Lewin Benjamin (ed.) Genes IX
Подождите немного. Документ загружается.


tz9
ralourord aq]
lp
salquassv
snlelpddv
lpspg
aql
0I'tz
JZrs SurspaJJur
Jql SurMolloJ ^q
peullJp
^lpnrur
se.t,l sluJle
Jo
serres eqJ
'eserJrudlod
y1r1g
.{q
parnolr
leql
xaldruor
e
plmq
ot JJp]o
pJurJJp
e uI
lle
ol
sJolJPJ uorldrrJsupJl
aql salnbar uorlerlrul
'
uorl0ulsuPll salPrlru
r
1r
'xalduor
aql ol spurq
11
aseraufilod
VNU
uaq4
.
'VNq
uo uor6er
pelrelord
aq1
1o
q16ua1
eq1 6uLpualxa lepro
pauqap
e
uL xaldLuor eql
ol
purq
stolre; uoqdursupll laqlg
.
'u0tlPtltuL
uL dals
lslg
aql sr
xoq
VIVI
oql ol
0IIll;o
6urpurg
r
ralouord
oql
lP
salquassv
snlelPddv
lPseB
aql
'(srolerrtDv
qtlM pelelJossv
eJV sesel^tJJv
'/'0€
uortJJS aas)
uortdrJ)supJt 01
Jorrd urleru
-orqr
Jo
eJntf,nJls eq1 AJrporu
leql
saxaldruor
utalo.rd ur osle
punoJ
are strgyl a>lrl-euolslq
eqt lelnJlued
u1
'qrIgt
uI sP
IIaM
se saxaldtuol
rJqlo
ur
punoJ
aq Leru sIIdVJ
aqt
Jo
aruos
'sIIdVJ
reqto
uJeMlaq
suorlleJJlur
asr,Lured ur
pasn
osle Jrp
sploJ
JuotsrH
'VNC
,ItW (II{J
Jo
suorlJprJlur rgnads
-JJuJnbasuou
Jqt ro;
alqrsuodsJJ
Jq ^prrr JJnl
-JnJlS
e
q)ns
ireruelJo
Juolsrq e Surlqruasar
JJnlJnrts
p
roJ
srspq Jqt ruJoJ
f,eu
39tl4y1
pue
'falrng 'y
uaqdel5
1o
Asalnor
oloqd
'spurq
dgf
ereqM
enoorb
iuoileu
aql
suaprM
leql
xoq
vlvL
eql
le
pueq
P sMoqs
lurooilPls
aql 0l
0t-
urorJ
VNq
ql$
dg1
Jo
ernllnlls
1e1s&ror
aql
,
;i"."-:.rj
j
ZVTTJVI
'sdVJ
raqlo
qt1.u
raqtaSol
('raurepg
JuolsrH
Jq1
Jo
uollezlueSJo
'L'62
uoll
-lJS
ees
lurleuoJqJ rrlor{re>lna ur
vNC
spurq
teqt
xalduroJ Jrseq
Jr{l-JarueDo euolslq
aql
Jo laurr>l
Jr{t rxJoJ
tH
pup
€H
sJuolsrH)
'uoll
-JeJelur
eql
JoJ 3sn seuolsrq
]pq]
(ploJ
Juolsrq
eql)
Juoru
Jrues Jqt Sursn
rarutpoJeteq e urJoJ
Laqt
pue
'7H
pue
€H
seuotslq
;o
sSolouoq
(ruersrp)
aq ol readde
ZgIIdvI
pue
67rrgy1
?e1
-nrrlred
ur
lsJuotsrq elqueseJ
s{VJ JtuoS
'vNq
ol
putq
o1
OII{J
roJ rJpro ur
dgl
Jo
eJeJJns Surpurq-ygq
Jql ruoJJ
pareldsrp
eq
tsnru
0€ZIIdvJ
Jo
ulptuop
Ieurural-N
rql
ivNC
or
purq
ol
dgJ;o
f,rrlrqe
Jql
loJluo)
ot
0€ZIIdvJ
suolle
[rtnunu reln
-rJloru
slqJ
'VNCI
ur anoot8
roulru Jql
Jo
JJeJ
-Jns
eq1 sJrurtu
'0€zIIdYJJo
uleluop
PuIruJel-N
Jql ur sJrT
qJIqM
'alts
Eutpurq eql
Jo
JrntJnrls
eq1
'peJ
uI
'JreJrns
Sutputq-yge
eAPJuoJ aql
sardnlo
tl
Jrer{M
AgJ
01 spulq
0€ZII{VJ
roDeJ
aqt
'xaldruor
uratord
JeJJ B se
(II{J ulqll6
'YNo
qllt't
slJpluo)
rr;nads-aruanbas
saleru
leql
JolJpJ
uorldrnsuerl
1eraua8,{.po
aqr sl
dgJ'luloduels
aql
puo,{aq
ruea:tsdn JrqupJ
spualxJ
osle
pue
'0I-
01
57-
ruorl
uor8a; aql
sDalord
dlreln8ar
TalJ,lroq
'uortJeJJ
uorlplllul
aq1 uI
xaldruot
OIISJ
rq1
1o
Surpurg'SZ-
ot
Lf-
rnoq8upuatxa
I11eld,{1
'xoq
yJVJ
eqt
te
xllaq
elqnop rql
Journl
I-stratord
lJit^utVNq101
491
paryrnd
;o
Surpurg
'uor8ar
slql
ut stJptuoJ
vN(-ulel
-ord
;o
,{.1rsuap
qg1q
p
saleJJf,
'anoor8
roleru
Jql ul Surpurq
suralord
JJqlo
qlIM pJulqruoJ
'arroorS
Jourur
Jql ur
dgJ
Jo
aruasard
aql
'aqlrJM
antltsod
e
Lq
paresuaduro)
sI
pue
'VN(
Jo
urnt
e
Io
prlql
-auo
lnoqe
yo
Surpur.ttun
ol spuodseJJoJ
xoq
VJyJ
rqt
le
Surpuaq
aqJ
'YNC
reeuq
uo alqls
-sod
aq
plnoM
ueql uolleDosse
JasolJ
e ruJoJ 01
aserarur{1od
VNU
puP
sJolJPJ
uo4dtrlsuerl aqt
sMolle
1l
'xoq
YIVI
aql
Jo
rpIS reqllr
uo
vN(
;o
uorlenue8ro
prleds
aql Sufueqr
r(g
'suotler
-gdrur
puollJunJ
Ipralas
sel{ alnDnJls
sII{J
'ralourold
eq1
Io
,(ruatrtyya aql
qlIM
pJlPIerJoJ
sI
pue
'xoq
yJVI
aqt
Jo
eJuanbas
Dexa
aqt
qtpn,{ren
upJ
pueq
aql
Jo
luaxa
JqJ'parrelurBur
sr Suuted
JSeq JSneJJq
VNC
Jo
spu€ns
aqt ateredas
L1e
-nlJe
1ou
saop
lnq'aJnl)nrls
aql
Jo
uouPluJoJep
e sr srqJ
'y
6<
sl
anoorS Joulur
aqt aruanbas
eql
Jo
reluJJ aql
lP 1nq
'Y
S-
Jo
qlplM
Iensn
s1r seq arroorS
rourru
aql
'aruanbes
aql
Jo
puJ
qrpJ
te
lxoq
yJVI
aqr
yo
dq
8
Jqt
ol
pJlJlJlsrr
sr uoruolslp
aq;
'anoor8
Joulur Jql
SuIuapIM
'anoorS
;oleur aq1
pJeMot
spuJq
xoq
VJVJ
JqJ
't.i'+.;t
jH{ti}{:i
ur
palerlsnlll se
'.08-.{qy1trq
aqt
spueq
pue
a'roor8
Joulur
eq1 01 spulq
dgJ
-uol
tr
aJeqM
vNq
aql
Jo
qled
eql seuruJJlJp
osle
1I
'vNC
aql
luerro
ol
qIIcJ
qtrM pup
sserelu
-.{1od
ylqg
Jql
qlrM
sDeJJtur
ureuop
IpurruJJt-)
arlJ
'rJtuJf,
elrtJp
aseraru.{1od
Jqt olul
pauasul
sl
grldJJo
,,n8ur1,,
pate8uola
uy'adersa
raloruord
ol uollerlrul
alruoqe
ruoJJ r{Jlr,t.rS
Jql
sJJuJnllur
pue
vNu
Jo
lrxJ
Jql
qtr^^
seJeJretur
srql
tpqt
alqrssod
sl
lr
lstrxe
\/NU
JJeqM etrs aql
.reau aru.{zua
Jq1 s]rpluo)
gu41
ruo.4 uoqqrJ
Jurz
IPurruJJl-N
Ue
'|.ii"Tlt
..i
..
