Наумов А.Д. Двустворчатые моллюски Белого моря. Опыт эколого-фаунистического анализа
Подождите немного. Документ загружается.

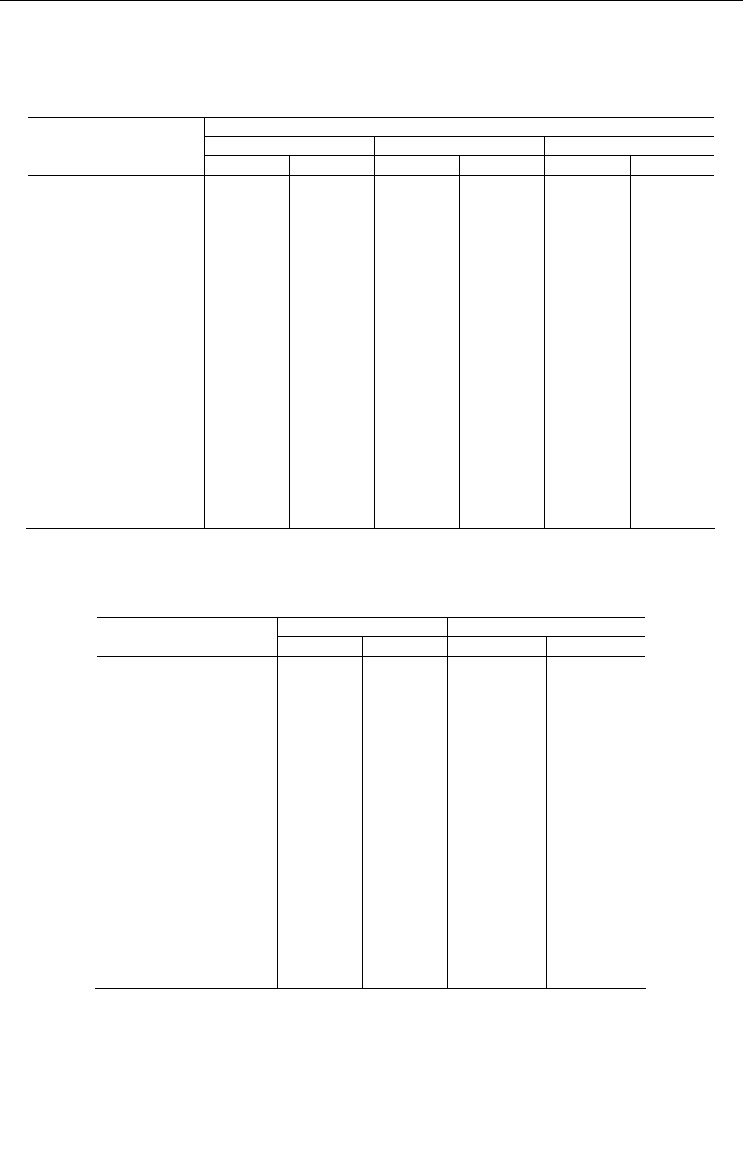
Глава 7. ВЕРТИКАЛЬНОЕ РАСПРЕДЕЛЕНИЕ ДВУСТВОРОК
140
Таблица 21
Вертикальное распределение двустворчатых моллюсков
в кутовой котловине губы Колвицы
Vertical distribution of clams in the head depression of the Kolvitsa Inlet
Глубина, м
0 ÷ 10 20 ÷ 30 30 ÷ 40
Вид
биомасса плотность биомасса плотность биомасса плотность
Serripes groenlandicus 95.800 24
Macoma balthica 0.173 80
Macoma calcarea 154.980 233 21.710 56
Elliptica elliptica 62.400 20 12.400 4
Hiatella spp. 17.800 30 1.464 4
Leionucula bellotii 7.370 18 1.076 6
Clinocardium ciliatum 7.276 4
Nuculana pernula 6.480 8 5.050 8
Musculus laevigatus 3.720 4 3.920 4
Nicania montagui 1.060 4
Thyasira gouldi 0.292 40 0.188 4
Crenella decussata 0.128 16
Yoldia hyperborea 0.100 8 6.010 8
Dacrydium vitreum 0.064 8
Portlandia arctica 67.500 154
Mya truncata 43.830 6
Pandora glacialis 0.920 4
Таблица 22
Двустворчатые моллюски, обнаруженные на порогах губы Колвицы
Bivalves found at the ridges of the Kolvitsa Inlet
Кутовой порог Входной порог
Вид
биомасса плотность биомасса плотность
Mya truncata 183.3 74 15.520 4
Nuculana pernula 2.4 4
Leionucula bellotii 1 4
Macoma calcarea 0.4 4
Thyasira gouldi 0.3 8
Macoma balthica 41.160 48
Clinocardium ciliatum 34.512 24
Chlamys islandica 15.088 16
Elliptica elliptica 1.184 8
Hiatella spp. 0.720 68
Musculus niger 0.664 16
Portlandia arctica 0.228 4
Nicania montagui 0.172 4
Heteranomia squamula 0.148 20
Crenella decussata 0.136 28
Dacrydium vitreum 0.048 4
Фауна губы Лов так же, как и губы Колвицы, представляет собой несколько обед-
ненную фауну Бассейна (Наумов, 1979). Встреченные в ней виды распределены не-
равномерно. Фауна входной котловины разнообразнее кутовой.
