Lewin Benjamin (ed.) Genes IX
Подождите немного. Документ загружается.


.!l
c
=o)
gene
whose sequence is different
must have
been
generated
by somatic changes.
One difficulty is to ensure
that every
poten-
tial contributor
in
the
germline
V
gene
segments
actually has been identified. This
problem
is
overcome by the simplicity of the mouse l.
chain
system. A survey of several myelomas
produc-
ing 1"1
chains
showed
that many have the
sequence of the single
germline
gene
segment.
Others,
however, have new
sequences that must have
been
generated
by mutation of the
germline gene
segment.
To determine the frequency
of somatic
mutation in
other cases, we
need
to examine a
large number of cells in which the
same
V gene
segment
is expressed. A
practical procedure
for
identifying such a
group
is to characterize the
immunoglobulins of a series
of cells, all of which
express
an immune response
to a
particular
antigen.
Epitopes used for this
purpose
are small
molecules-haptens-whose
discrete structure
is likely to
provoke
a consistent response, unlike
a large
protein,
different
parts
of which
pro-
voke
different antibodies. A hapten is conju-
gated
with a nonreactive
protein
to
form
the
antigen.
The
cells are obtained by immunizing
mice with the antigen, obtaining the reactive
Iymphocytes, and sometimes
fusing
these
lym-
phocytes
with a myeloma
(immortal
tumor)
cell
to
generate
a hybridoma that
continues
to express the desired antibody
indefinitely.
In one example, l0 out of l9 different cell
lines
producing
antibodies
directed against
the hapten
phosphorylcholine
had the same
Vs seeuence.
This
sequence was the
germline
V
gene
segment
TI5.
one of
four related
Vs
genes.
The other nine expressed
gene
segments
differed
from each
other
and from all four
germline
members of the family. They were
more
closely
related to the Ti5
germline
sequence
than to any of the others, and their
flanking sequences were the same as those
around
TI5. This suggested that
they
arose from
the T15
member
by somatic
mutation.
Fi*tjtI
r:1.:1 I shows that sequence changes
are localized
around the V
gene
segment,
extending in a
region from
-I50
bp downstream
of the
V gene
promoter
for
-1.5
kb. They take
the form of substitutions
of individual nucleotide
pairs.
Usually there are
-3
to
-
I 5
substitutions,
corresponding
to
<10
amino acid changes
in
the
protein.
They are concentrated in the antigen-
binding
site
(thus generating
the
maximum
diversity for
recognizing new
antigens).
Only
some of
the mutations affect the amino
acid
$3{rl,iF.tf,
il;i.il
li Somatjc
mutation
occurs
in the
region
surrounding the V segment
and
extends
over the
joined
VDJ
segments.
sequence, because
others
lie
in third-base
cod-
ing
positions
as well
as
in nontranslated
regions.
The large
proportion
of
ineffectual
mutations
suggests that
somatic
mutation
occurs
more or
less at random
in
a
region
including
the V
gene
segment and
extending
beyond
it.
There is a ten-
dency
for
some
mutations
to
recur on
multiple
occasions. These
may
represent
hotspots
as a
result
of
some intrinsic
preference in the system.
Somatic
mutation
occurs
during
clonal
pro-
liferation, apparently
at a
rate
-
t
g-r
per
bp
per
cell
generation.
Approximately
half of
the
prog-
eny
cells
gain
a
mutation;
as
a result,
cells
expressing
mutated
antibodies
become
a
high
fraction of the
clone.
In many cases,
a single
family
of
V
gene
seg-
ments
is
used
consistently
to
respond to
a
par-
ticular antigen.
Upon
exposure
to
an antigen,
presumably
the
V region
with
highest
intrinsic
affinity
provides
a starting
point.
Somatic
muta-
tion then increases
the
repeiloire.
Random
mutations
have unpredictable
effects
on
pro-
tein function;
some
inactivate
the
protein,
whereas
others
confer
high specificity
for a
par-
ticular
antigen.
The
proportion and
effective-
ness of the
lymphocytes
that
respond
is
increased
by selection among
the
lymphocyte
population
for those
cells bearing
antibodies
in which
muta-
tion has increased
the
affinity
for
the antigen.
Somatic
Mutation
Is Induced
by
Cytidine
Deaminase
and UraciI
Glycosylase
A cytidine
deaminase
is required
for somatjc
mutation as we[[
as
for ctass switching.
U raci t- D
NA
g
lycosytase
activity
i nftuences
the
pattern
of somatic
mutations.
Hypermutation
may be
initiated
by the sequentiaI
action
of these
enzvmes.
23.15
Somatic
Mutation
Is Induced
by
Cytidine
Deaminase
and Uracil
Gtycosylase
59r

Somatic mutation
has many
of the
same
requirements
as
class switching
(see
Sec-
Iion23.l3,
Switching
Occurs by
a Novel Recom-
bination
Reaction:
o
transcription
must
occur in the
target
region
(as
shown
in this
case by the
demand for
the enhancer
that activates
transcription
at each Ig locus;
.
it requires
the enzymes
AID and
UNG
and
r
the
MSH mismatch-repair
system is
involved.
The
way in
which the removal
of the
deam-
inated
base leads to
somatic mutation
is sug-
gested
by the
experiment
summarized in
FiGuftE
23.2?.
When AID
deaminates cytosine,
it
generates
uracil, which
then is removed
from
the DNA
by UNG.
Normally
all
possible
substi-
tutions
occur
at the abasic
site. If
the action of
uracil-DNA
glycosylase
is blocked,
though,
we
see a
different result.
If uracil
is not removed
from DNA,
it
should
pair
with adenine
during
replication.
The
ultimate result
is
to
replace
the
original
C-G
pair
with
a T-A
pair.
Uracil-DNA
glycosylase
can be blocked
by introducing
into
cells the
gene
coding
for a
protein
that inhibits
the enzyme.
(The gene
is a component
of the
bacteriophage
PSB-2, whose
genome
is
unusual
in
containing uracil, so that
the enzyme
needs
to be blocked during a
phage
infection.)
When
the
gene
is introduced
into a lymphocyte
cell
line, there is
a dramatic change
in the
pattern
of mutations,
with almost all
comprising
the
predicted
transition from
C-G to A-T.
The key
event
in
generating
a random
spec-
trum of mutations
is therefore
to create
the aba-
sic site.
The
MSH repair
system is then
recruited
to excise and replace
the stretch
of DNA
con-
taining
the damage. The
simplest
possibility
is
that
if
the replacement is
performed
by an
error-
prone
DNA
polymerase,
mutations
may
be
introduced.
Another
possibility
is that
so many
abasic sites are
created that the
repair systems
are ovenvhelmed.
When replication
occurs, this
could lead to the random
insertion
of
bases
opposite the
abasic sites. We
don't know
yet
what restricts
the action
of this system
to the
target region
for hypermutation.