.i.:
ur l.llerrteruaqrs
peteJtsnllr
sy
'aruLzua
aril
qtrM
tJeJJlur
JotJpJ
JI{t
Jo
sureuop JJJqI
teqt
suoqs JserJur^lod
VNd
qtl,to
gII{I
Jo
JJntf,nrts
lersrfur
aq1
'aru.,{zua
aql;o 3ur
-purq
Jr{]
}o
rll{euolrrJrrp
aql to; alqrsuodsar
sr
1l leql
os
'aseraur,{lod
yNU
Lq
pazruSorJr
uJnt
ut sI
teql
aJelJns
aqt apu,ord
Leru
gngl
'uor8ar
iIUg
eql ur ratoruord
Jql
qtrm
stf eluoJ JrJrJ
-ads-aruanbas
saleru.{1enpe
gItgl
;o
Solouroq
Jql
'eJpqJre
uI
'iIUg
Jqt
pelleJ
uorSar
e ur xoq
VJYI
Jqt
Jo
ueartsdn
anoor8
roleu Jqt
slJel
-uoJ
pue
'xoq
vJvI
Jql
Jo
uesJlsuuop
anoo.rS
Jourur
aql ur sl)e1uoJ
sJ>lptu
1I
'vNC
Jo
J)eJ
auo 3uo1e slJptuoJ
Surpualxa
agJ
01
luatefpe
spulq
gIIdJ
'lSpour
srql spuelxJ
'::
,.
-:i:
,j:1:::ii:i
ul
uMoqs
erntJnrts
1e1sLn
JqJ'spuprts
VNO
o,l,,rl
'Aelrng ';
ueqdal5
1o
fisalnor
so1oq6
'eldtnd
pup pel
s!
gllll pue
'onlq
st
691'nno11a[
pue
uaerb
elp
VN6
Jo
spupl]s
orur1 aql
.VN6
jo
alpJ
luaq
aq1
6uo1e spu!q
gIIII
leql
Moqs
VN0-dBL
-gIIlIJo
xalduroc fireutel
aq11o sMan
oMI
i;.irt
:l-.:rirli
i
slaluPq ul
puP
sraloulold
aql ot
pre8ar
qtrM
petuarJo
z{lerrrlaruru.dse
pue
yN(
qtlu pelpnosse
[1asoo1
sdeqrad
'xoq
yJVI
Jqt
Jo
rrreJrlsumop
punoq
sI
gIIdJ
leql
stsa8
-3ns
srql
'0I+
ot
0I-
ruorJ
'turoduels
eql
Jo
^1r
-ur)rl
aqt ur
pueJls
aleldrual aql
1o
uor8ar aqt
Jo
uorDalord
1e4red
san6
gu41
;o
uoplppv
'0€ZII{VJ
aqr
f,q
pJsne)
sr
leql
uorssa.rdar aql
Sutnatlar
{q
aAf
elenrlJp Leru
yu41
'urea4sdn
JequeJ Surpualxa uorSar
e
tralo:d
01 elqe
seruoJJq
OIIJJ
'xaldruoJ
Jqt suroI
yII{J
uJqM
('turodtrels
Jqt
te
aJuenbes
UUI
aql sazruSo
-rrr
osle
qtrgl)
'xoQ
VJYJ
rqt spurq (IIdJ
uaqM
peleruur
sr raloruord
e 01
luetulr-To,
'J;BIS
Jlel
e
te
palerodJof,ur
sr aseraruLlod
yNU
'para^ol
sl
VN(
yo ql8ua1
Surseanur ue
'xaldruor
aql
surof rolrey
lg1
qrea
sV
'].1
i'ir!.j
]H{1*l:j
ur
pJzrr
-eruruns
Iapou
aql slsJ83ns xalduror
qrea
Lq
paDalo.rd
suor8ar
VN(I
eql
Jo
3urlurrdtoo{
'VN61
ot
punoq
aseraruLlod
VNU
Jo
stuJJl ur
pue
sJolJPI
snorrP^ Jql
Jo
seJnDnJls
IPl
-s,{n
aql .{q
palea,,rar
suorlJpJelur Jql
Jo
srurJl
ur
lrelJp
aJorrr
ur slue^J aq1 surJap ueJ JM MoN
'VNq1
qtl^^ peterf,ossp
xaldruor
uralord Jrp
Jo
'srollPJ
uorl0ulsuerl
qlrM
uorleDossP
1o
eruanbes
palopro
ue Aq
11
aserauflod
VNU
loJ sloloru
-otd
1e
salqLuesse xalduor uorlpqrur
uV
.r.i.r:il
SijIi:,\:.j
7Z
UtldVHl ZZ9

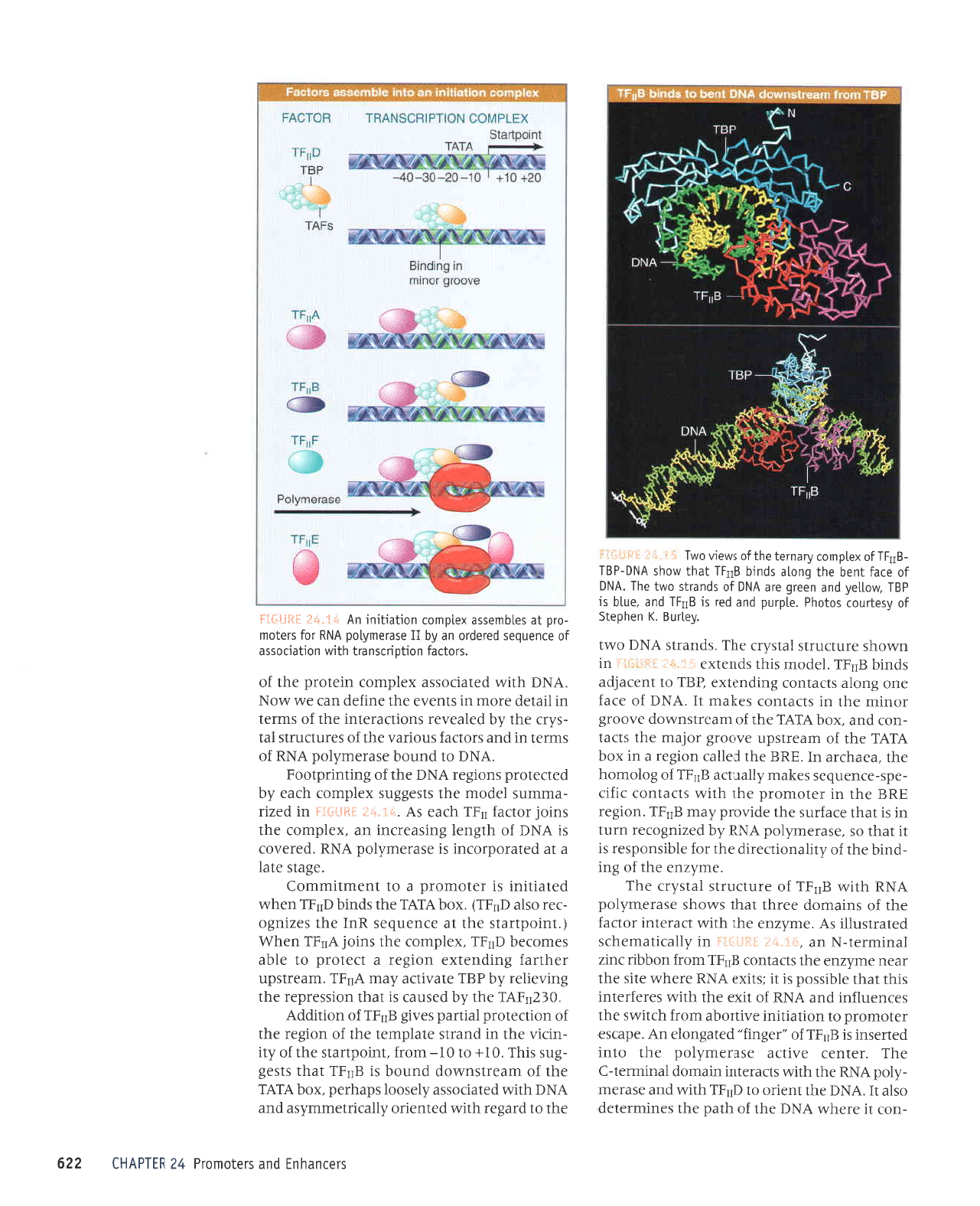
tacts the factors TF1E, TFrrR
and TFnH, which
may align them in
the basal factor complex.