По фауне в губе можно выделить два горизонта. Граница между ними в кутовой
котловине проходит на глубинах 7÷10 м, во входной – около 20 м. В верхнем гори-
зонте преобладают бореальные и арктическо-бореальные формы, в нижнем – аркти-
ческие (Наумов, 1979).

СОСТАВ И РАСПРЕДЕЛЕНИЕ ДВУСТВОРОК
141
Анализ распределения доли биомассы двустворок по глубинам позволяет разде-
лить каждый горизонт на два этажа. Верхний этаж верхнего горизонта (0 ÷ 5 м) засе-
лен в основном Mytilus edulis, нижний – Crenella decussata. Плотность поселений
кренеллы достигает почти 300 экз./м
2
. В верхнем этаже нижнего (холодноводного)
горизонта преобладает Macoma calcarea. Наконец, нижний этаж нижнего горизонта
кутовой котловины губы Лов, представляет собой настоящее царство Portlandia
arctica (Наумов, 1979). Фауна этого горизонта в основных чертах соответствует арк-
тическому фаунистическому комплексу больших глубин Бассейна (Зенкевич, 1927;
1963; Дерюгин, 1928; Гурьянова, 1948, 1949). Ведущей формой и здесь так же, как в
Бассейне и в других губах подобного типа, является моллюск Portlandia arctica. В
кутовой котловине губы Лов биомасса портландии превышает биомассу всех осталь-
ных моллюсков, вместе взятых, почти в пять раз (Наумов, 1979).
Недостаток опубликованных данных не позволяет описать вертикальное распре-
деление двустворчатых моллюсков в Палкиной губе сколько-нибудь подробно, одна-
ко, судя по данным А. Н. Голикова с соавторами (1982), на литорали этой губы до-
минирует Mytilus edulis, в верхней сублиторали – Musculus laevigatus и Tridonta bore-
alis, а еще глубже – Macoma calcarea. Как по его материалам, так и по данным Бент-
БоМ, на глубинах свыше 40 м и в этой губе доминирует Portlandia arctica (табл. 23).
Биогеографический состав населения любой акватории определяется термопатией
видов различного генезиса и термическим режимом водоема. Поскольку среди ков-
шовых губ в гидрологическом отношении лучше всего изучена Колвица, то просле-
дить закономерности вертикального распределения фауны удобнее всего на ее при-
мере. Из рассмотренных выше трех типов вод, характерных для этой губы, два пер-
вых по температуре не различаются сколько-нибудь существенно. Учитывая резкое
сокращение пресного стока зимой, воды второго типа можно, как и первого, считать
водами летнего происхождения. Биогеографический состав донного населения на той
или иной глубине определяется соотношением летних и зимних вод в период гидро-
логического лета.
Вертикальное членение бентали на участки, заселенные видами различной био-
географической природы, удобно проводить по соотношению их биомасс. В губе
Колвица выделяются три таких участка. Бореальный, занимающий глубины до 1.5 м,
расположен в районе влияния поверхностного слоя воды, гомогенного за счет ветро-
вого перемешивания, т. е. ядра вод летнего происхождения. Бореально-арктический
участок ограничен глубинами 1.5÷40 м и лежит в области, характеризующейся мак-
симальными градиентами (Наумов и др., 1986, б), т. е. в области смешения вод зим-
него и летнего происхождения. Наконец, арктический участок целиком находится в
пределах ядра глубинных вод.
