. По обестороны от А-диска находятся
участки, которые содержат только тонкие нити актина и кажутся светлыми, т. к.
они обладают одним лучепреломлением и называются изотропными или j-дисками.
По их середине проходит темная линия - Z-мембрана. Именно благодаря такому
периодическому чередованию светлых и темных дисков сердечная и скелетная
мышцы выглядят исчерченными (поперечно-полосатыми) (рис. 6).
Рис. 6. Электронномикроскопическая картина миофибриллы (схематизировано)(А).
Взаимное расположение толстых (миозиновых) и тонких (актиновых) нитей в
расслабленной (Б) и сокращенной (В) миофибрилле.
В состоянии покоя концы толстых и тонких: нитей лишь незначительно
перекрываются на уровне А-диска. В соответствии с теорией скользящих нитей при
сокращении тонкие актиновые нити скользят вдоль толстых миозиновых нитей,
двигаясь между ними к середине саркомера. Сами актиновые и миозиновые нити
своей длины не изменяют.
Механизм скольжения нитей. Миозиновые нити имеют поперечные мостики
(выступы) с головками, которые отходят от нити биполярно. Актиновая нить состоит
из двух закрученных одна вокруг другой цепочек (подобно скрученным ниткам бус)
молекул актина. На нитях актина расположены молекулы тропонина, а в желобках
между двумя нитями актина лежат нити тропомиозина. Молекулы тропомиозина в
покое располагаются так, что предотвращают прикрепление поперечных мостиков
миозина к актиновым нитям.
Во многих местах участки поверхностной мембраны мышечной клетки углубляются
в виде трубочек внутрь волокна, перпендикулярно его продольной оси, образуя
систему поперечных трубочек (Т-систему). Параллельно миофибриллам и
19
перпендикулярно поперечным трубочкам расположена система продольных
трубочек (альфа-система). Пузырьки на концах этих трубочек - терминальные
цистерны - подходят очень близко к поперечным трубочкам, образуя совместно с
ними так называемые триады. В этих пузырьках сосредоточено основное
количество внутриклеточного кальция.
В состоянии покоя миозиновый мостик заряжен энергией (миозин
фосфорилирован), но он не может соединиться с нитью актина, так как между ними
находится система из нитей тропомиозина и глобул тропонина. При возбуждении
ПД быстро распространяется по мембранам поперечной системы внутрь клетки и
вызывает высвобождение ионов кальция из альфа-системы. С появлением ионов
кальция в присутствии АТФ происходит изменение пространственного положения
тропонина, в результате чего отодвигается нить тропомиозина и открываются
участки актина, присоединяющие ми-озиновые головки. Соединение головки
фосфорилированного миозина с актином приводит к изменению положения мостика
(его "сгибанию"), в результате конформации этой части миозиновой молекулы, и
перемещению нити актина на один шаг (на один "гребок") к середине саркомера.
Затем происходит отсоединение мостика от актина. Ритмические прикрепления и
отсоединения головок миозина позволяют "грести" или тянуть актиновую нить к
середине саркомера.
При отсутствии повторного возбуждения ионы кальция закачиваются кальциевым
насосом из протофибриллярного пространства в систему саркоплазматического
ретикулума. Это приводит к снижению концентрации ионов кальция и
отсоединению его от тропонина. Вследствие чего тропомиозин возвращается на
прежнее место и снова блокирует активные центры актина. Вместе с тем,
происходит фосфорилирование миозина за счет АТФ, который не только заряжает
системы для дальнейшей работы, но и способствует временному разобщению нитей.
Удлинение (расслабление) мышцы после ее сокращения является процессом
пассивным, поскольку актиновые и миозиновые нити легко скользят в обратном
направлении под влиянием сил упругости мышечных волокон и мышцы, а также
силы растяжения мышц антагонистов.
Гладкие мышцы. Гладкие мышцы, формирующие мышечные слои стенок желудка,
кишечника, мочеточников, бронхов, кровеносных сосудов и других полых
внутренних органов, построены из веретенообразных одноядерных мышечных
клеток. Отдельные клетки в гладких мышцах функционально связаны между собой
низкоомны-ми электрическими контактами - нексусами. За счет этих контактов
потенциалы действия и медленные волны деполяризации беспрепятственно
распространяются с одного мышечного волокна на другое. Поэтому несмотря на то,
что двигательные нервные окончания расположены на небольшом числе мышечных
волокон, вследствие беспрепятственного распространения возбуждения с одного
волокна на другое в реакцию вовлекается вся мышца. Следовательно, гладкие
мышцы представляют собой не морфологический, а функциональный синцитий.
Особенностью гладких мышц является их способность осуществлять относительно
медленные движения и длительные тонические сокращения. Медленные, имеющие
ритмический характер, сокращения гладких мышц желудка, кишечника,
мочеточников и других органов обеспечивают перемещение содержимого этих
органов. Длительные тонические сокращения гладких мышц особенно хорошо
выражены в сфинктерах полых органов, которые препятствуют выходу содержимого
этих органов.
20