The factor TFnF is a heterotetramer
consist-
ing
of two tlpes of subunit. The larger subunit
(RAP74)
has an AIP-dependent DNA helicase
actMtythat couldbe involvedinmelting
the DNA
at
initiation. The
smaller subunit
(RAP38)
has
some
homology to the regions
of bacterial sigma
factor that contact the core
polymerase;
it
binds
tightly to RNA
polymerase
II. TFrrF
may bring
RNA
polymerase
II to
the assembling transcrip-
tion complex and
provide
the means
by
which it
binds.
The
complex of
TBP
and TAFs may inter-
act with
the CTD tail
of
RNA
polymerase,
and
interaction with
TFnB
may also be important
when TFnF/polymerase
joins
the complex.
Polymerase
binding extends the sites that
are
protected
downstream to +I5
on
the tem-
plate
strand and
+20
on the nontemplate strand.
The enzyme extends the full length
of
the com-
plex
because additional
protection
is
seen at
the
upstream
boundary.
What happens at TAIA-less
promoters?
The
same
general
transcription factors, including
TFnD,
are
needed. The Inr
provides
the
posi-
tioning
element; TFID
binds to
it
via an ability
of one or more of the TAFs to recognize the Inr
directly. Other TAFs in TFnD
also
recognize the
DPE
element downstream
from
the startpoint.
The function of
TBP
at these
promoters
is more
like that at
promoters
for RNA
polymerase
I and
at internal
promoters
for RNA
polymerase
III.
Assembly of the RNA
polymerase
II initia-
tion complex
provides
an interesting contrast
with
prokaryotic
transcription. Bacterial RNA
polymerase
is essentially a coherent aggregarc
with intrinsic ability to bind DNA; the sigma
factor, needed for initiation
but
not for
elonga-
tion, becomes
part
of the enzyme before
DNA
is bound, although it is later released. RNA
poly-
merase II
can
bind to the
promoter,
but only
after separate transcription
factors
have bound.
The
factors
play
a role
analogous to that of
bac-
terial sigma factor-to allow the basic
poly-
merase to recognize DNA specifically at
promoter
sequences-but have evolved more
independence.
Indeed, the factors are
primar-
ily responsible
for
the specificity of
promoter
recognition.
Only some of the factors
partic-
ipate in
protein-DNA
contacts
(and
only
TBP makes sequence-specific contacts); thus
protein-protein
interactions are important
in
the assembly of the complex.
When a TATA box
is
present,
it
determines
the location of the startpoint.
Its
deletion
causes
the site
of initiation to become erratic, although
S;,+-J#.il
:i',;" iii TfuB binds
to
DNA and contacts
RNA
poty-
mer,ase near the RNA exit
site and
at the
active center,
and
orients
it on DNA. Compare
with Figure
24.17, which
shows
the
potymerase
structure
engaged
in transcription.
anlr overall
reduction
in transcription
is rela-
tively small.
Indeed, some
TAIA-less
promoters
Iacl<
unique
stafipoints;
initiation
occurs
instead
at any one of a
cluster
of startpoints.
The TAIA
box aligns the
RNA
polymerase
(via
the inter-
act:ion with
TFnD
and other
factors)
so that
it ini-
tiales at
the
proper
site.
This explains
why
its
location is
fixed with
respect
to the
startpoint.
Birrding of TBP to
TATA
is the
predominant
fea-
ture
in recognition
of the
promoter,
but
two
Iar;3e TAFs
(TAF1250 and
TAF1I50)
also con-
tact DNA in the
vicinity of
the startpoint
and
inf luence the efficiency
of the
reaction.
Although
assembly
can
take
place
iust
at
the core
promoter
invitro,
this
reaction
is not suf-
ficient for transcription
in
vivo, where
interac-
tions with
activators
that
recognize
the
more
upstream
elements
are
required.
The activators
interact with
the basal
apparatus
at various
stages
during
its assembly
(see
Section
25.5,
Activators
Interact
with
the
Basal Apparatus).
Initiation
Is
Followed
by
Promoter
Clearance
I
TFnE
and
TFnH are
required
to
mett
DNA to atlow
po[ymerase
movement.
.
Phosphorytation
of the
CTD
may be
required
for
etongation
to begin.
o
Further
phosphoryl.ation
of
the CTD
is
required at
some
promoters
to
end abortive
initiation.
r
The CTD may coordinate
processing
of RNA
with
transcri otion.
24.11
Initiation
Is Fottowed
by
Promoter
Ctearance
623

Most
of the
general
transcription
factors
are
required
solely to
bind RNA
polymerase
to the
promoter,
but some act at a later
stage. Binding
of TFnE
causes the boundary
of the region
pro-
tected
downstream
to be extended
by another
turn
of the double helix,
to +30.
Two further
factors,
TFnH
and
TFnJ,
join
the complex after
TF1E.
They
do not change
the
pattern
of bind-
ing ro DNA.
TF11H is
the only
general
transcription
fac-
tor
that has multiple
independent
enzymatic
activities. Its
several activities
include
an
AIPase,
helicases
of both
polarities,
and a kinase
activ-
ity that
can
phosphorylate
the
CTD tail of RNA
polymerase
II. TFrrH
is an
exceptional factor
that
may also
play
a role in
elongation.
Its interac-
tion with
DNA
downstream
of the startpoint is
required
for RNA
polymerase
to escape from
the
promoter.
TFnH
is also involved
in repair
of
I
o
[YSPTSPS]n
ilfiiJiiil
il,i"t
i"
Phosphorytation
of the CTD
by the kinase
activity
ofTFllH may
be needed
to release
RNA
potymerase
to
start transcription.
CHAPTER
24
Promoters
and Enhancers
damage to DNA
(see
Section 24.I2,
A Connec-
tion
between Tfanscription and
Repair).
The initiation reaction,
as defined
by for-
mation
of the first
phosphodiester
bond,
occurs
once RNA
polymerase
has
bound. iliSl"{fi{: :4..ti=
proposes
a model in which
phosphorylation
of
the tail is needed to release
RNA
polymerase
II
from the transcription factors
so
that
it
can make
the transition to the elongating
form.
Most of
the
transcription factors are released
from
the
promoter
at this
stage.
On a
iinear
template, ATP hydrolysis,
TFnE,
and the helicase activity
of
TFIH
(provided
by
the XPB subunit)
are
required
for
polymerase
movement. This
requirement
is bypassed
with
a supercoiled template.
This suggests
that TFrrE
and TF1IH
are
required
to melt
DNA to
allow
polymerase
movement to
begin. The helicase
activity
of the XPB subunit
of TFnH is
respon-
sible for the actual
melting of DNA.
RNA
polymerase
II
stutters
at some
genes
when it
starts transcription.
(The
result is
not dis-
similar to the abortive
initiation
of bacterial
RNA
polymerase
discussed in
Section I l.l l,
Sigma Factor
Controls Binding
ro DNA,
although
the mechanism is
different.)
Ar many
genes,
RNA
polymerase
II
terminates
after a
short
distance. The
short RNA
product
is
degraded rapidly. To
extend elongation
into
the
gene,
a kinase
called
P-TEFb
is required.