Таблица 23
Двустворчатые моллюски, обнаруженные в котловине Палкиной губы
Clams found in the depression of the Palkina Inlet
Вид Биомасса Плотность
Portlandia arctica 85.974 714
Clinocardium ciliatum 11.350 6
Macoma calcarea 4.924 20
Mya truncata 5.760 4
Lyonsia arenosa 0.148 8
Pandora glacialis 0.056 4
Leionucula bellotii 0.028 8

Глава 7. ВЕРТИКАЛЬНОЕ РАСПРЕДЕЛЕНИЕ ДВУСТВОРОК
142
Как видно, условия на верхнем и глубинном участках можно характеризовать
термическим режимом одного конкретного типа вод, чего нельзя сказать про средин-
ный. По аналогии с биогеографическими областями, каждой из которых соответству-
ет определенный тип водной массы, мы выделяем вертикальные биогеографические
зоны, коих в Колвице оказывается всего две – арктическая и бореальная. Граница
между ними проходит на глубине 6÷15 м (Наумов и др., 1986, б). Именно на эти глу-
бины приходится наибольший градиент температур и, как показано в разделе «Гид-
рологические особенности», здесь проходит верхняя граница распространения вод
зимнего происхождения.
В зависимости от доминирования той или иной биогеографической группировки в
бентосе, каждый из полученных участков можно разделить на верхний и нижний
горизонты. Верхний горизонт бореальной зоны, характеризующийся преобладанием
атлантических форм, можно отнести к Норвежской провинции Атлантической боре-
альной области. Что же касается нижнего горизонта арктической зоны, то он, в силу
обедненности видового состава фауны, может быть отнесен к Беломорскому округу
Беломорско-Шпицбергенской провинции Арктической области (Федяков, 1986).
Два оставшихся горизонта, где доминируют бореально-арктические виды, отли-
чаются друг от друга не только соотношением субдоминантных биогеографических
группировок, но и характерными для них биоценозами. Так, в нижнем горизонте бо-
реальной зоны отмечены биоценозы мелких полихет, Balanus crenatus, Serripes
groenlandicus и Муа truncata, а в верхнем арктической – Macoma calcarea. Интересно,
что в Кандалакшском желобе все эти биоценозы занимают нижний горизонт боре-
альной зоны (Семенова и др., 1995, а, б). Верхний же горизонт арктической зоны в
Бассейне заселен биоценозами Nuculana pernula, Gersemia fruticosa, Pectinaria
hyperborea и др., совершенно выпадающими в Колвице. Интересно отметить, что
сообщество, переходное к высокоарктическому, в Бассейне в качестве субдоминант-
ной формы включает в себя Nuculana pernula, которая как в Колвице, так и в других
ковшовых и лагунных губах, заменяется другим детритофагом – Macoma calcarea.
Аналогичная вертикальная зональность описана и для Лов губы (Наумов, 1979).
Сходство этой зональности в губах Долгой, Лов и Колвице отмечается и В. В. Федя-
ковым (1986). Нечто аналогичное описано и для Бабьего моря (Гурвич, 1934) с той
лишь разницей, что на его максимальных глубинах имеется зона сероводородного
заражения. Таким образом, мнение о том, что ковшовые и лагунные губы представ-
ляют собой природные модели, в миниатюре отражающие основные фаунистические
и гидрологические особенности Белого моря в целом (Наумов, 1979), получает до-
полнительное подтверждение.
Вероятно, обнаружение в ковшовых губах биоценозов высокоарктических видов
на относительно небольших глубинах привело к появлению расхожего мнения о
смещении зон в этих водоемах. На самом деле, как справедливо отмечал К. М. Дерю-
гин (1928), никакого смещения зон не происходит: изменяются лишь диапазоны глу-
бин, на которых они расположены. Более того, в губе Колвица сохраняются даже
пропорции этих диапазонов на всех горизонтах (Наумов и др., 1986, б).
