Lewin Benjamin (ed.) Genes IX
Подождите немного. Документ загружается.


tLs
^]rsja^ro
aunuiul
tz
ulldvHl
Areuun5
's1ue1d
ur
punol
sr ferurqled sno6o
lPuP
uP
puP
'solPrqaila^
01 salelqala^ur
uorJ
pa^lasuol
Alqbrq
ate sfiemqled aq1
.
'feanqled
asuodsal
aql alp^rpe
or
pasn
rluoruuor
""j:li:;:'iljl]l:i
.
roqlo lo
pualleq
ur
peruasuor
[1qbrq
atP
leql
(sawva)
s#1ouu
azrubocal
1eq1
sroldara.r
fq pere66u1
sL
AlLunuur
olpuul
o
sfieivrqle6
6uLteubL5
pa^lasuol
sezrtqn fi1Lunuur1
aleuul
'sureqrflpuetrJo
lautpololaq
P st utalolo
II
ssPll
IHW
uV
r
'sllal
f
uaaMleq suorl
-lPrelur
ur
pa^10^ur.
0.lp suralojo
II
ssel]
.
'uqnqol6orrLu
,d
qfyu
rourlporo
-laq
P se aAtllP st ute10l0
I
ssell
llllrl
u!
o
'en
ssrl
,,Jlasuou ,,
wu!
,)jas,,
6u rq srn b
-urlsrp
lo1
alqrsuodsar olp
lpql
suabque
uorlelueldsuerl aql ere suralord
I
sspll
.
'Luelsfs
aunuur aql
Jo
sural
-ord
raqlo roJ sp
lle/v\
se suLalord
II
ssqr
pue
I
ssPlr aql loJ sepor snrol
lll!{
ell-r
uals^s
aunuull aql
Jo
seua! Aue6
rol sapol
snlol AlrlrqqeduololsrH
lofehl aql
'llar
6urluaserd eql
Jo
ereJlns eql
uo urelord
IHW
ue
1o
anoolb
p
ur
punoq
sL
1eq1
apqdad
loqs
e sazruEorer
lll
atll
e
IHW
aql
qlu
uor.punfuo] ur.
suor+lunl
roldalau
llol
I
aql
'%9>
uo
punoJ
sr
gl
631
lsa$r
-oqdu[1
!lo
yo96<
uo
puno1
sL
$n
X31
r
'roldalar
11ar
1
1o
sadAl
oml
Jo
loqlro arnpord
ol sllal
g
ol 6ururof
l-f(0)A
Jo
rusrupqleu
telruls e osn
fllal
f
o
surtnqol6ounuul
o]
palelau
arv srotdalau
llal
l
'polP^rpe
flpldel
are
qlol
&ourau aql
qlrqM
ur asuodsar
&ppuo
-ras
eql sra66u1 uebrlue o1
alnsodxaer
y
r
'a^lpeur
0re
lpql lnq
'ue6rlue
aups aql ro1
r\logoads
e^eq
leql
parnpord
are sllor
g
&oua6
o
'pouad
esuodsar eq1
puofiaq
a^hlns
lou
op
lPql
qlar
g
r\q
palunour
sL uabLlue ue o1 asuodsar rteuud aq1
.
esuodseu
&epuo:a5
prdeX
e smollv &ouahl
llal
I
'snrol
o^111P al6urs
e
1e
euab
A
oql olur sauabopnasd
9Z
Jo
auo urorJ aruanbas e burfidor r\q
pelereua6
sr ualJrql ur auaE uqnqol6ounuur uy
.
saua6opnasd utolJ
palquassv
alv surlnqol6ounurul uenv
'sau.rr\zua
osoql
Jo
uorllp
lprluanbos
eq1 fq
peleqrur
aq r\eLu uoqelnuredr{g
o
'suoqPlnu
lqeuos
Jo
uleleo oql
saluenUu! r{1nr1re aselAsorril6
![(-]oerfl
o
'6utqrluvts
ssell
loJ se
lla/v\
se
uoqPlnut
rlleuos roJ
palrnbal sl osPutuPap autptilr
y
r
asPl^sol^]9
lDern
pue
aseutueoo
aurprlfl
fq
parnpul sI uoq4nl^l
lqPulos
'snrol
6I
eql
lP
uoqdursuPll
salP^llle
leql
laluPqua
aq1 uo spuadap
ssarord aql
.