The difference in
the systems
is at
the end
of the
process,
when double-strand
breaks are
introduced
in
class switching.
but individual
point
mutations
are created
during
somatic
Cytidine
deaminase
creates
a U-G
pair
Uracil DNA-glycosylase
creates
an abasic
site
Normal
mutation
pattern
Replication
inserts
bases
at
random
opposite
the
gap
G
-+
A= 48%
G-->C=39%
G
-+f
=
12o/o
Replication
of
a
U-G
oair causes
G to A transition
+ +
Uracil DNA-
grycosyrase
BLOCKED
G-+A=84%
u+t/= to-lo
A
G
flE*l.JftFj
i3"?f When
the
action of
cytidine deamjnase
(top)
is foltowed
by that of
uraciL DNA-
gtycosytase,
an abasic
site is
created. Reptication
past
this site shoutd
insert
a[[
four
bases
at ran-
dom into
the
daughter
strand
(center).
If the
uraciI
js
not removed
from
the DNA, its reptication
causes
a C-G to T-A
transition.
592
CHAPTER
23 Immune
Diversitv

mutation. We
do not
yet
know
exactly
where
the systems diverge.
One
possibility
is that breaks
are introduced at abasic
sites in
class switching,
but the
sites are erratically repaired
in somatic
mutation. Another possibility
is that breaks
are
introduced
in both
cases, but are repaired
in an
error-Drone manner in
somalic
mutation.
rearranged V11 seeuence
has four to six
con-
verted segments spanning
its
entire
length,
which are
derived
from different
donor
pseudo-
genes.
If all
pseudogenes
participate,
this allows
2.5
x
108
possible
combinations!
The
enzymatic
basis
for copying
pseudo-
gene
sequences
into the expressed
locus depends
on enzymes involved
in recombination
and
is
related
to the
mechanism
for somatic
hyper-
mutation that
introduces diversity
in
mouse
and
man.
Some
of the
genes
involved
in recom-
bination are required
for the
gene
conversion
process;
for example,
it is
prevented
by dele-
tion of R.4D54.
Deletion of other
recombination
genes (XRCC2,
XRCC3,
andP.1'D5LB)
has another,
very
interesting effect: Somatic
mutation occurs
at the
V
gene
in the expressed
locus. The
fre-
quency
of the somatic
mutation
is
-
I 0x
greater
than the
usual
rate of
gene
conversion.
These results show
that
the absence
of
somatic
mutation in chick
is
not due to a defi-
ciency
in
the enzymatic
systems
that
are respon-
sible in mouse
and
man. The
most likely
explanation for a connection
between
(lack
of
)
recombination and somatic
mutation
is
that
unrepaired breaks at
the locus
trigger the
induc-
tion of
mutations.
The reason
why somatic
mutation occurs
in mouse
and
man but
not in
chick may therefore
lie with
the details
of the
operation
of the repair
system
that operates
on
i1{-;i.iiriIi:i,i.i
Thechickentambdalightl'ocushas25Vpseudogenesupstream
of the sing[e
functionaI
V-J-C
region. Sequences
derived
from the
pseudo-
genes,
however, are
found
jn
active
reananged
V-J-C
genes.
Avian ImmunogLobu[ins
Are Assembled
from
Pseudogenes
r
An
immunogtobulin
gene
in
chicken is
generated
by copying a sequence from
one of 25
pseudogenes
into
the V
gene
at a single active
r.ocus.
The chick immune
system is the
paradigm
for
rabbits,
cows, and
pigs,
which rely
upon using
the diversity that
is
coded in the
genome.
A sim-
ilar mechanism is used
by both the single light
chain
locus
(of
the
l"
tlpe) and the H
chain
locus.
The organization
of the
l,
locus is
drawn
in
:rii:riir::
.,'::
.i
..
It has
only one functional
V
gene
segment, J segment, and
C
gene
segment.
Upstream of the functional
V11
gene
segment
lie 25 Vr
pseudogenes,
organized in
either ori-
entation. They are classified
as
pseudogenes
because either the coding segment is
deleted at
one or both ends, or
proper
signals for recom-
bination are missing,
or both. This assignment
is
confirmed by the fact that
only the
V1y gene
segment recombines with
the J-Cr
gene
segment.
Sequences of active rearranged
V1-J-C1 gene
segments show considerable
diversity, though!
A rearranged
gene
has one or more
positions
at which a cluster of changes has
occurred
in
the sequence. A sequence identical
to the new
sequence
can almost
aiways be found in one of
the
pseudogenes (which
themselves remain
unchanged).
The
exceptional sequences that
are not found in a
pseudogene
always
repre-
sent changes at the
junction
between the orio-
inal sequence and the altered
sequence.
Thus
a
novel mechanism
is employed to
generate
diversity. Sequences from
the
pseudo-
genes,
between
l0
and 120 bp in length, are
substituted
into
the active V11 region by
gene
conversion.
The
unmodified V11 sequence is not
expressed,
even at early times
during the
immune response. A
successful conversion
event
probably
occurs every ten to twenty cell
divisions to every
rearranged
Vtl sequence. At
the end of the immune maturation
period,
a
vrlJ C
Pseudogene
sequence
replaces
corresponding
part
of V11
23.16
Avian
Immunogtobutins
Are
Assembted
from
Pseudogenes593
breaks at the locus. It is more
efficient in chick,
so that the
gene
is repaired
by
gene
conversion
before mutations
can be induced.
l@
B
Cel"L Memory Altows
a Rapid
Secondary
Response
o
The
primary
response
to an antigen is mounted
by
B
cetls that
do
not
survive beyond the response
oeriod.
Memory
B
cetls are
produced
that
have
specificity
for
the
same antigen, but that
are
inactive.
A reexposure
to
antigen triggers the
secondary
response
in which
the
memory
ce[[s are rapidty
activated.