И все же нельзя утверждать, что арктические сообщества ковшовых губ полно-
стью идентичны биоценозу, заселяющему максимальные глубины Бассейна. На
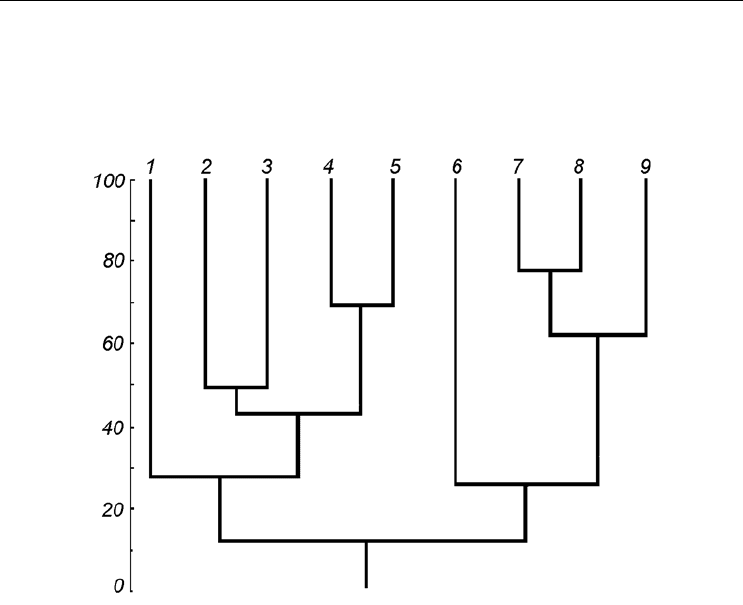
рис. 78 представлена кладограмма сходства таких сообществ из различных районов
Белого моря. Хорошо видно, что биоценозы открытых участков Бассейна образуют
один кластер, а биоценозы ковшовых губ – другой. Интересно, что к последнему
примыкает и высокоарктическое сообщество, обнаруженное в Двинском заливе, а в
Кандалакшском несколько особняком стоит аналогичный биоценоз из района архи-
пелага Средние луды. Эти две точки расположены на окраинах арктического сообще-
ства, встреченного в Бассейне. Факт этот требует дополнительного изучения.
СОСТАВ И РАСПРЕДЕЛЕНИЕ ДВУСТВОРОК
143
Рис. 78. Кладограмма сходства биоценозов с доминированием Portlandia arctica в различных
местах Белого моря, построенная методом взвешенных средних.
По оси ординат – сходство по Жаккару (Jaccard, 1912), вычисленное по долям биомасс видов. 1 – Двин-
ский залив, 2 – Палкина губа, 3 – кутовая котловина губы Колвицы, 4 – входная котловина губы Колвицы,
5 – Долгая губа, 6 – Кандалакшский залив, точка возле архипелага Средние луды, 7 – Великая салма, 8 –
Кандалакшский залив между м. Турьим и м. Шараповым
, 9 –центральная часть Бассейна.
По: Naumov, Fedyakov, 2000, a, с изменениями.
Fig. 78. Jaccard’s (1912) similarity of communities with Portlandia arctica dominating calculated by
species biomass fraction using weighted means method.
Y-axis – similarity. 1 – the Dvina Bay, 2 – the Palkina Inlet, 3 – the head depression of the Kolvitsa Inlet, 4 – the
mouth depression of the Kolvitsa Inlet, 5 – the Dolgaya Inlet, 6 – Kandalaksha Bay near the Sredniye Ludy Archi-
pelago, 7 – the Velikaya Salma Strait, 8 – the Kandalaksha Bay between Toury Cape and Sharapov Cape, 9 –
central part of the Basin. Modified from Naumov, Fedyakov, 2000, a.
É·‚‡ 8
éÅôÄü ïÄêÄäíÖêàëíàäÄ îÄìçõ
ÑÇìëíÇéêóÄíõï åéããûëäéÇ ÅÖãéÉé åéêü
Фаунистические комплексы беломорских двустворчатых моллюсков
Несмотря на то, что биогеография морских организмов все еще практически не
формализована и остается одной из наименее разработанных в теоретическом отно-
шении дисциплин среди всего комплекса направлений морской биологии, биогео-
графический анализ локальных фаун широко применяется на практике и дает непло-
хие результаты. Подобному анализу подвергались и беломорские моллюски (Федя-
ков
, 1980, 1986; Наумов, Федяков, 1987, а).
Номенклатура биогеографических характеристик видов весьма разнообразна, и ее
нельзя назвать устоявшейся. В одни и те же термины разные авторы вкладывают за-
частую совершенно разный смысл. Одни дают биогеографическую характеристику
вида на основании его современного ареала, в то время как другие имеют в виду ме-
сто его происхождения,
причем как те, так и другие далеко не всегда четко оговари-
вают, что именно они подразумевают. Об этом писал еще С. М. Герценштейн (1885),
и тех пор мало что изменилось. В проводимом ниже биогеографическом анализе
фауны беломорских двустворчатых моллюсков нас не будет интересовать происхож-
дение видов, поэтому мы не будем делить их
на атлантические и тихоокеанские, на
амфибореальные и циркумполярные. Нас будет интересовать только обобщенная
характеристика термопатии, поэтому на основании современных ареалов, которые и
являются в некотором смысле ее отражением, мы будем делить беломорских дву-
створчатых моллюсков всего на три категории: бореальные, бореально-арктические и
арктические виды. По сути дела, наиболее корректно
было бы проводить биогеогра-
фический анализ на основе биомасс соответствующих видов, однако по сложившейся
традиции подобные исследования проводят на уровне видовых списков, поэтому в
настоящей работе для сравнимости результатов с данными других авторов биогео-
графический анализ проводится также на основе видового состава.