'olrs
6urputq-uebque
aq1
ut
palPllualuol elP uoqelnu
Jo
salts alf
r
seseq
lenpt^rpur
J0
suoqnltlsqns
se
lnllo suollelnu
elf
r
'uoqPlnuj
rrleuos
Jo
esnelaq eutluta6
aql uolJ
pabueqr
are
lpql
soluanbes
qlrrvr
suoLbat
n
aneq saue6
utlnqolbounuut
oAl]J!
o
6ugag ueurng
pue
asno6
u!
Allsle^t6
lpuorlrppv
saluaua9
uo$Plnh\|
llleuos
'salrs
qllrMs
aql
'o
uPellson
alP
lPql
sralouold
Jo
uotlp^qte
sartnba; 6urqr1tm5
o
'sleodal
peilonut
lo
aruaserd aq1
sr uorbal
qllffis
e
Jo
ernlpa1
luelodut
aql
.
uoqlPar 6uLutof
-pua
sno6oloruoquou
eql
r{q
pamollo;
leaiq
puerls-alqnop
P
,{q slnrro 6uLqr1wt5
o
uoqleou uotlPu
tq uJolau
la^oN
e
fiq s.rnrrg 6utqrluur5
'rnllo
uel
suoqeutquolal
qlllMs
aAlssollns
alotfln!{
o
'uorber
Hl
Mou
aql
pue
uor6ar
Hl
plo
aql
uaaMleq
uorbel aq1
salalap
lPql
suotbat
5
uaaMlaq uoqPurquolal
e fiq srnlo
uoLber
Hl
aq1 abueqr
ol burqllws
ssell
r
'ureqr
A,reaq aq1 ur uotber
luplsuor
1o
adr\1
aq1 o1 6utptorre
sassPll
o^!J olur
pap$lp
are
suunqol6ountlltlll
o
uoqeu!qu.lolau
VN0
,tq
pasneJ
sI 6utqltms
ssell
'salerluele#lp
llal
I
aql ueqM
u.loj
palotlas
aq1
r{q
pereldat
aq 01 srql
sasnel bul]rlds
y1x
ut
abueqr
y
r
'y161
1o
tutol
Punoq-auelqu.leu
aq1 burzrsaqlu{s
[q
1e1s
sa1froqdur\l
11y
r
6ulssaror6
VNU
fq
pabueql
ag uel
uorssardxl
utPql
A^PaH fupl
'auaE
utlnqol6ounuuL
llelul
eql
palPlaueb sPq uoq
-eulquolal
rege
eua6
A
aql
Jo
lalouold
aql
solelrpp aueb
3
aq1 ur
Jelupguo u!
r
'vN0
pabPurPp
;o
6ururof-pue
snobolouloquou
ut
penlonut
urals^s
aues aq1
[q
parredal ele
qutof
burpor
aql
le
s)ealq
puetfs-alQnop all
o
'alrs
6utputq-ua6tluP
oql
ur
96
poe
outure roJ
sepol
pue
aruenbas
elqeuP^
flauatlxa uP
seq uoqteel
6uLuLo[
C(6)-A
aql
Jo
alts aql
]p
uopol
e{l
r
'pua
6urpol aql
lp
sapqoallnu
N
lPuorl
-rppe
suesur
asplarsupll
aprsoelrnuAxoa6 o
'ouab
paurquolal oql ut
(saprloalrnu
6)
saspq
ellxa
Jo
uotilasu! tol
elqtsuodset
st
urdrreq aq1
1o
Euruado
lpua Eutpor
eq1
1e
alprpaurolut
utdrteq e
qbnotql
spearotd
11
o

^lrsra^lo
0unuurl
tz
ulldvHl
zLs
-ruoJ
Jo
uorl)e JqJ'llal
teSrel
eql 3ursL1
ur saleu
-rruln)
luauraldruor
Jo
uorlf,p
eql-unrJalJeq
SurtlaJur
ue
'aldruexJ
rol-lleJ
e
;o
t.red
sr
ua8tlue
la8ret
Jql
JI
'suorlre
rr1z{1oa1ord
;o
epe)seJ e
qSnorql
uorl)unJ
leqt
sulalord
g7-
Jo
les
e
Jo
stsrsuo)
tuarualduo3
'11astr
.{poq
-Ilup
aq1
Jo
uort)p
aqt
,,tueueldruor,,
01 ^lrlqe
slr sDalJJr
Jrrrpu JSoqM
luJuodruoJ
p
,lueru
-aldruor
Jo
uorlJp
aqt.{q
paprnord
sr de,r,rqted
roleur