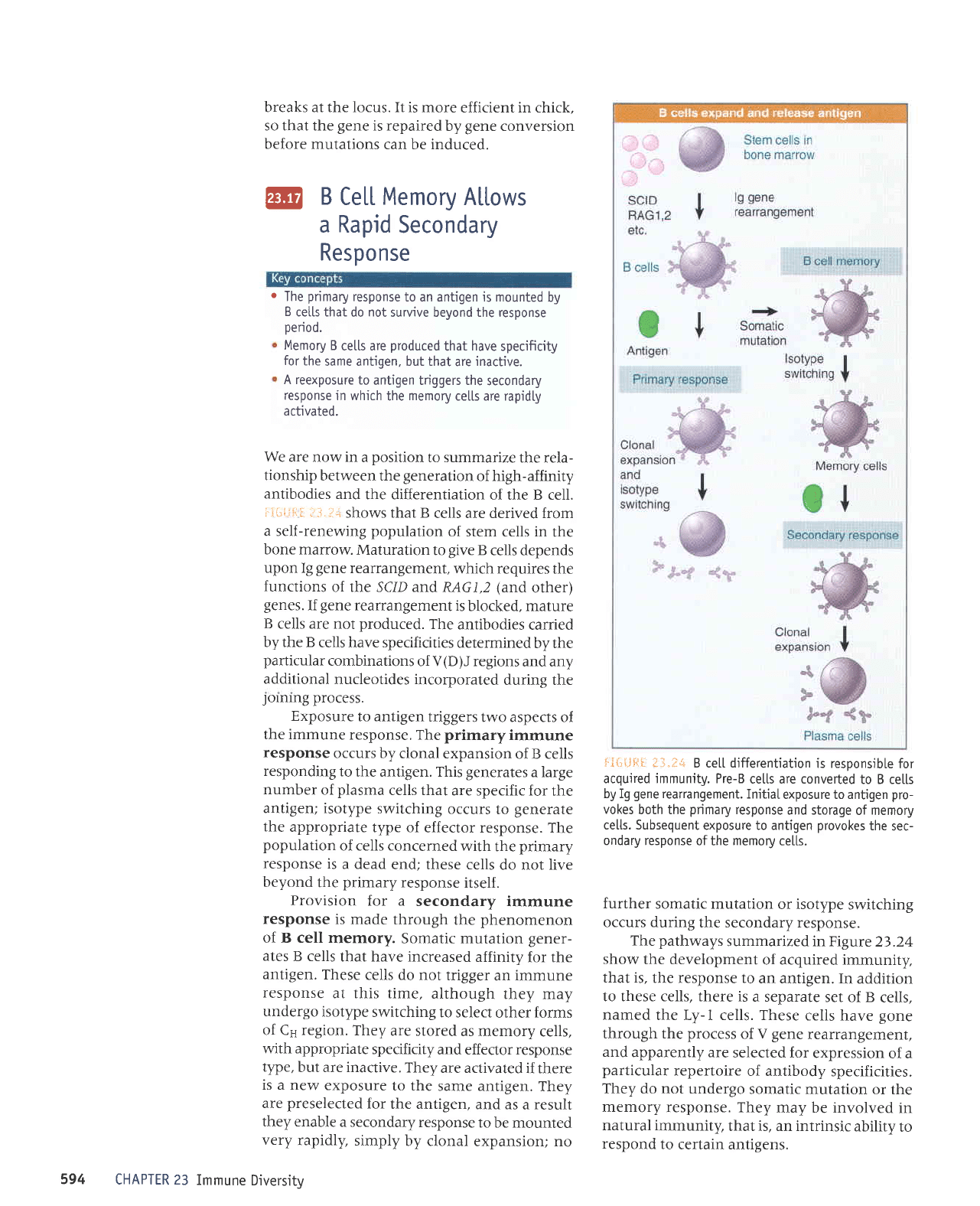
We
are now in
a
position
to summarize
the rela-
tionship
between the
generation
of high-affinity
antibodies
and
the differentiation
of the B cell.
:.r-*ii-:=l i3.l:*
shows that B
cells are derived from
a self-renewing
population
of stem cells in
the
bone marrow.
Maturation
to
give
B
cells depends
upon Ig
gene
rearrangement,
which requires
the
functions
of the SC/D arLd RAG1,2
(and
other)
genes.
If
gene
rearrangement
is
blocked, mature
B
cells are not
produced.
The
antibodies
carried
by the B
cells have specificities
determined
by the
particular
combinations
of V(D)J regions
and any
additional
nucleotides incorporated
during the
lolnng
process.
Exposure
to antigen
triggers
two aspects of
the immune
response. The
primary
immune
response
occurs by
clonal expansion
of B cells
responding
to
the antigen. This
generates
a large
number
of
plasma
cells that
are specific for
the
l#'3il;
ii,Xlli;',il51'l?"T;H,Ti#
lt :
population
of cells concerned
with
the
primary
response
is
a dead
end; these cells
do not live
beyond
the
primary
response
itself .
Provision
for a
secondary
immune
response
is made
through
the
phenomenon
:iJ;::1,ffiTi:';.":1X::#:lililf"11":
antigen.
These
cells do not
trigger
an immune
response
at this time,
although
they may
undergo
isotype
switching
to select
other forms
of Cs region.
They
are stored
as memory
cells,
with appropriate
specificity
and effector response
type,
but are inactive.
They
are activated
if there
is a new
exposure
to the
same antigen.
They
are
preselected
for
the antigen,
and as
a
result
ffI
".:ilff
,,#ilT;:"'"lT:':,
"""T::lT
3
CHAPTER 23
Immune
Diversitv
i*{i{.lFlI
t-t.:+
B ce[[ differentiation
is responsib[e
for
acquired immunity. Pre-B
cetls are converted
to B ce[[s
by Ig
gene
rearrangement.
Initial
exposure to antigen
pro-
vokes
both the
primary
response
and storage
of memory
ce[[s. Subsequent
exposure to antigen
provokes
the
sec-
ondary response
of the
memory
ce[[s.
further
somatic mutation
or isotype
switching
occurs during
the secondary response.
The
pathways
summarized in Figure
23.24
show
the development
of acquired immunity,
that is,
the response to
an antigen.
In addition
to these cells, there is
a separate
set of B
cells,
named
the Ly-1 cells. These
cells
have
gone
through the
process
of
V gene
rearrangement,
and
apparently are
selected for
expression
of a
particular
repertoire
of antibody
specificities.
They
do not undergo
somatic mutation
or the
memory
response. They
may be involved
in
natural immunity,
that is, an intrinsic
ability
to
respond
to
certain antigens.
594
969
surtnqol6ounuul
ol
pelplau
alv s.roldalau
llel
I
gI'€z
JIqe
uor8Jr JIqerJeA
e JAeq
tpql
suratoJd
JlpJa
-ue8
ot slle)
J
pue
sllJJ
g
qtoq
ur
pesn
sr JJnp
-JJoJd
Jplrrurs
V
:rusrunuoddo
.d:euorlnlolJ
Jo
aldurexa
ue sluasard
a8eaurl adroqdru.dl aql
.o/og>
uo
punoj
sr
gI
931
:sa1r{roqduAl
11o
0/096<
uo
punoJ
sr
dn
UI
.