Среди беломорских двустворчатых моллюсков по числу видов преобладают боре
-
ально-арктические формы: их 79%. Бореальные представлены 18%, а арктические –
3%; при этом на разных глубинах пропорциональное соотношение видов различной
биогеографической принадлежности может отличаться. Учитывая это обстоятельство
В. В. Федяков (1980), анализируя сходство видового состава двустворчатых моллю-
сков в Белом море, в основном в Кандалакшском желобе, на разных глубинах, при-
меняя показатель сходства Жаккара
и индекс различия Престона, делит этот водоем
по вертикали на основные 2 зоны. Первая из них в среднем по морю занимает глуби-
ны от –2 до 2 м, т. е. отвечает литорали и самой верхней части сублиторали, где еще
заметно сказывается влияние волнового воздействия. В. В. Федяков указывает, что в
этой зоне встречается 6
видов, причем самым характерным из них, не выходящим за

ФАУНИСТИЧЕСКИЕ КОМПЛЕКСЫ
145
ее пределы, оказывается Mya arenaria. Вторая зона распространяется в среднем от
10 м до максимальных глубин. Между этими зонами расположен промежуточный
пояс, который характеризуется сменой биогеографического состава двустворчатых
моллюсков, резким градиентом изменения видового состава с увеличением глубины
и не имеет характерных, только ему свойственных видов. Вторую зону В. В. Федяков
разделяет на
три горизонта.
Верхний из них занимает в среднем область между 10 и 80 м. В нем В. В. Федяков
отмечает 36 видов, из которых 21 отмечен только здесь. Характерные виды этой об-
ласти – Nuculana minuta, Crenella decussata, Chlamys islandica, Thracia myopsis,
Serripes groenlandicus и Arctica islandica. Далее следует средний горизонт с глубина-
ми в среднем от 80 до 240 м. В. В. Федяков
считает, что его население – обедненная
фауна верхнего горизонта. Характерные виды здесь – Nuculana pernula, Yoldia
hyperborea и Musculus corrugatus.
Нижний горизонт по В. В. Федякову занимает глубины от 240 до 343 м и населен
обедненной фауной среднего. Здесь встречено 7 видов, наиболее характерный из ко-
торых – Portlandia arctica.
Соглашаясь в целом с описанным выше вертикальным членением Белого моря
и с
тем, на каких глубинах проходят границы между зонами, мы склонны предложить
иные трактовки вертикальной зональности рассматриваемого водоема.
На наш взгляд, в Белом море имеется не две, а три фаунистические зоны, между
которыми располагаются промежуточные области. Верхняя зона, выделенная
В. В. Федяковым, вне всякого сомнения, действительно отвечает литорали и
самым
верхним отделам сублиторали. Следующий за ней пояс, несомненно, промежуточ-
ный, переходный ко второй фаунистической зоне. При низкой прозрачности бело-
морской воды можно сказать, что он отвечает в целом области фитали, так как глуб-
же него опускаются только багрянки, да и те, как правило, не достигают 20-метровой
изобаты (Голиков и др
., 1985, а, б; Луканин и др., 1995, а). На этих глубинах, дейст-
вительно, число как бореальных, так и бореально-арктических видов достигает 50%,
т. е. происходит смена биогеографического состава фауны, что хорошо видно на
рис. 2 цитируемой работы (Федяков, 1980) (следует иметь в виду, что на этом рисун-
ке при публикации были перепутаны обозначения биогеографических
группировок).
Далее идет верхняя сублитораль со своим набором характерных для нее форм, и ее
можно считать второй самостоятельной фаунистической зоной. За ней следует вто-
рой промежуточный пояс, приуроченный к нижней сублиторали. Не следует сму-
щаться тем, что по диапазону глубин он превышает сто метров. На большей части
акватории эта область принадлежит
достаточно крутому свалу
1
(рис. 9), что делает ее
не столь протяженной, а в Двинском заливе, где наблюдается пологий уклон на соот-
ветствующих глубинах, фаунистический комплекс третьей, наиболее глубоководной
зоны поднимается несколько выше. В пределах этого второго промежуточного пояса
так же, как и в первом, происходит смена биогеографического состава фауны, и в
следующей фаунистической, наиболее
глубоководной, зоне по числу видов начинают
преобладать эндемики Арктики и высокобореально-арктические формы. Эта зона
отвечает псевдобатиали по терминологии, предложенной А. П. Андрияшевым (1974).