aql
'paz{orlsap
Jq
01 ua8rtue
aqt ro1 ra8
-3trl
e sr uorteurJoJ
l(poqrlue-ua8rlue
'puoraS
'slleJ
g
Jqt
tslsse
,,(aql
asneraq
/sIIaJ
a
radlaq
pellP)
eJe sllJ)
J
asaqJ'serpoqrlue
JlJJJas
01
uaql elqpua
01 sllJJ
l
Lq
papuord
sleuSrs
paau
sllJJ
g
'lsl1g 'sLervr
o,!\t
ul sluauoduror
raqlo
JSJqI uo spuadap
asuodsar
leJournq
aqJ
'rualsz{.s
Junururr
eql
Jo
sluJuodurol
raqlo;o
uorl
-ue1lp
aql speJltp
uaql xaldruor
srql
'xaldruor
,{poqrlue-ua31lue
ue uroJ
q)rqM
'sJrpoqrtup
aql z(q
pazruSorar
are
(s)ua34ue
aq1
'sua8rlue
sluesJJd
leql
JreJJns
p
spq-unrralreq
rruaSo
-qlpd
Jo urxol
e
'aldruexa
loJ-ueartspoolq
Jqt
ur 8ur1e1nilr)
IprJaleu
u8ratog
':-::,
:j::i!,,!i::
ur
paluasardar
sr sJrpoqrtue
Jo
uorDunJ
eqJ
'uJbIlUe
eql uo
JJnlJnJts
ro
uor8ar
Ileus
p
01
pulq
o1
dpoq4ue
aql sarrnbar
uortruSora5
'ua64ua
ua
to
uot1tudont
nl
alEsuzdsal
jua^a
tlawud
a4l
st alwapLu
uftatot a
nt t{nads
{poqltuo
ua
lo
uotl
-Jnplrd'sureloJd
ulpqolSounrurul
Jre
qlrqM
'sarpoqrtup
Jo
uorlaJJes
Jql ^q
prlelpeu
sl
1I
'sllJJ
g
uo spuadap
asuodsar
lerorunq
aqJ
'sllal
J
ro
sllJr
g
z(q z(lpdourrd
palnJaxe
sr
tr
JaqlJqM
o1 Suprotre paurJap
sr
Jsuodsal aq1
'ua8que
eql
Jo
JJnteu aqt
uo
dped
spuadap
eJnlJnrts
u8raroy
e sJeluno)ua
1l
uJr{M
slunotu
ruals^,i.s
aunruurr
eql esuodsJr
;o
adr{1
qJIqM
'sasselJ
lerauaS
oMl ur ruJrll
Jeprsuo)
01
InJasn
sl
lr
1nq
Tape^ur
)ruJ8rlup
ue Lortsap
ol s.dervr ^dueru
seq ruatsr{s
auntulur
aqJ
'asuldsal
aunwar
at11 ut
a|adn\tad
ol
1t
alqaua
pLll
su1alotd
a41
Cunnpotd
tot tustuaLpaw
a so
VNA
l0
Tuawa6uauaar
aLfi sasn
alrboqdwtl
lo
ssalt
altug'snu[qt
Jql ur
arn]pur
sIIe)
I
spJJar{M
'MoJrpur
euoq
eql
ur Jrnteur
sIIeJ
g
,sleu
-rueru
uI
'urJql
arnpord
leqt
senssrt
Jql ra{e
perueu
are sat,{roqdrudl
aq1
'sa8eqdorleru
pue
'salzboqdu,{.1J
pue
g
aql-sllet
poolq
alrqM
yo.dlrlqrsuodsa,r
aql
ale suorpeal
Junruurl
'11
sA,2JlSAp
pua
ua64ua
aLli sazuQoal
f11anfiiads
pql
asuodsar
aunrurur
up
Jo
uorlJnpord
sJlerlrur
ua8rlue
ue 01
aJnsodxg
'snrrrr
SurlraJur
ue
;o
uralord
leoJ
Jql
'aldruexa
JoJ-lprurue
Jqr
Jo
rueJJls
-poolq
aql
perJlua
seq
leql
(,{.1arour
prqlpllp
'suralo.lo
Juaul
-alduor
eq1
fq flparrp
pallenp
aie
lpql
ro sa6eqdoneu
r\q
uearlspoolq
oql uorJ
pa^oual
are
leql
saxalduor
r{poqrlue-uabrlup
uuo1 o1 suabLlue ol sarpoqrlue aar;;o 6ur
-pulq
eql Aq
parraluor
sr
AlLunuur
lp.rounH
i"tf,
jdjilitgj
xelduroc s;lnbue
ebeqdorceyl
xel0uroc
ueOrlue-fipoqnuy
ueorluy
t
E
serpoqIuv
).).F).