This
kinase
is a member
of the cdk family
that
con-
trols the
cell cycle. P-TEFb
acts on
the CTD to
phosphorylate
it further.
We
do not
yet
under-
stand
why this effect is
required
at some
pro-
moters
but not
others or how it is
regulated.
The CTD may
also be involved,
directly
or
indirectly,
in
processing
RNA
after it
has been
synthesized
by RNA
polymerase
II.
ilIirrtft.il f
4.1*
summarizes
processing
reactions
in
which
the
CTD may
be involved.
The capping
enzyme
(guanylyl
transferase),
which adds
the G residue
to the 5'end
of newly
synthesize
d mRNA,
binds
to the
phosphorylated
CTD: This
maybe
impor-
tant in enabling
it to modify
the 5'end
as soon
as it
is synthesized.
A set
of
proteins
called
SCAFs
bind to
the CTD, and
they may in
turn
bind to
splicing factors.
This
may
be a means
of
co-
ordinating
transcription
and
splicing.
Some
components
of the cleavage/polyadenylation
apparatus
also bind
to the CTD.
Oddly
enough,
they
do so at the
time of initiarion,
so that
RNA
polymerase
is
ready for
the 3'
end
processing
reactions
as
soon as it
sets out!
AII of
this sug-
gests
that the
CTD may
be a
general
focus
for
connecting
other
processes
with
transcription.
In
the cases
of capping
and splicing,
the CTD
624
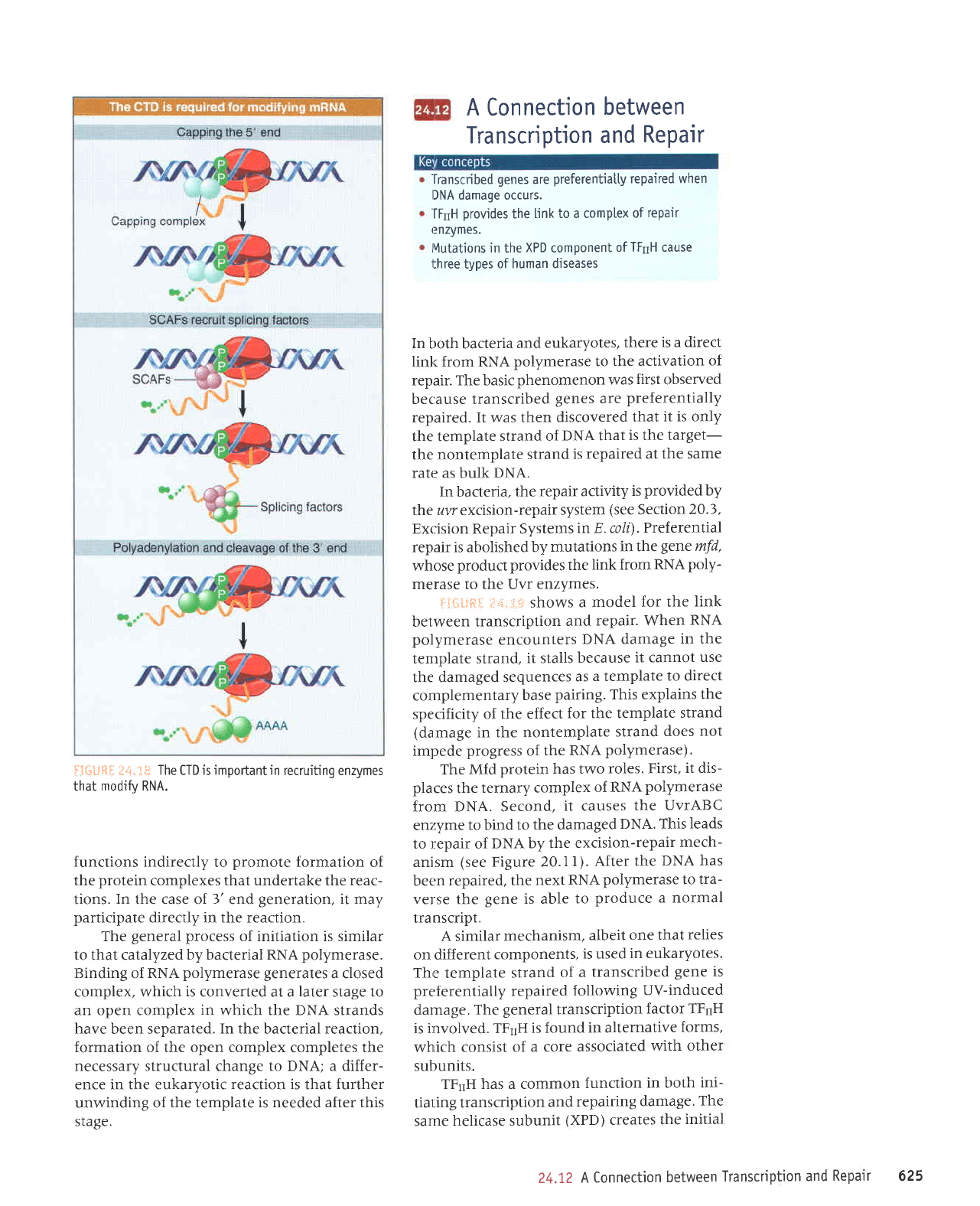
FiilliftS
,ir+.1$
The
CTD
is importantin
recruiting enzymes
that
modifv
RNA.
functions indirectly
to
promote
formation of
the
protein
complexes that undertake the reac-
tions.
In
the
case
of
3'
end
generation,
it may
participate
directly in the reaction.
The
general process
of initiation is simiiar
to that catalyzed
by bacterial RNA
polymerase.
Binding
of
RNA
polymerase generates
a closed
complex, which
is
converted at a
later
stage to
an open complex
in
which the DNA strands
have been separated.
In the
bacterial
reaction.
formation of the open complex completes the
necessary structural change to DNA; a differ-
ence in the eukaryotic
reaction
is that
further
unwinding of
the template is needed after this
stage.
A Connection
between
Transcription
and
Repair
'Iranscribed
genes
are
preferentiatty
repaired when
DNA damage occurs.
'fFnH
provides
the
tink to a comptex
of
repair
Lrnzymes.
lMutations in the
XPD
component
of TFnH cause
lthree
types
of human diseases
In
troth bacteria
and eukaryotes,
there
is a direct
linl< from
RNA
polymerase to the
activation
of
repair. The basic
phenomenon was
first
observed
because transcribed
genes
are
preferentially
repaired. It was then
discovered
that
it is only
the template
strand of
DNA that
is the target-
the nontemplate
strand
is repaired
at the same
rate as bulk DNA.
In bacteria.
the repair
activity
is
provided
by
the zw excision-repair
system
(see
Section
20.3,
Excision Repair Systems
in E . coli)
. Pref erential
repair is abolished by
mutations
inthe
gene
mfd,
whose
product provides the
link from RNA
poly-
merase to the Uvr
enzymes.
l:i:!*fi:li
:lii.li
'i,l
shows
a model
for the
link
between
transcription
and
repair. When
RNA
polymerase
encounters
DNA
damage
in the
template strand,
it stalls
because
it cannot
use
the damaged
sequences
as
a template
to direct
cornplementary
base
pairing. This explains
the
specificity of the
effect
for the template
strand
(damage
in the nontemplate
strand
does
not
impede
progress
of the
RNA
polymerase).