Мысль о фаунистической самостоятельности комплекса донных организмов, оби-
тающих в псевдобатиали, высказывалась нами и раньше на основе анализа зависимо-
сти числа видов от солености (Бергер и др., 1985).
1
«Крутизна» свала составляет в среднем десятые доли градуса, что, однако, на порядок
превышает уклон дна выше и ниже этой области.

Глава 8. ОБЩАЯ ХАРАКТЕРИСТИКА ФАУНЫ
146
Полученные результаты подтверждают мнение В. В. Федякова (1986) о том, что
мелководные районы Белого моря по составу фауны принадлежат к Атлантической
бореальной области, а глубоководные – к Арктической.
Итак, литораль населяют в основном бореальные формы, верхнюю сублитораль –
бореально-арктические, в псевдобатиаль – арктические. В промежуточных поясах
происходит смена преобладания группировок. Сходная вертикальная зональность
была
получена и на основе анализа зависимости числа видов бентоса от глубины
(Naumov, 2001). Все три фаунистические группировки заселяли Белое море различ-
ными способами в результате различных фаунистических процессов, причем боре-
альные и бореально-арктические формы пришли с запада, а арктические – с востока
(см. ниже).
Имеющийся в нашем распоряжении материал позволяет разделить
двустворчатых
моллюсков Белого моря по характеру их батиметрического распределения на не-
сколько групп.
Литоральный фаунистический комплекс включает в себя 6 видов, из которых 2 –
Mya arenaria и Turtonia minuta – не обнаружены за пределами литоральной фауни-
стической зоны, Macoma balthica достигает первого промежуточного пояса, а
Musculus discors, Musculus laevigatus и Mytilus edulis обнаружены и в сублиторальной
фаунистической зоне.
Наиболее богат
видами сублиторальный фаунистический комплекс. В его состав
входит 26 видов, из которых Yoldiella nana, Chlamys islandica, Lyonsia arenosa,
Thyasira equalis, Axinopsida orbiculata, Panomya arctica, Montacuta maltzani и
Lyonsiella abyssicola не встречены за пределами сублиторальной фаунистической
зоны; Crenella decussata, Heteranomia aculeata, Tridonta borealis, Clinocardium
ciliatum, Serripes groenlandicus, Arctica islandica и Mya truncata обнаруживаются так-
же и в первом переходном поясе; Yoldia hyperborea, Modiolus modiolus, Elliptica
elliptica, Pandora glacialis и Thracia myopsis
встречены также во втором промежуточ-
ном поясе, а Nuculana minuta, Musculus niger, Heteranomia squamula, Nicania
montagui, Hiatella spp. и Macoma calcarea отмечены в обоих промежуточных поясах.
В псевдобатиальный фаунистический комплекс входит 6 видов, из которых ни
один не приурочен к нему строго. Portlandia arctica выходит во второй промежуточ-
ный пояс, а в ковшовых губах поднимается даже еще выше,
правда, практически не
выходя за пределы области круглогодичных отрицательных температур. Nuculana
pernula, Dacrydium vitreum и Musculus corrugatus захватывают и сублиторальную
фаунистическую зону, а Leionucula bellotii и Thyasira gouldi выходят даже в первый
промежуточный пояс.