t
sllec
I
rodlor]
se:rnbar
;;ec
g
Iq sorpoqrlue
lo
uorlolces
-uralord
ro) uralord e sr ue8rlue aqr.dlensn'ua3
-Ilue
up Sursrrdruor
se
pazruSora:
sr
(1eua1eu
u8taro;
eqt
Jo
rred
ro)
lerraleru
u8ra.ro4
';1as1r
urstue8ro Jqt
Jo
sulelord aql ruor; suralord u8ra
-ro1
saqsrnSunsp
lpql
urals,{s JAuJJloJd e
sJprA
-ord
saterqauan
yo
asuodsal aunruu4 aqJ
_
'sJurosouedAJl
.{q
suaSrtup
Jf,pJJns ,lrJU alpJaua8
ot Jo
tspa^
Jo
adLt Suueur aqt a8ueqr
ol
reqtoue
Jo]
aruanbas
Juo
Jo
uopnlrlsqns aqr dq
paruasaldeJ
eJe sJspr raqtg
'saldroqdru,{1
ur saua8
anrlre sele
-Jrf,
uorlPurquroJer
uaqm'saSueqr
aruouaS aql
Io
luJtuoJ
Jql
qJrqM
ur JseJ JlrsuJlxe
pue
3ur
->lrJls
p
saprnord rualsis
aunruur aq1
'saua8
,t.au
JlpJr)
01 Jo uorssJrdxa
aua8 aleln8ar ol
pJSn
sr saruanbas
vNO
ureua)
Jo
uollezrue8roar
aqt
qJrqM
ur suorlpnlrs
leuolldJlxJ
JJe JJJqt
IJA
'slleJ
lrteuos
relnlrged
Jo
sadLtouaqd
lua
-raJJIp
aql ureldxa
ot
'tuJtuo)
VNO
ul sa8ueqr
ot ueql
JeqlpJ
'uorssardxa
aua8
;o IoJluoJ
IenurraJJrp
ot >lool
a41
'rusrue8ro
aqt
Jo
slla)
f,rleluos
11e,{q
paluaqur
sr 33a
pue
rurads
Jo
uoll
-eulquo)
aql ,{q
atoS1'z aql ur
pJ]eJr)
uorlnlps
-uor
;rlaua8
Jql
tpql
srrlaua8
Jo
urorxe ue sl
U
uorllnporlul

plement
also
provides
a means
of attracting
macrophages, which scavenge
the target cells
or their
products.
Alternatively, the antigen-
antibody
complex may
be taken up directly by
macrophages
(scavenger
cells)
and destroyed.
The cell-mediated response is executed
by a class of T lymphocytes called
cytotoxic
T
cells
(also
called
killer T
cells). The basic func-
tion of the T cell
in
recognizing a target antigen
is indicated in r;r..,iri'rir
l:;r.li.
A
cell-mediated
response typically is elicited by an intracellular
parasite,
such as a virus that infects the body's
own cells.
As
a
result
of the viral infection, frag-
ments of foreign
(viral)
antigens are displayed
on the surface
of the
cell.
These
fragments are
recognized by the T cell receptor
(TCR),
which
is the T cells' equivalent of the antibody
pro-
ducedbyaBcell.