'l0l0elel
llar
I
Jo
sad[1 orvrl
Jo
taqlra arnpord ol slla]
B
ol
bururot
3-g(6)4
Jo
ursrueqtau
lplulrs
p
asn sllal
f
r
surlnqol60unuul
ol
palelau
arv
t
t
ir:
age6orrns
:iq0r1
d
:^AeaLl
roldecer-g-er6
pessetdxe
sr ureqc
ele6ortns
1qDt1
t
sllac
10[
x
I
t
'"-""'1fl:1i";
;;ac
g-eld
e6re1
sroldalau
llal
I
'sllJJ
I
lJolJq
qllM
sllJJ
g
pJlPArlJP-ue3rlue
Jo
uortJeralur
Jql Jterpau
ot'aldruexa roy
'srotdarJJ
Jeqto
qlIM
suollf,Pralut Lq
paruenlJul
oslP sI
U)g
eql
Jo
uorlelrtJe
aql'3upurq,{poqrlue-ua8rtue
ol
asuodsaJ ur
sz(e,trqled JplnlleJprtur ra33r.r1
leql
stuauodruoJ
Suqeu8rs
aqt Jpr^oJd
^JqJ
'dBI
pup
n31
pa11er
surJloJd
JueJquJrusupJl
qlrM
palprJ
-ossp
ulnqolSounruun
ue
Jo
stsrsuoJ
pup'(U)g)
roldaral
IIaJ
g
aql
pJIIeJ
sr JJeIrns
IIaJ
eq1
uo xaldruor
a^It)e aql
]eq]
sMoqs
r.:
I
,r
:i
:t*!i'!:.jl
j
'uorssardxa
eJpJJns ,{q
pue
sllJJ
g
ruorJ uorl
-apas
.,(q
qloq
uortJun;
sulnqopounrurul
'sllJJ
g
olur eJnleru
sllar
aqt se ureqJ
ri
yo
reqt
ot
peppp
sr urpqf,
,{.Aeaq
g
;o
uorssa;dxJ Jql
pue
lnJJo suorsr^rp
'(dnbl)
rourpolalaq 6uonpsuerlleu6rs
aq1
1o
satdor
oMl ol
palurt
(z1zg)
tauetlal uqnqolbounuut
upJo slstsuotroldarar
uebquellel
g
eq1
rri
il
j
:i!jr'lli.l:;j
IIJJ
JeqUnd
'slleJ
g
aJnleruurl
eql
Jo
e)PJJns
aqt
uo s:eadde uqnqolSounulu
IeuoIDunJ
puP
passardxa
uJql
sr urcqr
lq8q
's11ar
g-ard
gerus
aruoJJq
01 epr^rp sllal
Jr{l
puP
'uleqJ
fs
Jo
sISJql
-u^s
sassudJJ
uleqJ
d;o
uorpnpord aql
'auo
sP uoll
-JunJ
tou
sJop
tnq'xaldruor
uqnqolSounurur ue
sJlqruJseJ
U
'roldarar-g-ard
aql urJoJ ol uIeI{J
Laeaq
rl
aql
qrlzrt
selpl)osse
pup
eJeJJns Jql
uo
passardxa
sr
qJrqu
'urpqJ
(fS)
lq8u
aleSoJ
-Jns
aqt
pJIIeJ
uleqJ
1
e Sulqutasar uralord e
ssardxa sllal
asaqJ
'€I'€Z
arnEtg
ut ,{.lsnonard
uMoqs
sP'JnJ)o
Leru sluaruaSuBrrear arrqrnpord
pup
alrDnporduou
Jo
uorsse)Jns
p
SuIAIoAur
slueAJ
uorlPurqruoJeJ
IPJeAas
'uleqJ
Irreaq
d
e salpJaueS
qJIqA{
'uolleulqurorar
q-n
Lq
pJpJaJJns
sl slqJ
'urpqJ
,{neaq
rl
aqr
1o
sluJru
-8as
1
aqr
pue
sluelu8as
q
Jq1 uJaMlaq uoupulq
-ruoJeJ
sr dals
tsrg
Jr{J'irt'f
* ;js{:*i,J
uI umoqs sI
tuarudola,rap
IIeJ
gJo
MJIA
pellelap
JJoIuv
'sabels
lerluanbas
qbnorql
spaerord
luaudolanap
llar
I
!e'lJ
$*ft+gi

e Surureluot
qleJ
srJlsnl)
ol^t ruoJJ
pJlpJ
-edJS
stueur8as aua8
A
qtlm
'tuJJJJJrp
sr uorl
-ezruegrc
Jql
leql
sMoqs
;i,:.ill
:T::.rii:j!.j
'HBI
Jo
Jsoqt alqruJsar
$
u)t
;o
sluauoduro)
JqJ
('dproqs
ssnJsrp
JM
qJrqM
'sluJru8as
g surel
-uoJ
osle snJol srql
'sluaru8as
n
eql ol uortrppe
u1)
'sluaru8as
n1
pue
stuaru8as
aua8
nn
Jo
lJq
-runu
Jql ur
zi.1uo
sJJuJJeJJrp
Jruos
qtru
'Jsnotu
pue
ueunq
qloq
ur Jplrlurs
sr snlol Jql
Jo
uorl
-ezrue8ro
aq;
'luaru8as
aua8
3
a13urs
e sepaJ
-ard
leqt
sluaru8as
f
Jo
JJlsnlJ e
uoJJ
paleredas
sluaurSas
aua8
n
qU,lt
')t
31
yo
reqt
salquesel
n
U)J
Jo
uorlezrueS.ro
eqt
'iii:.:,:it
:i**t,ij
ur
pezrl
-eurruns
sy
'3uqrr1s
sr
sJue8 31 aqt
qtr,u
saua8
uf,I
Jo
uotlezrueSro
Jql
Jo
eJuplquJseJ
JqJ
'uoIlf,unJ
uralord
ol
pelelJr
sr aJnlJnrls
uoJlur-uoxA
'suotl:od
rrruseldoldr
pue
eueJqruarusuerl
Lq
peMolloJ
ureuop
tuelsuof,
a13urs e seq
pue
suor8ar 8I
tuBlsuor
Jqt
ol
paleler
sr uor8ar
) dlJ
aqJ
'surelord
Ult
pue
31
qtoq
ur uortezrue8ro
IeuJelur leraua8
Jrues
Jql a,Leq satuanbas
n
JqJ
'surlnqolSoununur
eqt
Jo
teqt
selquesJJ
sutalord
U)I
Jqr
Jo
uorlezrue8ro
aq1
'sJrlnupJ
31 aarqt
aqt
ur
punoJ
Jsoql
se erues
Jql are sluauodruoJ
eqJ
'a1hot4dw[1
a4t ot t{nads
ulu)oal
ulrjowEuont a f.q
n4la6ol146notq
ala
Wqi
sruawqas alatadas
qltrw'saua6
uqnqopounwlm
aql
so
[.am
atuas
a4l ut
pazua1to
st srcll
4tug
'suot8ar
luelsuo)
pue
elqerrel
Jo
asn Jql ur saua8 uqnqolSounuur
eql sJlqruJsal
saua8 rotda)JJ
IIaJ
I
Jqt
Jo
uorlezrue8ro
aq1
pue
[e.,u
etues eqt
ur
pelloseJ
sr slle)
J
pue
sllJJ
g
dq uoqruSotar