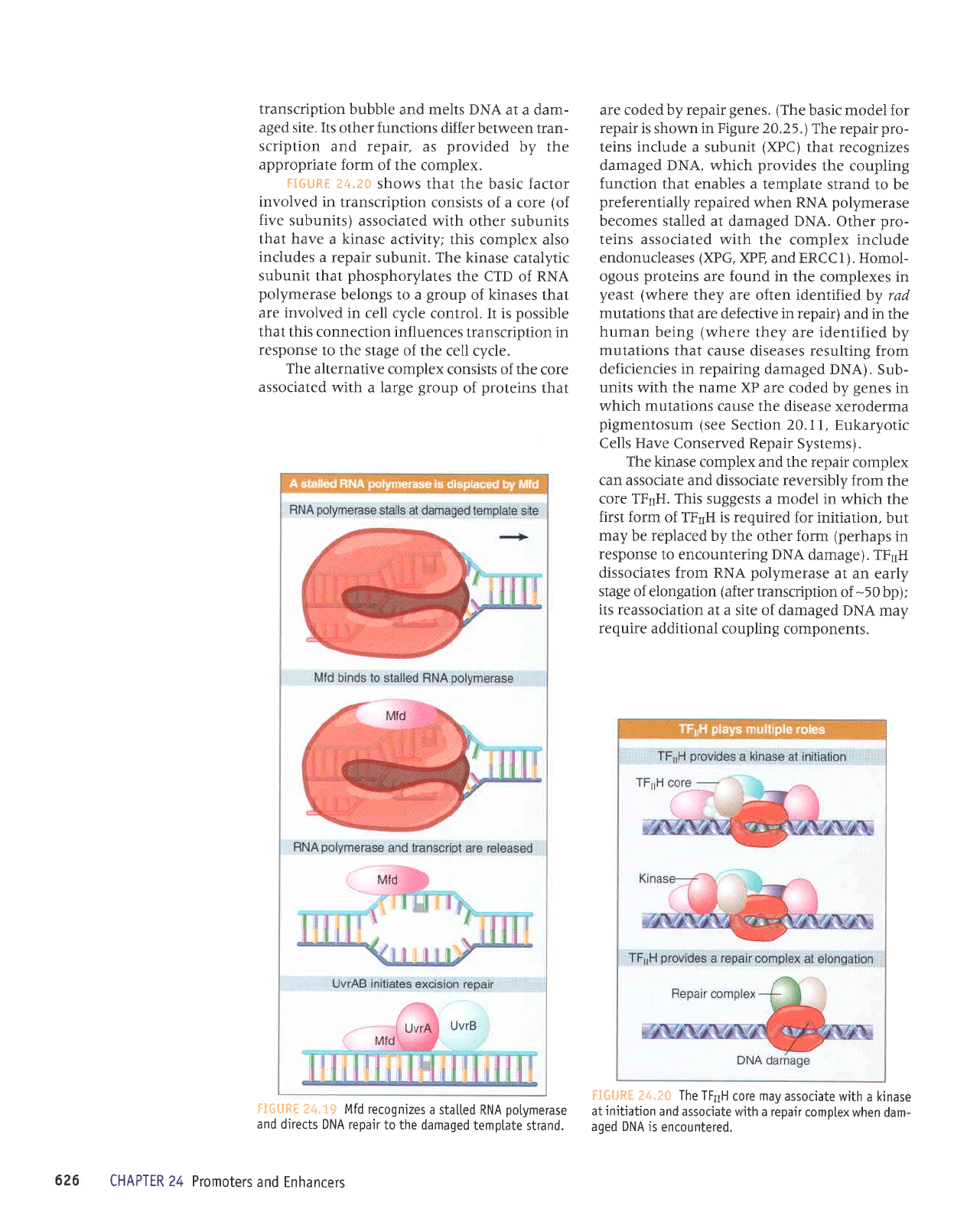
The Mfd
protein
has
two
roles. First,
it dis-
places
the
ternary complex
of RNA
polymerase
from DNA. Second,
it causes
the
UvTABC
en,ryme
to bind to
the damaged
DNA. This
leads
to r:epair of
DNA by the
excision-repair
mech-
anism
(see
Figure 20.I1).
After the
DNA has
been
repaired, the
next RNA
polymerase
to tra-
verse
the
gene
is able
to
produce a normal
transcript.
A
similar
mechanism,
albeit one
that
relies
on different
components,
is used in
eukaryotes.
The template
strand
of a
transcribed
gene
is
prerferentially
repaired
following
UV-induced
damage. The
general
transcription
factor TFnH
is i:nvolved.
TFIH
is found
in alternative
forms,
which consist of
a core
associated
with
other
sutrunits.
TFnH
has a common
function
in both
ini-
tiating transcription
and
repairing
damage.
The
sarne
helicase subunit
(XPD)
creates
the
initial
24.1,2
A Connection
between
Transcription
and
Repair 625
transcription
bubble and melts DNA
at a dam-
aged site. Its
other functions
differ between tran-
scription
and repair,
as
provided
by the
appropriate
form
of the complex.
FIGURn
?4.20
shows that the
basic factor
involved
in transcription
consists of a core
(of
five
subunits)
associated with
other subunits
that have
a
kinase
activity;
this complex also
includes
a repair
subunit. The kinase
catalytic
subunit
that
phosphorylates
the CTD of RNA
polymerase
belongs to a
group
of
kinases
that
are involved
in cell cycle
control. It is
possible
that this connection
influences
transcription in
response
to the stage of
the cell cycle.
The
alternative complex
consists
of
the
core
associated
with
a
large
group
of
proteins
that
i:E*iJRS
f;4.1#
Mfd recognizes
a
stalted RNA
potymerase
and directs
DNA repair
to the damaged
temptate
strand.
CHAPTER
24 Promoters
and Enhancers
are coded by repair
genes. (The
basic model for
repair is
shown
in Figure 20.25
.\ The repair
pro-
teins
include
a subunit
(XPC)
that recognizes
damaged DNA, which
provides
the coupling
function that
enables
a
template strand
to be
preferentially
repaired when RNA
polymerase
becomes stalled
at damaged DNA. Other
pro-
teins associated
with the complex include
endonucleases
(XPG,
XPF,
and ERCC I
).
Homol-
ogous
proteins
are found in the
complexes in
yeast
(where
they are often identified
by rad
mutations
that are defective in repair)
and in
the
human
being
(where
they are identified
by
mutations
that cause diseases resulting
from
deficiencies in repairing
damaged DNA).
Sub-
units with
the
name
XP are coded
by
genes
in
which
mutations
cause the disease
xeroderma
pigmentosum
(see
Section 20.1I, Eukaryotic
Cells
Have
Conserved Repair
Systems).
The kinase complex and
the repair
complex
can associate
and dissociate reversibly
from the
core TFnH. This
suggests a model
in which
the
first
form of TF11H is required
for initiation,
but
may
be
replaced
by the other form
(perhaps
in
response
to encountering DNA
damage).
TF11H
dissociates from RNA
polymerase
at an early
stage of elongation
(after
transcription
of
-50
bp);
its reassociation at
a site of damaged
DNA may
require
additional coupling
components.
fI6{J*[ ?1+.8* The
TFrrH
core
may
associate
with
a
kinase
at
jnitiation
and associate
with a repair
comptex
when dam-
aged DNA
is encountered.