Распространение двустворчатых моллюсков в Белом море
Помимо вертикального распределения, важную роль в фаунистическом анализе
играет географическое распространение видов. Для исследования этого вопроса ак-
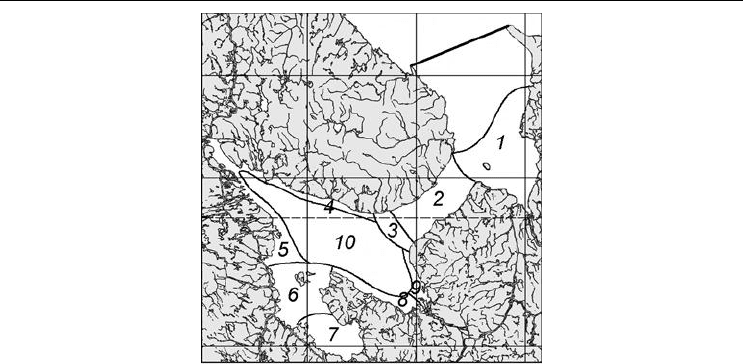
ватория Белого моря была разбита на районы, установленные на основании анализа
распространения видов моллюсков различной биогеографической принадлежности
(Федяков, 1986). Это районирование было дополнено гидрологическими границами в
Горле, Онежском и Двинском заливах, принятыми на основе
областей фронтов, от-
меченных А. Н. Пантюлиным (2003) (рис. 79). Так было выделено 10 исходных рай-
онов. Затем в каждом из них подсчитывалось количество встреч всех видов, за ис-
ключением крайне редких (Yoldiella nana, Panomya arctica, Thyasira equalis,
Axinopsida orbiculata, Montacuta maltzani и Lyonsiella abyssicola). На основании полу-
ченных таким образом данных была рассчитана матрица парных коэффициентов
РАСПРОСТРАНЕНИЕ ДВУСТВОРОК В БЕЛОМ МОРЕ
147
Рис. 79. Разбиение Белого моря на районы для последующего анализа.
Fig. 79. Preliminary division of the White Sea into regions for the further analysis.
корреляции Браве-Пирсона между всеми районами. Районы объединялись, если кор-
реляция между ними превышала 0.5. Это значение при данном числе видов досто-
верно на 95% уровне вероятности. В результате было получено 6 районов, отличаю-
щихся друг от друга (рис. 80, А).
Полученные результаты лишь отчасти совпадают с теми выводами, к которым
пришел В. В. Федяков
(1986). Северная часть моря и северная часть Онежского зали-
ва так же, как и у него, оказались весьма сходными по составу встреченных в них
двустворчатых моллюсков, однако южная часть Онежского залива отличается от них
вполне существенно. Достаточно однородными по числу встреч двустворок оказа-
лись мелководные области вдоль Терского, Кандалакшского, и Карельского
берегов.
В самостоятельный район выделилось прибрежье Двинского залива, и, наконец, со-
вершенно особняком стоит Кандалакшский желоб, что полностью совпадает с дан-
ными В. В. Федякова.
На основе проведенного районирования было проанализировано распространение
видов двустворчатых моллюсков в Белом море. Для этого на том же материале по
встречаемости отдельных видов в выделенных районах
(см. выше) была рассчитана
матрица корреляций между видами. Считалось, что они имеют в Белом море одина-
ковый тип распространения, если корреляция между ними составляла более 0.65, что
при полученном числе районов достоверно на 95% уровне вероятности.
Следует помнить, что почти все моллюски могут быть встречены практически в
любой части моря, и выделенные
типы их распространения (в дальнейшем мы не-
сколько условно будем называть эти типы локальными ареалами) характеризуются
сходством по преимущественной встречаемости. Если тот или иной вид не встречен
в каком-либо районе совершенно, то это оговаривается. В первую очередь это отно-
сится к Горлу, где не встречено значительное количество видов. Этот вопрос
будет
рассмотрен особо. Всего было получено 5 основных типов локальных ареалов дву-
створчатых моллюсков в Белом море.
Моллюски, имеющие первый тип локальных ареалов (рис. 80, Б), встречены
преимущественно в Горле, на мелководьях вдоль Терского, Кандалакшского и
Карельского берегов, а также в северной части Онежского залива. Можно выделить
два варианта этого типа ареалов.
К первому варианту принадлежит распространение
Musculus laevigatus, Mya truncata и Hiatella spp. Эти виды встречаются относительно
одинаково часто на всей указанной площади, однако тяготеют в основном к Канда-
лакшскому заливу и северной части Онежского. По второму варианту распростране-

Глава 8. ОБЩАЯ ХАРАКТЕРИСТИКА ФАУНЫ
148
ны Nuculana minuta, Crenella decussata, Modiolus modiolus, Chlamys islandica и
Heteranomia squamula. Они приурочены в основном к северной части Онежского за-
лива и редки в Кандалакшском. Из всех этих видов ни один, кроме Modiolus modiolus
и Hiatella spp., ни разу не встречен в Бассейне; Nuculana minuta не отмечен в Мезен-
ском заливе, а Chlamys islandica вообще не обнаружен ни в
Двинском, ни в Мезен-
ском заливах.