ua8rtue
yo
rualqord
aql
'eJn1
-Jnrls
alqpDrpardun
;o
ua8rlue uSraro;
e )zru
-3orar
tsnru
ufJ
e
'surTnqoFountutura>lrl
'slue^e
luSLuaEueJreaJ
luapuadapur
senlonur
pue
'srsJqtu,{s
gl,
Ulf
ur
pellolur
esoql
ruoJJ slleJ
Jo
a8eaurl alere
-das
e
dq
pazrsaqluds
sl
tI
'rotdarar
tupurruop
-ard
aql sr
u
qurq.{.9
'uorletsa8
ra4e
s,{ep
41
ol
E I
le luaredde
saruo)aq
lsJrJ tl
'arrur
u1
'gl,
ueqt
luarudolJ^ap
IIaJ
J
ur
Jalel
pazrsaqtu,{s
s1
lI
'sa1z{roqdruLllo
ohS6<uo
punoJ
sl
dn
U)J
'02
[ep
tp
qurq.{q
rsol
uaaq Lilenurn
seq
lnq
'uortplsa8
yo
s,{ep
S
I>
le
rlqppatap
rotdarar
,(po
aql sl
tl
'eJrru
uI
'luarudola^ep
IIJJ
J
Jo
a8els Lpea
ue
te
z(po
pansaqtuLs
s1
t1
'safroqd
-udl
t
lo
"/oS>
uo
punoJ
sr rotdarar
g,t aqf
'l-i:"i
i iitlilijl"*
uI
pJZIJPIU
-Iuns
se
'luarudolanap
IIaJ
J
Eurrnp
saurl
tueJeJ
-JIp
]e
pJzlsaqlur(s
a:e sroldarar
IIaJ
J
tuaJaIIIp
aql
'roldarar
IIJr
J
1o
saddi
oMl
Jo
reqlre
eJnp
-ord
s11ar
J
'suortf,unJ
rotJaJJa
;o
a8uer
llerus
e
JoJ
lunoJ)e
pue pelnurT
eroru JJe suor8ar
luels
-uoJ
speJaqaa z(lrsralrp
luerqru8rs
aprnord
ol
Alrsra^ro aunuurl
tz
ulldvHl
'slolsnlr
l-t-0
oMl aq]
1o
ureerlsdn
qI
O8Z-
ell
lpql
ql
O0g-
lano
peerds
slueu
-6as
aueb
l
Aueu suLeluor snrol
$931
eql
tiil"E;:
-4i"l*i.{
'slueubas
9A
aloul
spq
lnq
lplrurrs sr snrol osnou aql
'sluaubas
rl-t
aql
pup
sluauEes auab
1
aq1 uaamlaq art sluau.l6as
sl-C-0
aq1 ral
-sn1r
01
aq1
urqluvl
palplol
sr
luaubas
sn
y
'sluaubas
g
pue
n
pos.raoslelur
seq snlol
pu]l
uPunq eql
*+'i,,:
3$f:llE:j
I
lg
1
7p
:&euuns
D
ueu,lnH
oz
on 09 08 00t ozt ott
qr
ffiw
oo!_l"r
sc sr sc
"nsngt*fon
uorlezrue6ro uEurnq
pup
osnoyU
'raq1a6o1
pazru6orar
ale
uabque flqrqrleduorotsrq
lsoq
pue
uabrlue
1abre1
qrrqm
ur'Alrunuut
palprpauJlal,,lelrssell,,
.ro; alqrsuodsar
sr
pup
ralpl
pazrsaqlufs
s!
dr
Ull
'luaudolanep
11ar
1
ur Apea
pazLsaqlufis
sr.roldaral
gl
aql
i;'{f T#tii}I::
-..*..
"n
Pa0uerlearu6l
peOuerearul
.".,.*?dO
dn
qsa*u^ss,L{
I
e t
'C
Z
'A
L,
:fueuruns
!
ueung
€-
0
0L 0z 0e
qI
0Bz
q)i
009
.' ii *- ..
..n"
.**.,
t u.Ltgl 09>
A CIC CfC
A
??
Zd td
uollezlueolo ueulnLl
pue
osno!\

169
IHhl
aql
q]!M
uollrunluo]
ut suor]lunl
lotdalau
llel
I
aqt
6I.tz
I
JIXolol^)
'Jsuodsal
eunrurur
aql uI
pa^lo^ur
SIIJJ UAJMIEq
SUOI]JPJAIUI
qll^^
PElJJUUOJ
suollJunJ
Jo
^lJup^
e e^eq
teql
sed^tqns
IeJe
-AJs
olur
pJpl^lp
erp
sJotdJJsJ
dn
qllM
sllJl
J
llar
6uquasold
eql
Jo
olP#ns
aql uo urelold
IHW
up
Jo
o^ooi6
e u!
punoq
sr
leql
opqdad
iloqs
p
sozruEotar
lll
all
r
IHl/\l
aql LlllM
uor.punluo]
ut suorllunl
JoJ srspq eqJ
'sluJruaSueuear
n
ur
punoJ
oslP
eJe auros
lnq
'luaua8uerJeJJ
g JoJ anbrun JJP
auos
:punoJ
JJe sluerxSas
9A
IPJJAJS
',asnoru
uI
'luJruJ8ueJJpar
g
JoJ
Jsn
IPJJUJB
uI sI
lueru8Js
JueB
A
Juo
^1uo
'3uraq
upunq aqt uI
'sur€q)
g
uf,I
anule ur
punoJ
aJP sJJuJnbas
n,rnal
.r{ran
'uopsJnb
Surlsaralur ue s1
tuaruaEuerreJJ
g
aql rq
pasn
sluaru8as
aua8
A
eql
]o
eJntpu eql
'f11(A
alnlJnrls aql a,rr8
ot urpqJ g aql olq
pale.rod
-rolur
aq Leru sluaru8as
(
eql
Jo
r{log
'stueu
-8as
o)-of
eql
pup
sluaru8as aua8
l
aql uaemlaq
ar1
93-s1-sq-sq
sluaru8Js aqJ
'g(.'€Z
arn8rg
ur
,{.1snorna;d
pJlpJ}snllr
sp
'snfol
n
U]J
Jql
tp
rll
leqt
stuaru8as
z(q
papol
sl
tpnqns
g eqJ
'tuaru8as
aua8
3
qJee
JoJ stuaru8as
1
a1dr11nru
seq Suraq ueunq aqJ
'uorleluJlJo
uI
peualul
are sluaru8es
Jruos
lnq
'Ilol
)-f
IeuoIDunJ
€
seq Jsnow
'Jsnoru
pue
uPunq
ul
luaJeJJIp
sr uorlezrue8Jo eqJ
'sluaru8as
A
IpuolDun]
8-
ql1,!r
dlrsranrp 3l1r{,{1a,rqe1ar
sPq snJol sq1
lPq1
sMoqs
*il'[i: :{}.Jf,}lt$j
'sluaru8as
)-f
Jo
sJlres e
u]oJJ
pateredas
sluaruSas aua8
n
qtlM
'y
31
;o
reqr
sJIqruJsJJ
snrol
l"
eql
Jo
uoltpztue8ro
aq1
'sllJJ
J
ut
paSuerrear
are sroldalar
IIaJ
J
seaJJqM
'sllal
g
ur
pa8uerrpal
ale no1 31
teql
os
pJIIoJluoJ