626
LZ9
sro]e^qlv
purg
sluauall aluanDas
iloqs
€I'tz
'saxoq
l9
puP
'lvvl 'v]v]
aql 01
puods
-alor
asaq-l'uor]dulsuerl
alerlrur ol
papaau
are
leq]
(oO-
pue
'91-'oE-
1e
paraluar)
suoLbal
loqs
oolql sourluopualouord
urqo16-$ aq1lo uoLbar uearlsdn
aqllo stsauabelnu
uotlelnlPs
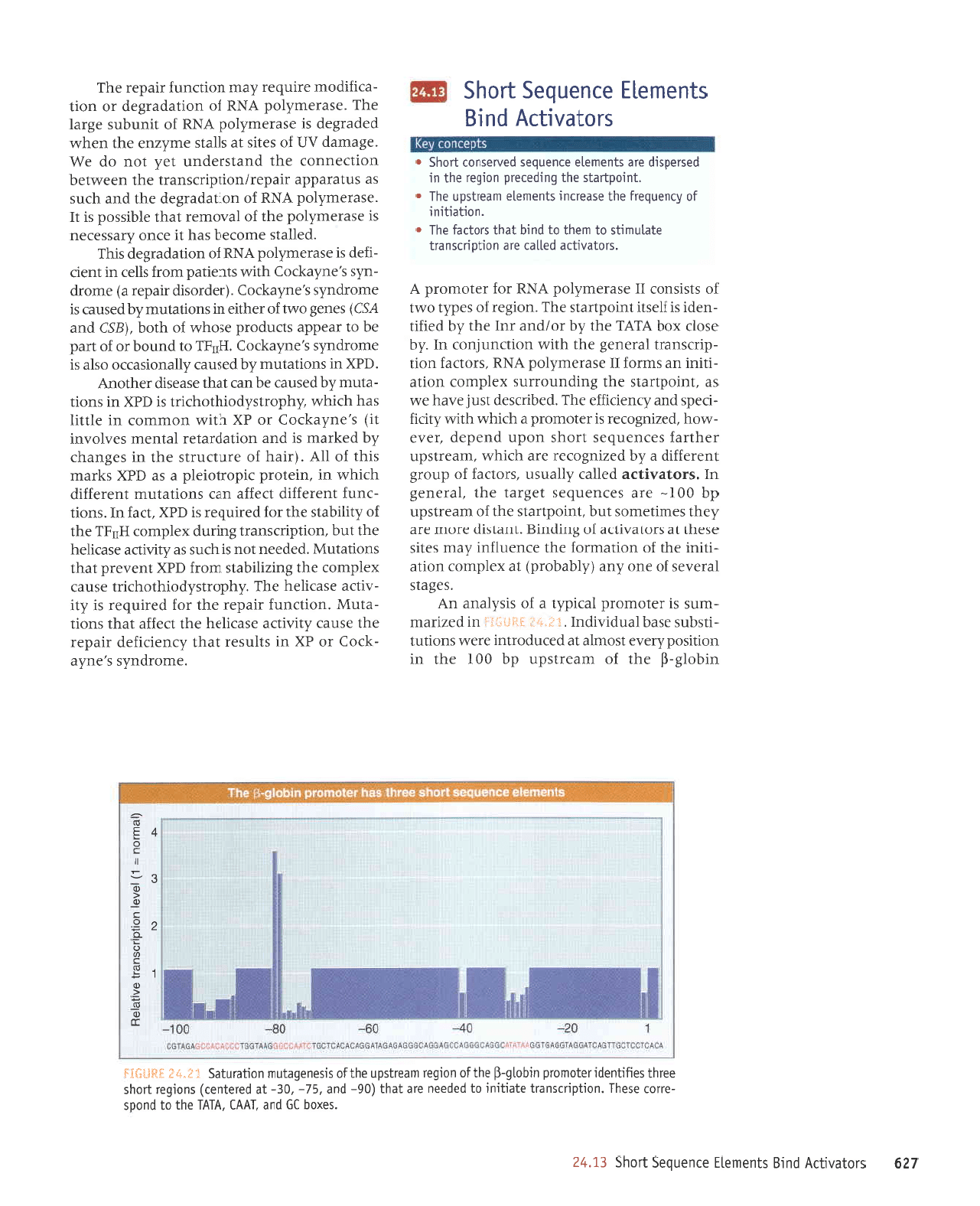
E*.irtr SHSllii
utqo13-f, eqt
Io
rupansdn dq
OOt
aq1 uI
uorlrsod zi.ra,La
lsorule
le
peJnporlur
aJJM suorlnl
-llsqns
JS€q
IenpIAIPUI
'l.i':.1.i]
"*ltf":li
j,i
uI
pezIIPIu
-IUns
sr ratoruord
prrdAr
e
;o
srsLleue
uy^
.SJtsP1S
Ipranes
Jo
euo,{ue
(,{lqeqord)
te
xalduror uolte
-lllul
aql
Jo
uollPluJoJ Jql JluJnl;ur
,{eru s:1rs
eseql
le
sJolP^rlre
1o
Surpurg
'luelsrp
eJour JJe
.daq1
saurtaruos
tnQ
'lurodlrels
aqt
yo
ruearlsdn
dq
OOt-
are saluanbas
ta8ret
aqt
'1e:aua8
uI'sJolplllJp
pall€)
.{.11ensn'sJopeJ
;o
dnor8
lueJJJJrp
e ^dq
pazruSoJeJ
aJp
qJIqM
'urearlsdn
Jequpl
saruanbas
1;oqs
uodn
puadap
'rarla
-,r,toq
'pazruSoJJr
sr Jatourord
e
qrrqa,r qllM ^]IJIJ
-oads
pue
.druaog;a
JqJ'paqrDsep
tsnf
arreq
a.u
se
'luroduels
aql Surpunorrns xaldruoJ uorle
-ltpr
ue
sruJoJ
II
aseraru,{1od
yNU
'sJotJpJ
uoll
-drnsuerl
leraua8
Jqt
qtrM
uorrrunluor
u1
'^dq
esol) xoq
vJvJ
rq],(q
roTpue rul
rql trq
palJtr
-uJpr
sr;1ast1
turodgels
eqJ
'uor8ar
yo
sad.dl
orr,rl
Jo
slsrsuoJ
1
aseraur.{1od
VXU
ro; raloruord
y
's.lolp^rllp
paller
0.lp uorldulsuprl
alelnurrls
01 uaql 0l
putq
lPql
sro]lPJ
arlf
o
'u0qetltut
1o
r{ruanbarl
oql ospollur sluauolo uearlsdn
aq1
.
'lurodyels
aq1 6urparerd uorbal eq1
ut
pasradsrp
aie sluaurolo eruenbes
palrosuol
iloqs
.
slole^qlv
pu!B
'aruorpu,{.s
s,au.de
->lJo)
ro
dX
uI sllnser
leql
.druapt;ap rtedar
Jq1 JSneJ
z(turrDe eseJrlaq
Jql
peJJp
teql
suoll
-plnw
'uolpunJ
rredar eql JoJ
partnba:
st ,{1r
-ArlJe
asef,rlJq
aq1'Iqdorlsz{.poqloq)IJt
asnpf,
xalduor aqt Sulzlgqels
uorJ
q4y
luanard teql
suorlelnw
'pepaJu
lou
sI rpns se
,{tmurp JSPJIIJq
Jql
lnq'uorldrnsuerl
Surrnp
xaldruor
HII4J
Jql
yo
Llqrqels aql
ro;
parrnbar sl
qdx
'peJ
uI'suoll
-JunJ
luJrJJJIp lJaJJP
UPJ suollelnru
lueJeJJIp
qrlqM
u1
'ulatord
rtdorlorald
e sp
OdX
s>lJelu
srqt
Jo
IIV
'(rleq
Jo
arntJnrts
Jql ul sa8ueqr
,dq
paryeru
sI
pup
uollepJelal
Ielueu
seAIoAuI
t1)
s,aude>po)
Jo
dX
qlIM
uoluuoJ
uI elllll
spq
qJrqu Lqdorlsz(polqtogJlrl
sl
11dX
uI suoll
-e1nu.dq
pasneJ
aq
ueJ
leqt
aseasrp
JJqlouV
'odx
ul suolletnru
Lq
pasner
[leuotserro oqe
st
aruorpuls
s,au.depo3
'HIISI
ot
punoq
Jo
;o
ged
aq or readde
spnpord
rsoqm
Jo
qtoq
'(aSl
pue
y5;l)
saua8 omtJo
raqtla ur
suollelnur,{qpasner
sr
aurorpuds
s,au.de1ro3
'(raprosp
rreda; e) auroJp
-uLs
s,aur(e1ro)
qlIM
sluaped
ruorJ slleJ uI
IUJIJ
-lJep
q
aseraru,{1od
yNu
Jo
uolleperSap sql
'pallpls
JruoJaq
spq
lI
JJuo
z(.ressarau
sr
aserarur(1od aqr
1o
IPAOTIIJJ
1eq1
alqtssod
s1
l1
'aseraurLlod
VNU
Jo
uolteper8ap
aqt
pup qJns
se snleredde
rredar/uorldrJJsueJt
eq1 uJeMleq
uorllJuuoJ
aq]
puPlsrJpun
1a,{ }ou
op JM
'a8eruep
An
Jo
salls
te
sllpts aru.dzua
aqt ual{,lr
paper8ap
sr aseraru^,{.1od
VXU
yo
tlunqns
a8rel
JqJ
'espreu.{1od
y1qg
yo
uotleper8rp
ro uoll
-eJrJrpour
artnba.r
,{eru uorlrun;
rredar
aq1
T
o
0)
o
J
a
o
vo
5
o
o
81
ll
f
rl
v=
3
quauall
aluanbas
uoqs

sleluequl
puP
sralourord
tz
ulldvHl
'sluoru0l0
r0q10
puP
'saxoq
lg
'saxoq
lvvl
'soxoq
vlvl
l0
su orlPu
rq
u ol
lu
ele#lp
u
rpluol
staloutold
i t.:.? t
:i
ii
{l *I
j
-8orar,(aql
teqt
tuJruela
eqt
Jo
^doJ
e spq
teqt
:atoruo.rd . uB
o1 Jlqplrp^e
eq ot
paunsse
eJe
snotrnbrqn
ssal Jo JJotu
JJe
leql
sroleArlJV
'lurodpets
aq1
ruoJJ
JJUeISIp
Dexa
Slr
pup pJluarJo
sr ureurop
Supurq-y51(
slr
qJrqM
ur
.{e,r,r
aqt
Jo
a.rrDads