Моллюски, обладающие вторым типом локальных ареалов (рис. 80, В),
встречены преимущественно на мелководьях вдоль Терского, Кандалакшского и Ка-
рельского берегов, а также в северной части Онежского и в Двинском заливе. Этот
тип ареалов распадается на три варианта. К первому из них следует отнести распро-
странение
Musculus discors, Tridonta borealis и Lyonsia arenosa. Все эти виды тяготе-
ют в основном к Кандалакшскому заливу и северной части Онежского. Второй вари-
ант составляют Macoma calcarea и Mya arenaria, относительно редкие в Онежском
заливе и приуроченные в первую очередь к Кандалакшскому. Третий вариант пред-
ставляет собой распространение Macoma balthica. Этот вид чаще всего встречается в
Кандалакшском
и Двинском заливах. Ни один из этих видов, судя по доступному
автору материалу, ни разу не встречен в Горле; Lyonsia arenosa, Macoma balthica и
Mya arenaria не отмечены в южной части Онежского залива; Musculus discors,
Tridonta borealis, Macoma balthica и Mya arenaria не обнаружены в Бассейне, а по-
следний из этих видов – и в Мезенском заливе.
Третьим типом
локальных ареалов (рис. 80, Г) обладает 5 видов двуствор-
чатых моллюсков. В этом типе, который охватывает все мелководные области, кроме
Горла, можно выделить два варианта. К первому из них следует отнести Heteranomia
aculeata, Thracia myopsis и Arctica islandica, которые отмечены в Мезенском заливе и
чаще всего встречаются в северной части Онежского. Ко второму варианту принад-
лежат Clinocardium ciliatum и Serripes groenlandicus. Оба эти вида не отмечены в Ме-
зенском заливе, причем первый из них чаще всего встречается в Кандалакшском за-
ливе и в северной части Онежского, а второй – в Кандалакшском и Онежском зали-
вах. Ни в Горле, ни в Бассейне ни один из этих видов на доступном нам
материале не
отмечен.
Четвертый тип распространения двустворчатых моллюсков в Белом море
(рис. 80, Д) имеет три варианта. Этот тип ареала захватывает практически все море,
за исключением Горла, Мезенского залива и южной части Онежского. К его первому,
более мелководному варианту принадлежат Leionucula bellotii, Musculus niger,
Pandora glacialis и Thyasira gouldi. Все они чаще всего встречаются в
Кандалакшском
заливе, а два последних из них – еще и в северной части Онежского. В южной части
этого залива совершенно не встречен только Musculus niger, а остальные представле-
ны единичными находками. К относительно глубоководному варианту можно отне-
сти распространение Nuculana pernula, Dacridium vitreum и Musculus corrugatus. Пер-
вые два из них чаще встречаются в Кандалакшском заливе, а
последний – в северной
части Онежского. Этот же вид на нашем материале ни разу не встречен в Двинском
заливе. Особняком стоит Portlandia arctica: этот вид встречается только в Канда-
лакшском желобе, ковшовых губах с холодноводным режимом и локальных депрес-
сиях кута Кандалакшского залива с низкими температурами. Ни один из этих видов,
за
исключением Musculus niger, ни разу не встречен в Горле.
Пятый тип локальных ареалов (рис. 80, Е) охватывает всю акваторию Бело-
го моря. В этом типе распространения моллюсков тоже есть три варианта. К первому
принадлежит ареал Yoldia hyperborea, вид, не отмеченный в Горле и Мезенском зали-
ве, а ко второму – Mytilus edulis. Этот мелководный и литоральный
вид распростра-
нен вдоль всего побережья, а на открытой акватории встречен только в южной части
Онежского залива. Наконец, Elliptica elliptica и Nicania montagui распространены
практически равномерно по всему морю.

РАСПРОСТРАНЕНИЕ ДВУСТВОРОК В БЕЛОМ МОРЕ
149
Рис. 80. Районы Белого моря, сходные по преобладающим видам двустворчатых моллюсков
(А), и основные типы выделенных локальных ареалов.
Районы, близкие по фауне двустворчатых моллюсков, заштрихованы одинаково. Б – локальный ареал
первого типа, В – локальный ареал второго типа, Г – локальный ареал третьего типа, Д – локальный ареал
четвертого типа, Е – локальный ареал пятого типа.
Fig. 80. The White Sea regions similar by predominating bivalves (A) and main types of local
distribution areas distinguished.
Regions similar by bivalve fauna have the same hatch. Б – local distribution area of the first type, В – local distri-
bution area of the second type, Г – local distribution area of the third type, Д – local distribution area of the fourth
type, Е – local distribution area of the fifth type.