sr ssarord
Jql ,t,roq Mou>l
lou
op eM
'kt'tZ
arn8r4 aas)
laporu
uollrlrp
aql '(q
.rnlo
.{lqeqord sluarua8uerreJJ
lsoW'pJAIoAur
are sarudzue
Jrues eql
lpqt
d13uor1s san8re
srql
'saua8
31 aqr
z(q
pasn
Jsoql ot
IpJItuapI
saruanbas
snsuasuoJ rarueldaq
pue
Jarrreuou
,{.q
papunorrns
JJp sluaru8as
931
Euutqruorar
eqJ
'slleJ
1ur
saua8
u)I
pue
sllal
g
ut saua8
31 aurqruorar
1eql
suollf,eer
JI{l uI
paAIoAuI
aq ot
L1a>11 eJp srusrueqJalu
Jrues aqJ
'surPq)
g
luereJJrp
q0I
Qllu
pelPrJosse
sureqf, n
luJrelFp
L}I
x
s'z
urpluoJ Suraq
uerunq eql ul sllJJ
J
zr0I
Je1
leql
Moqs .dlrsrarrtp
Jo
tuJxe
eql
Jo
sluaueJns
-eJW
'IJol
UIJ
aql
1e
JnJJo
lou
saop uollelnur
)rlpuros
reqt
sl 3I
pue
u)J
uaeMleq aJuareJJlp
V
'(saruanbas
rarueldaq
pup
Jaupuou aql
Jo
uoll
-ezyue?to
alerrdorddB
ue
,{q
paDarp)
sutoI
q-q
Lq
pale.raua8
are
qrrq,lt
'sluaru8as
CI
omt ateJ
-odrorur
sureqJ
d
xrr
aruog
'(71'97
arn8tg aas
lsrprloJlJnu
N
pue
d
Jo
lrroJ aql
ut) sluauod
-ruoJ
eseql uaemlJq
suotpunf
aql
1B
saruanbas
^tJu
JO
uorlJnpoJlul
aql uoJJ
sllnsJJ ,(ltsr:ntp
Ipuortrppe
eruos
isluaru8as
3
pue
'f '61
A
Jo
,hauerr
p
Jo
uorleurquo)
aql
ruo;; sllnsar
futs
-ralrp
JrsurJlul
'surlnqolSounrurur
uI se sruslu
-eqJJru
erups
Jql dq
pateraua8
st,{.ltsrant(
'stpn
sf
pue
sA
eql
Jo
sJJqunu Jqt
ur JJp esnour
pue
Sutaq uerunq
uJJMlJq sa)uJJaJJrp
,{po aqt
'utu8y 'luaru3as
aua8
3
e
pup
'stueur8as
1
pra,ras
'luaru8as
(
09 08 001
ql
BUinH
uolsseroxo
lo
uoncorlc
- +>
->
+
<-
++ ++
ncnr
zrZC
lC tl
osnorl
rotdarau
llal
I
aql
'pa1e:aua8
uaJq
seq a1a1e anrDnpord
e relJp
JnurluoJ .,i.eur
satuanbas
nn
Jo
uorln4s
-qns
leql fuqrqrssod
aql
1sa33ns tuarua8ue.r.rear
penurtuoJ
Jo
sJSeJ
IeJJAJS
lluJlJJlrp aq ,{.eru
snJol
rr eqJ
'peterJua8
uaaq seq JIJIIe Jlrt]np
-ord
e aruo
passarddns
sr
luaua8uerrear
1oo1
ulpqolSounrurur
se dB,u.
arues eql
qJnu ur uors
-nlf,xJ
JITellP sMoqs
snrol
fl
eqJ,'e^lpnporduou
Jo
JArDnpoJd aq Leru
'saua8
uqnqolSounrurur
yo
asoqt r>lll
'pol
U)J
Jqt
te
stuauaSuerreag
'sJnJJo
luarua8uerrear
n1-Dn
Jql uJqM
.,(1arr1ua
tsol
sr snJol
g aql JSnelJq
'alJIIe
Juo
LuB
le
Jlrsnlf,xJ ,{.1en1nru
sr roldarar gl, aqr
pue
roldarar
dn
Uff
er11
Jo
srsaqtu,{.s
teql
sarldrur
saua8
;o
luarua8ue.rre
pas:adsralur
aqJ
'tuJru
-a8uerrear
g
ot anbrun.4.1ear are.daqr raqraqa,l
uo
luaur8pnI
alrasar
lsnru
JM
'rele,lroq
lsluJru
-3as
aua8
sA
sp
surpqJ g ur
punoJ
JJe
lpql
sluau
-8as
n
aqt
peleqel
JAeq a,tr'tuasard rql ro{
'suralord
a,,lrDe a.Lr8
ueJ
(9A
se
paurlJp
eroJ
-arJqt)
aruos.dpo
teql
tnq'luaru8as
9t((
Jqt
01
paurof
aq uer
sluaruSas auaS
n1
aqt
;o
Lueru
]eqt
sr Llqrqrssod
JUO
'u,lrou>I
lou
sr
luerue8uerrear
g
pue
n
u1 stuaru8as
n
Sursooqr
ur
.dtnr;nads
'llol
l-C
aq1
1o
tuearlsdn art
lpql
(uruoqs
1ou
saue6
-opnesd
auros osle
pue)
sluauEes aue6
n
leuorlrunl ;o
laqunu
llpus
e sureluor snrol 1.611 aql
itir'i;i:
j#itiji:i

AlrsranL6
aunuul
EZ
U:lIdVHl 869
roldarar
peler)osse
sll
uaqM JorJeJur Jql ol
ITaJ
aql
Jo
eJpJJns aqt
uoJJ
leu8rs
e SulillursuBrt
ur
pJAIoAur
sr
q)rqM
/€q)
pJIIp)
surato-rd
yo
xald
-u-roJ p
qlrM
pJlprJosse
sr lotdaral
IIJJ
J
JqI
'+8(If,_'Ol
Pue
_gc)+to) rre
qrlqM
'sasselJ
alboqdruLl
1
aleredas
eql ot asrr anr8
,(aqt
uotpalas anrle8au
luanbasqns
Jql eArAJns
sa1l:oruu{ql aql
I
'(qleap
11a]
ruorJ salboruz(qt
Jql sanJser
qrlqM)
uortJJIJS
anrlrsod Surrnp
e)eJJns eql uo
pe>lurlssoJJ
ssruoJJq rerurpora
-trq
dn
U)J
a)plJns eqJ'sasseJord
anrpalas aql
Jo
llnsal
aql se sat[roqduzll
JJn]eur atuo)aq ro
slep
E-
raqe rrp raqlra
sa/.rorudql
4C
aq1.'sural
-ord
39ry
qll.tr
uortJpJJlul
J^lolur uortJales
;o
sad[1
qlog'(spue8tl-y1as-atetrdorddeut
qlla\
lrpJJtul leqt
saxaldruor
tsure8e)
uorpalas
azrrte8au
,(q
pue (pue3r1
e
pulq
ot alqp saxald
-rxoJ
Uf,J
arnleru ro;)
uortJJIJS arrrtrsod