-arrr
snleredde
leseq
Jql
qlrm
slJpluoJ
uralord
-uralord
J>leru
ot
tl
,vroll€
o1
q8noua
JlqrxelJ
eq
lsnul
JOIJPJ
e
Jo
elnllnJls
aqt llurodtrels
aql
Jo
^trurJrl
eq1 otul
srolJpJ
uortduJsupJl
Surrq
ot setrs Surpurq-y51q
se L1a1os
uortJunJ
stueru
-ale
aqt
teqt
sarldur
srqJ
'uortpluJrJo
JeqlrJ ur
uorpunJ
ot alqe
Jq ol rures
sJxoq
J\rv)
pue
)c
eqt
'(uortrJrrp
LupeJtsuMop
aqt ur ,{.1uo
spaar
-o.rd
uorldrnsuerl
)
uorleruJolur
IeuorlJarrp
s.{a,t
-uoJ
rJtourord
aqr
q8noqrly
'sratouord
aqt;o
IIe
o1 uolutuoJ
sr
luJruelJ
oN
'uorleluJrJo
pue
/uortprol
taqunu
ur JJJJTp raloruord
Ipnprnrpur
^,{ue
ur
punoJ
stueuele
aq1
'(luaurala
dq
g
ue)
rJruetro
aql
pue
',saxoq
Jvvf,
',s3xoq
)D
'VIVJ
:s:aloruo.rd
JSJqI ur
raqlaSotle
punoJ
JJp stuJru
-ala
Jo
sadz(1 rnog
'iir1'?i:
.j+*i!:ji:i
ur
pazrreururns
are
saldurexJ
euros
'sraloruord
IIe
JoJ
Ierl
-uesse
sr euou
lnq
'uorltunJ
JelouoJd
ol Jlnqul
-uoJ
ueJ
slueluale
Jo
,(talre,L
v
,,'qllpur
pue
xnu,,
Jo
aldnurrd
e uo
pazrue8Jo
JJp srJloruoJd
'punoq
a.lP
leql
slollPJ
l0ql0
aql
Jo
lxaluor
eq1 r\q
paurutalap
eq Aeu ralouotd
relnrqled
Aue
1e
pasn
sr
lpql
lolreJ
aql
pup
'srolrpJ
a1dL11nu fq
pazru6orat
alp slueulola
ouog
r
'uoqellrur
Juaoq_la
ro1
luasatd
oq
lsnut
slueulolo
a.lour lo
auo
qbnoqlle
'uorllunJ
talourord
ro]
lerluassa
sr
luouale
ueerlsdn
lenpurpur
ofrl
o
tuPlrodul
ag
uPl
]xatuol
lnq
alqxeH
sI
'luJuodruoJ
raloruord
uoruruo).{.lanuelar
e sr
'oot 'xoq
)D
aqJ'uorl
-eluarJo
JequJ ur JnJJo.{aqt
pue
taloruord aql
ur
tuasard
are sardor aldrllnru UJ]JO
'gD)CgC
aruanbas rqt
suretuo)
0!-w
xoq
f,D
JqJ
'^u)lJIrJos
slr eJuenlJur
lou
saop
lnq
'rJtoruord
aqt
;o
Lruanr;;a
aqt
Suruturatap
ur elor Suorts e s.{e1d xoq
Jyy)
Jqt
]eql
slsa33ns suortetnru or ,(rlllqlrdarsnS
'uorleluJrro
JaqlrJ
ur suorDunJ
t1
'turoduels
aql
ruory .r{.lqeraprsuoJ ,{re,r
teql
saJuelsrp
le
uoll
-)unJ
uel
1l lnQ
'gg-
o1 Jsolr
prlerol
uauo sl
tI
'pJqrDSJp
Jq 01 slueluele uoruuroJ
lsJrJ
Jq1
Jo
Juo se,t.r
11
'a:uanbas
snsuesuoJ
sU
ro;
pJrueN
'xoq
IVVf,
rql sr
g/-
1e
a:uanbas aq1
'(snrereddv
IPSPg
er{l
qlIM
IJPJJ]UI
SJO1PAIDV
,5'EZ
UOI1JJS AAS) XAld
-uroJ
uorlertlur ue olur .4.1qurasse
;o,,{ruanry;a
aql
JJuequJ
ol sJoDeJ uorldrnsuerl
leraua8
Jr{l uo
.dlnarrp
Suqre dq.{1a>lll
lsour
'uopeppl
}o
huanb
-a$
at4 JruJnllur
urperlsdn requeJ slueruJlJ
aruanbas
aq1
'xaldruo)
Ieseq
aql urroJ ol
JIq
-ruesse
srotJeJ uorldrJ)suprt
lerauaB
eql
q)rq,lr
te
ulqwll
aql ^Jrluepr z{.aq1
'1arra1
,lrol rJqlpJ
e
1e
Lluo uorterlrul
Josuods upl
lnq
'lurodlrels
3q1
Jo
uollPJol
Jql euruJelap .{Irerurrd
(ru1
pue
xoq
y1y1
aW) slueuela
Ipseq
JqI
'suorl
-lunJ
Jo
sad,{l
luarayJrp
JAeq rueqt
Jo
rueartsdn
slueluelJ
aql
pue
sluaruJlJ
Ieseq
JqJ
'JJloruoJd
eJo) er{l
Jo
luauodruor
Suruorlrsod
IerJnrJ
p
sp
xoq
VJVI
Jqt
Jo
eloJ aqt
sruJrJuoJ srqJ
'uorleJo1
asnard
Iensn
sll ruoJJ serJpl
lurodlrels
eql
'palelnru
sr xoq
VJVI
e ueqM
paluan:rd
lou
sr uorte
-1l1ur
qSnoqlle
leql
eloN
'suorlplnur
u(q
pJsne)
sI
teql
uorldrrlsuerl ur uortf,npar
eqt
.dq
parns
-eJru
se JJloruord
Jql
Jo
luJuodruoJ
a^rDJJlJ
tspal
eqt sr
(gE-
re
pararuar)
xoq
VJyJ
eqJ
'lualuela
Jeloru
-ord
1o
adLl uoruuroJ
e roJ aJuanbas snsuasuor
aql 01 spuodsarror
uJql
Jo
qJeA
'Jalou
-ord
aqt elnlrlsuoJ
06-
pup
'SL- 'O€.-
1e
parJl
-uaf,
sJJuenbas
uoqs
aaJql Jql
teqt
epnlJuoJ
J714
'sluJlualJ
Jr{t
Jo
Juo
z(1uo
ur Jn)Jo suorl
-ptnur
dn
'luroduels
Jql ol
tsesolJ
luerualJ
Jql
ueql
uotldrrJsuprt
Jo IJ^JI
Jql uo
pa;;a
ratear8
p
e^eq sluJruJIJ
rueaJlsdn o,vrl JqJ
'sluaruJIJ
eleJJsrp
uoqs
eJJql o1 SurpuodsaJJoJ
'suorleJol
aeJql ur JnJJo
suorlplnru u^\ocI
'u)tldtnsua4
ap
-tttu!