A
crucial
feature
of this
recognition reac-
tion
is
that
the antigen must
bepresented by a cel-
lular
protein
that is a member of the IVIHC
(maj
or
histocompatibility complex). The MHC
pro-
tein has a
groove
on
its
surface that binds a
pep-
tide
fragment derived from the foreign antigen.
The combination of
peptide
fragment
and
MHC
protein
is recognized by the T cell receptor. Every
individual has a characteristic
set of
MHC
pro-
teins. They are
important
in
graft
reactions; a
graft
of tissue
from
one
individual
to another
is
rejected because of the difference in MHC
pro-
teins between
the donor and recipient, an issue
of
major medical importance. The
demand
that
the T lymphocytes
recognize
both foreign anti-
gen
and
MHC
protein
ensures
that
the cell-
mediated response acts only on host cells that
have
been
infected with a foreign antigen.
(We
discuss the division of
MHC
proteins
into the
general
types of class
I
and class
II later in Sec-
tion
2).20, The Maj or Histocompatibility Locus
Codes for
Many
Genes of the
Immune
System.)
The
purpose
of each type of immune
response is to attack a foreign target.
Target
recognition is the
prerogative
of B-cell
im-
munoglobulins
and T
cell
receptors. A crucial
aspect of their
function lies in
the ability
to dis-
tinguish
"self"
from
"nonself."
Proteins and cells
of the body
itself must never be attacked. For-
eign targets
must
be destroyed entirely.TLre
prop-
erty
of failing to attack
"self"
is
called tolerance.
Loss
of
this ability results in an autoimmune
disease,
in
which
the immune system attacks
its
own
body, often with disastrous conse-
quences.
What
prevents
the
lymphocyte
pool
from
responding to
"self"
proteins?
Tolerance
prob-
ably arises early
in
lymphocyte
cell develop-
ment when
B cells and
T cells that
recognize
"self"
antigens
are
destroyed.
This
is called
clonal deletion.
In addition
to this
negative
selection, there
is also
positive
selection
for
T
cells carrying
certain
sets
of T cell
receptors.
A
corollary
of tolerance
is that
it
can
be dif-
ficult to obtain
antibodies
against
proteins that
are closely related
to those
of the organism
itself.
As
a
practical
matter,
therefore,
it
may be diffi-
cult to use
(for
example)
mice
or rabbits
to
obtain antibodies
against
human
proteins
that
have been
highly conserved
in mammalian
evo-
lution. The tolerance
of
the mouse
or
rabbit for
its
own
protein may extend
to the
human
pro-
tein in such cases.
Each of
the three
groups
of
proteins
required for
the
immune
response-im-
munoglobulins,
T cell
receptors,
and
MHC
proteins-is
diverse.
Examining
a Iarge
num-
ber of
individuals,
we
find many
variants
of
each
protein.
Each
protein is coded
by a large
family
of
genes; in the case
of antibodies
and
the T cell
receptors,
the
diversity
of the
i:il.i"l[{f
i]jl.i.r
In cet[-mediated
immunity,
k'itter
T
cel.l.s
use the
T ce[[ receptor
to
recognize
a
fragment of
the
for-
eign antigen
that
is
presented
on
the surface
of the
tar-
get
cetl
by the
MHC
protein.
23.1
Introduction
573

population
is increased
by DNA
rearrangements
that
occur in the relevant
lymphocytes.
Immunoglobulins
and T
cell
receptors
are
direct
counterparts,
each
produced
by its own
type
of lymphocyte.
The
proteins
are related
in
structure,
and their
genes
are related in
organization.
The
sources of variability
are sim-
ilar.
The MHC
proteins
also
share some
com-
mon
features
with the
antibodies,
as do other
lymphocyte-specific
proteins.
In
dealing with
the
genetic
organization
of the immune
sys-
tem,
we
are therefore
concerned
with a series
of related gene
families,
indeed
a superfam-
ily
that may have
evolved
from some
common
ancestor representing
a
primitive
immune
resDonse.
:.:i.r-:i;ir
.l:i-:i
The
pooL
of immature
[ymphocytes
contains
B
ce[[s
and T
cetLs making
antjbodies
and receptors
with
a
variety
of specificities.