qloq,{q
sanurluoJ ssarord
uorlernleru aq1'
sal,boru.{ql
dC
Jqt ut sJnulluof
lu:uaSuerreJJ
urpr-{J
D
'salIroudql
d(I
salereuaB
y'uo4ta1as
$
pelleJ
sr
luarudola,tap
ur
turod
slVJ'+gOl+?O)
ot -8(f,-t(:) ruorJ
pJualuor
sl Jt^loqd
-ruz[1
aqr
]eqt
os
'uo
pJuJnt
are
sauaS
gq)
pup
te)
Jqt
pue
'pa8ueueJJ
ale sureqJ
n
r{Jrqm
Suunp
'uorsrlrp
IIaJ Jo
spunol
IeJeAJS
sraSSrrl
xaldruor
eql uorJ
3ur1eu3r5
'(rvroleq
aas) xald
-ruor
EutTeuBIs
Eql
eqt
01 spurq uer{t JJrurpolJ
-taq
U)Ja;d
JqJ
'gO)
ro
t(I)
suratoJd
aJelrns
al{t
Jo
Jeqlra
passardxa
1ou
seq afroqdur(1
aqt
'a8e1s
srql
tv
'nulJard
pellpr
ureqJ
n
ale3or
-rns
Sur8uerJpJJuou
e spurq
srqJ
'ureqJ
d
Uff
JArlJp ue
uuo} ol
luaua8uerrear
sr a8els
lslr}
JqI
':
i:";*
_qsi;Tij'
ur
pezrJeruurns
se
'alLroqdrur(1
1
Jql
Io
luaudo1a.r.ap
eql 01
prlull
sr sureqf,
UJJ
leuou)un;
aleraua8
ot uorleurqruoJJU
'ulI
gn
aql
lnoqe
paulJep
eq
ol sJnssl Surtsaralur
lsoru
Jql;o
auo sr L1r
-reder
pnp
srql JoJ
srspq eqJ
'ua8rtue
u8rarol aql
JoJ se
IIJM
se
)HI I
relnllred
e roy Ltor;r;ads
seq
U)J
ua,rr8
y ('3uo1
sanprsar
ual ot
1r{3ra
;o
sluaur8er;
aleraua8
ol uralord
u8raroy aql
saper8ap
auroseatord
eql uJqM
paleraua8
sr apq
-dad
aq1)
'U)J
aql ol aprrdad
aqtruasatd
olprcs
sl
)HW
aqJ
')HW
aqt
Jo
eJeJrns
Jqt uo a.LoorS
e ur aprldad aqt
sazruSoreJ
ueqt
u)J
Jqt
pup
'ua8rtue
u8raro;
eqt uoJJ pJAuJp
aplldad
uoqs
e spulq ulrtord
)HW
eqJ
'urato-rd
IHW
lsoq
eql
pue
ua8tluB
u8taro;
eq1
qtoq
SurzruSorar
;o
z(lrlqrsuodsar
aql
t1
uodn sareld
srql
'L1ru
-nurur
palerpJru-llal
ur uorlJunJ
IIel
I
TJIIDI
JoJ
pue.{lrunurur
IeJorunq
ur uorDun}
1ar
l
radlaq
ro;
alqrsuodsar
sr .roldarar
$n
U)J
eql
'urJtoro
I
sselJ
)HW
ue .{q
patuasard
aq o1 ua8rlue
arrnbar
'seEels
lerluanbas
qbnorql
spaarord
luaudolanep
llel
t
i.t'fl :*t{{-i:};j
sller
J
rallDl
seeJeqM
'uralord
1
ssep
39141
ue,(q
patuasard
aq o1 ua8gue JJrnbJJ slleJ
J
radlag
'(uats.{S
eunturul eql
Jo
seueg .{ueyq'to; sepo)
snrol LlqrqrtedruorotsrH rofeyg
aqJ'02'€Z uolt
-ra5
aas)
JSe)
q)pa
ur
pasn
are uralord
31111
;o
sad,{.1
luJJeJJIp
lua8uuB
}o
uollPtuesard aql
JoJ stuJruaJrnbar
luara;Jrp
J^eq
,{aql
tnq
.{ea,r
srqt
ur
>lJoM
slleJ
I
rellDl
pue
sllJl
l
redlaq
qrog
'7'97
an8rl ur Llsnor,Lard
pa1er1sn11r
sp
'llaJ
Surtuasard
aqt.{q
parJJef,
uralo,rd
)HW
ue
pup
ua8rlue
u8raro; aql sazruSorar Llsnoauellnrurs
IIe)
I
eqt snqJ
'uratord
lffW
ue ,{.q
1ar
1
aqr
01
,,pJtuesrrd,,
sr
tuaru8erJ
apudad aql
'suorl
-rear
a8eneap z{.q ua8rlue Jqt ruorJ
passarord
ueeq seq
teqt
(3uo1
sprJe ourrue aArJ 01 rnoy) app
-dad
lerus
e 01 spurq Jotdarar
IIJJ
J
Jr{J
'ua8rtue
Jqt
urqtrM
(adolrda
ue) uofar
uoqs
e sazruSo
-rar
Lpoqrlue
uV
'ua8rlue
eq1 Jlpueq,{.aqt
reqr
.,(em
aq1 sr sller
1;o
sroldarar
aql
pup
sllal
g
Jo
sJlpoquup
Jql uJJMlJq J)uJJJJJlp ro[eLu
y
'uorlJPJJlUr
ua8rlue-,{poqltup palerpeur-lla)
g
ro Surqq
taB
-rpl
prlerpeu-llal
I
lslssp
slar
l
radlag
'1ar
1aB
-Jel
peDJJul
ue as.{1 ot,{.lpeder
aql ssassod s11ar
t
I
uorsserdxe
$g91
passardxe
80c
pue
to3
uotssoJoxe
DUcfeld
ureL1c
D
eleOot.rns
sputq
dHcr
uorsserdxe
Sg31
uorleurqurocoJ
pole!pau-evu
80c rcc

is
activated by binding
antigen.
Our
present pic-
ture
of the components
of the
receptor
com-
plex
on a T cell is illustrated
in
FSftljttt
j.:
jj.li;:.
The
important
point
is that
the interaction
of
the
TCR
variable regions
with antigen
causes
the
(
sub-
units
of the CDI complex
to activate
the T
cell
response. The
activation
of CD3
provides
the
means by which
either
crB or
y6
TCR
signals that
it has
recognized an
antigen. This
is compara-
ble to the constitution
of the B
cell receptor, in
which immunoglobulin
associates
with the IgoB
signaling
chains
(see
Figure
2j.26).
A central dilemma
about
T cell function
remains
to be resolved.