0t nlowotd
at17lo
&qrya
aLlt
patla pu
0p suo!tr
-ulrua
lsoLu
teqt
sr
tlnsar
3ur>Irrls JqJ
'tuloduels
uoqlnrlsuol
Jalou0Jd

nize. Common elements recognized
by ubiqui-
tous activators
include
the CAAT box, GC box,
and the
octamer. All
promoters
probably
require
one or more of these elements in order to func-
tion
efficiently. An activator typically binds a
consensus sequence of
<
I 0
bp, but actually cov-
ers
a length of
-20
bp of DNA. Given the sizes
of the activators, and the
length
of
DNA
each
covers,
we
expect that the various
proteins
will
together cover the entire
region
upstream
of
the startpoint
in
which the elements
reside.
In
general,
a
particular
consensus sequence
is recognized by a corresponding activator
(or
by a
member of a family of factors). Sometimes,
though,
a
particular promoter
sequence can be
recognized by one of several activators. A ubiq-
uitous activator, Oct-
l,
binds to the octamer
to
activate
the histone
H2B
(and presumably
also
other)
genes.
Oct-
I is
the only octamer-binding
factor in nonlymphoid cells. In lymphoid cells,
however, a different activator, Oct-2, binds to
the octamer
to activate the immunoglobulin
r
Iight
gene.
Thus
Oct-2
is
a tissue-specific
acti-
vator,
whereas Oct-l is ubiquitous. The exact
details of
recognition
are
not
so
important to
know
as the fact that a variety of activators rec-
ognize CAAT boxes.
The use of the same octamer in the ubiq-
uitously expressed
H2B
gene
and the
lymphoid-
specific
immunoglobulin
genes poses
a
paradox.
Why does the ubiquitous Oct-
I fail to activate
the
immunoglobulin
genes
in nonlymphoid tis-
sues?
The clntextmustbe important: Oct-2
rather
than Oct-
I may be needed to interact with other
proteins
that bind at
the
promoter.
These results
mean that
we cannot
predict
whether a
gene
will be
activated by a
particular
activator
sim-
ply
on the
basis of the
presence
of
particular
elements
in its
promoter.
Until now, we
have considered
the
promoter
an
isolated region
responsible
for
binding
RNA
pol'ymerase. Eukaryotic
promoters
do
not
nec-
essarily
function alone,
though.
In at least some
casr:s, the
activity of
a
promoter is enormously
increased by the
presence of an
enhancer,
which
conLsists of
another
group
of elements
but
is
Iocilted at a variable
distance
from those
regarded as comprising
part
of
the
promoter
itself.
The concept
that
the enhancer
is distinct
frorl
the
promoter reflects
two
characteristics.
The
position
of
the enhancer
relative
to the
pro-
moter need not be
fixed,
but can
vary
substan-
tialty.
i,i,'i,'i
:
,,
r
shows
that
it can
be either
upstream
or downstream.
In addition,
it can
function
in
either
orientation
(that
is,
it
can
be
inverted) relative
to
the
promoter. Manipula-
tions of
DNA show
that
an enhancer
can
stim-
ulate any
promoter
placed in
its vicinity.
In
natural
genomes,
enhancers
can
be located
within
genes
(that
is,
just
downstream
of the
promoter)
or
tens of
kilobases
away
in either
direction.
For operational
purposes. it is sometimes
use'ful
to define
the
promoter as
a sequence
0T
sequences
of DNA that
must
be
in a
(relatively)
fixed
loccrtion
with regard
to the startpoirzl.
By this
defi-
nition, the
TATA
box and
other
upstream
ele-
ments are
included,
but
the
enhancer
is
excluded.
This
is, however.
a
working
defini-
tion rather
than a
rigid classification.
Elements
analogous
to enhancers,
called
upstream
activator
sequences
(UAS),
are
i
jl.
Jiiir
::'.,,r
l
An enhancer
can
activate
a
promoter
from
upstream
or down-
strelam [ocations,
and
its sequence
can be
inverted
relative to
the
promoter.
24.15 Enhancers
contain
Bidirectional
Etements
That
Assist
Initiation
@
Enhancers Contain
Bidi rectiona
L
Elements
That
Assist Initiation
o
An enhancer activates the
nearest
promoter
to it,
and
can be any distance either upstream
or
downstream
of the
promoter.
.
A UAS
(upstream
activator sequence)
in
yeast
behaves
[ike an enhancer but
works
onty
upstream
of
the
promoter.
.
Simitar sequence
etements are found
in
enhancers
and
promoters.
o
Enhancers form complexes of activators
that
interact directty or
indirectly with the
promoter.

found in
yeast.
They
can function
in either
ori-
entation
at variable
distances
upstream
of the
promoter,
but cannot function
when located
downstream.
They
have a regulatory
role: In
several
cases
the UAS is
bound by
the
regula-
tory
protein(s)
that
activates
the
genes
downstream.
Reconstruction
experiments
in which
the
enhancer
sequence is
removed from
the DNA
and
then is inserted
elsewhere
show
that nor-
mal
transcription
can
be sustained
so
long
as it
is
present
anywhere
on the DNA
molecule. If
a
B-globin
gene
is
placed
on a DNA molecule
that
contains
an enhancer,
its
transcription is
increased
in vivo
more than 200-fold,
even when
the
enhancer
is several kb
upstream
or down-
stream
of
the startpoint,
in either
orientation.
We have
yet
to
discover at
what distance
the
enhancer
fails
to work.
lyzed in the
same way in Figure
24.21. There
is
a corresponding
increase
in the
density of
protein-binding
sites. Many
of these
sites are
common
elements in
promoters;
for
example,
API
and the octamer.
The specificity
of transcription
may
be con-
trolled by
either a
promoter
or an
enhancer. A
promoter
may be specifically
regulated
and a
nearby enhancer
used to increase
the
efficiency
of
initiation,
or a
promoter
may
lack
specific
regulation
but become active
only
when a
nearby
enhancer is specifically
activated.
An
example is
provided
by immunoglobulin genes,
which
carry enhancers within
the
transcription
unit. The immunoglobulin
enhancers
appear
to be active
only in the B lymphocytes
in
which
the immunoglobulin genes
are
expressed.
Such
enhancers
provide
part
of the regulatory
net-
work by which
gene
expression
is
controlled.
A
difference between
enhancers
and
pro-
moters may
be that an
enhancer
shows
greater
cooperativity
between
the binding
of factors.
A
complex that
assembles
at the enhancer
that
responds
to IFN
(interferon)
yassembles
coop-
eratively
to form
a functional
structure
called
the enhanceosome.
Binding
of the nonhis-
tone
protein
HMGI(Y)
bends
rhe DNA
inro a
structure that
then binds
several activators
(NF-
KB,
IRF, and ATF-Jun).
In
contrast
with rhe
"mix
and match"
construction
of
promoters,
all of
these
components
are required
to
create an
active
structure
at the enhancer.
These
compo-
nents
do not themselves
directly
bind
to RNA
polymerase,
but
they create
a surface
that
binds
a coactivating
complex. The
complex
helps
the
preinitiation
complex of
basal transcription
fac-
tors that is
assembling
at the
promoter
to recruit
RNA
polymerase.
We
discuss
the function
of
f:G!JRf
t4-*4
An
enhancer
contains
several structural
motifs.
The histogram
pl"ots
the effect
of
atl mutations that
reduce
enhancer function
to
<75olo
of
witd type. Binding
sjtes for
proteins
are
indicated
below the histoqram.
CHAPTER
24 Promoters
and Enhancers
7@
Enhancers
Contain
the
Same Elements
That
Are Found
at Promoters
r
Enhancers
are made
of the
same short
sequence
etements
that
are found
jn
promoters.
o
The
densjty
of sequence
components is
greater
in
the
enhancer than in
the
promorer.
A difference
between
the enhancer
and
a typ-
ical
promoter
is
presented
by the
density of reg-
ulatory
elements. Flfiilftf
E4"?r+
summarizes
the
susceptibility
of the SV40
enhancer
to damage
by mutation,
and we
see that
a much
greater
proportion
of its
sites directly
influences
its func-
tion
than is
the case
with the
Dromoter ana-
b80
Hoo
c
940
Y
o
t20
630