Cell-mediated
immunity
requires
two
recognition processes.
Recogni-
tion
of the
foreign
antigen
requires
the ability
to respond to novel
structures.
Recognition
of
the MHC
protein
is
of course
restricted to
one
of those coded by the
genome,
but even so there
are many
different MHC
proteins.
Thus
consid-
erable diversity is required
in both
recognition
reactions. Helper
and killer T
cells rely upon
dif-
ferent classes of MHC
proteins;
however,
they
use the same
pool
of
cr and
B
gene
segments to
assemble their receptors.
Even
allowing for the
introduction of additional
variation
during the
TCR recombination
process,
it is not
clear how
enough different versions
of the T cell receptor
are made available
to accommodate
all these
demands.
V-region
of
TCR
recognizes
antigen
(
chain is efiector
i:ii:,iii:
l:
;;..:;r
The
two chains of the T ce[[ receotor asso-
ciate with the
potypeptides
ofthe CD3 compLex. The vari-
abte
regions
of the
TCR
are exposed on
the ce[[ surface.
The
cytoplasmic
domains
of the
(
chains of CD3
provide
the effector
function.
The Major
Histocom
patibitity
Locus
Codes
for Many Genes
of the
Immune System
r
The MHC locus codes for the ctass
I
and class
II
proteins
as we[[ as for other
proteins
of the
immune
system.
.
Ctass I
proteins
are the transplantation
antigens
that are responsible for distinguishing
"self'from
"nonself'
tissue.
.
An MHC
class I
protein
is active as a
heterodimer
wjth
Fz
microgtobu[in.
.
Class II oroteins are invotved
in interactions
between T ce[ts.
.
An MHC
class
II
protein
is
a
heterodimer of
g
and
B
chains.
The major histocompatibility
locus occupies
a
small segment of a
single chromosome
in the
mouse
(where
it is called
the H2 locus)
and in
man
(called
the HLA
locus). Within
this seg-
ment are many
genes
coding
for functions con-
cerned with the immune
response.
At individual
gene
Ioci whose
products
have been
identified,
many
alleles
have been
found in the
popula-
tion; the locus
is
described
as
highly
polymor-
phic,mearring
that individual
genomes
are likely
to be different
from
one
another.
Genes coding
for certain other functions
also are
located in
this
region.
Histocompatibility
antigens
are classified
into three types by
their immunological
prop-
erties. In addition, other
proteins
found on lym-
phocytes
and
macrophages
have a
related
structure and are
important
in
the
function of
cells of the
immune system:
MHC class
I
proteins are the transplan-
tation antigens.
They are
present
on every
cell
of
the mammal.
As their
name suggests,
these
proteins
are
responsible
for the
rejection
of foreign tissue, which
is
recognized as such
by virtue of
its
particular array of
transplanta-
tion antigens.
In the
immune system,
their
pres-
ence on target
cells
is required
for the
cell-mediated
response.
The types of class
I
pro-
teins are defined
serologically
(by
their anti-
genic properties).
The
murine class
I
genes
code
for the H2-I( and
H2-DlL
proteins. Each mouse
strain has one of several
possible
alleles
for each
of these
functions. The
human
class I functions
include
the classical
transplantation
antigens,
HLA-A. B. and C.
23.20 The Major Histocompatibitity
Locus Codes
for
Many
Genes
of the
Immune System
599

NIHC
class
II
proteins
are
found
on the
surfaces of both B
lymphocytes
and
T lympho-
cytes, as well as on macrophages. These
pro-
teins
are
involved in
communications between
cells that are necessary to execute the immune
response; in
particular,
they are required for
helper T
cell
function.
The murine class II
func-
tions
are defined
genetically
as I-A and I-E. The
human
class II region
(also
called HLA-D) is
arranged into four
subregions, DR, DQ,
DZIDO,
and DP.
The
complement
proteins
are coded by a
genetic
locus
that
is also known
as the S
region;
S stands for
serum,
indicating
that the
proteins
are components
of
the
serum.
Their role is
to
interact
with antibody-antigen complexes to
cause
the lysis of cells in the classical
pathway
of the humoral response.
The
Qa
and
Tla loci
proteins
are found on
murine hematopoietic
cells. They are known
as
differentiation antigens, because
each
is found
only on a
particular
subset of the blood cells,
presumably
related
to their function. They are
structurally
related to the class I H2
proteins,
and like
them are
polymorphic.
We can now relate
the types of
proteins
to
the
organization of the
genes
that code
for
them.
The
MHC region
was originally defined by
genetics
in
the mouse, where the classical H2
region
occupies 0.3 map units. Together with
the adjacent
region where mutations
affecting
immune function
are also found,
this corre-
sponds to a region
of
-2000
kb of DNA. The
MHC region has
been completely
sequenced
in
several mammals,
as well as in
some birds and
fish. By
comparing these
sequences, we find
that the organization has been
generally
conserved.
The
gene
organization
in mouse
and man
is
summarized
in
il{iti't!:
ii;-,:.1.
The
genomic
regions
where the class
I
and class II
genes
are
Iocated mark the original boundaries of the locus
(going
in the
direction
from telomere
to cen-
tromere;
right to left as shown in
the figure).
The
genes
in the region that separates the
class
I and
class
II
genes
code for a variety
of func-
tions; this
is
called the class
III region.
Defining
the ends of the
locus
varies with the species,
and the region beyond the class I
genes
on the
telomeric side
is
called the extended class I
region. Similarly, the region
beyond the class II
genes
on the centromeric side is
called the
extended class II region. The major
difference
between
mouse and human is
that the extended
class II region contains some class I
(H2-I()
genes
in the mouse.
There
are several
hundred
genes
in
the
MHC regions of mammals, but it is
possible
for
MHC functions to be
provided
by far fewer
genes,
as
in
the case of the chicken, where the
MHC region is 92 I(b and has only nine
genes.
As in comparisons of
other
gene
families, we
find
differences in the exact numbers
of
genes
devoted to each function. The
MHC locus shows
extensive variation between individuals,
and
as a
result
the
number
of
genes
may
be differ-
ent
in
different individuals. As a
general
rule,
however, a mouse
genome
has fewer
active H2
genes
than a
human
genome.
The
class II
genes
are unique to mammals
(except
for one sub-
group),
and as a rule, birds and
fish have
differ-
ent
genes
in their
place.
There
are
-8
functional
i':i.l:.i!il-
il.il.i,:;
The MHC region
extends for >2 Mb. MHC
proteins
of classes
I
and II are
coded bV
two
separate regions. The
class III region is
defined as the segment
between them. The extended
regions
describe
segments that
are syntenic on either
end of the ctuster. The major
difference
between mouse
and human is
the
presence
of H2
class
I
genes
in the
extended
region
on
the [eft.
The
murine
[ocus
is
[ocated
on chromosome 1.7, and the human
locus is located
on chromosome
6.
CHAPTER 23
Immune
Diversitv