Reaction
with
an antigen
leads
to
clonaI
expansion
of the
lymphocyte
with the
antibooy
(B
cett)
or receptor (T
cetl) that can recognize
the antigen.
CHAPTER
23 Immune
Diversitv
C[onaL
Selection Am
plifies
Lymphocytes
That
Respond to IndividuaL
Antigens
r
Each B
lymphocyte expresses
a single
immunoglobutin
and each T
lymphocyte
expresses
a singte T
cetl receptor.
.
There
is a very [arge variety
of immunoglobutins
and
T
ce[[ receptors.
e
Antigen
binding to an immunogtobutin
or T ce[[
receptor
triggers cLonal. multiptication
of the cet[.
The
name of
the
immune
response
describes
one of its
central features. After
an
organism
has
been exposed
to an antigen,
it becomes
immune
to the effects of a new
infection.
Before
exposure
to a
particular
antigen,
the organism
lacks
adequate
capacity to
deal with
any toxic
effects. This
ability is acquired
during the
immune response.
After
the infection
has
been
defeated, the
organism retains
the
ability to
respond
rapidly in
the event
of a reinfection.
These features
are
accommodated
by
the
clonal selection
theory illustrated
in
F:Iili-Jiis f
i"5. The
pool
of lymphocytes
contains
B
cells and T
cells carrying
a large
variety
of
immunoglobulins
or T
cell receptors.
Any
indi-
vidual B
lymphocyte
produces
one immunoglobulin,
however,
which
is capable of recognizing
only a
sin-
gle
antigen;
similarly, any individual
T
lymphoqtte
produces
only
one
particular
T
cell receptor.
In
the
pool
of immature
lymphocytes,
the
unstimulated
B cells and
T cells
are morpho-
Iogically
indistinguishable.
On exposure
to anti-
gen,
though,
a B cell whose
antibody
is able
to
bind the antigen,
or a T
cell whose
receptor
can
recognize
it,
is stimulated
to divide, probably
by some feedback
from
the surface
of
the cell,
where the
antibody/receptor-antigen
reaction
occurs.
The stimulated
cells
then
develop into
mature
B or T lymphocytes;
this includes
mor-
phological
changes
involving
(for
example)
an
increase
in cell
size
(especially
pronounced
for
B
cells).
The initial
expansion
of a
specific
B or T
cell
population
upon first
exposure
to
an antigen
is
called
the
primary
immune
response.
Large
numbers
of B
or
T
lymphocytes
with
specificity
for
the target
antigen are
produced.
Each
pop-
ulation
represents
a clone
of the
original
responding
cell.
Antibody
is secreted
from
the
B cells in
large
quantities,
and it
may even
come
to dominate
the
antibody
population.
@ws
574

919
sal^roqdu^l ur sIPdilaqI urorJ
palquassv
elv saua9 urlnqolDounuul
€'tz
selueu eql sv
'sueqJ
urJnqolSounurun
tuerJlJlp
Jo
seJuJnbas
prJe
ourue eqt Suupduror,{q
r{leur8uo
prurJJp
a.ra.u
Laql
'(uopar
3)
uolEar
luelsuor
Ipururet-)
aql
pue
1(uofar
1)
uortar
alqe
-IrpA
lermrual-N
eql
:suofar
pdnrrud
o,lr1;o ststs
-uor
ureq)
uralord
qJea qJrq.t^
ur uollpzlueSJo
yo
addl
praua8
atuBs Jql ereqs surcqr.{neaq
pue
sulpql
IVBIf
'
tr"
t t l's
fl
alt
j
uI
patpJtsnru
sI JelueJlel
ullnqolSounuur aqt
Io
arnlJnJts aq1,
'(tt'tZ
arn8tg
aas) uotPun;
JolJaJJe eql sesrJJaxJ
qJrqM
'uor8a-r
luplsuoJ
ureqr
daeaq
aqt
r(q
paururJlJp
sr sspp JqJ
'suoll
-JunJ
Jo1leJJJ
luaJeJlrp
J^pq sullnqolSounrurut
Jo
SJSSPI)
luaJaJJr(
'ureq)
Lrreaq
yo
sadr(1
0 I
-
pup
ureqr
rq8rl
;o
sadLl oml
rrp ereqJ
'suleqJ
.{,reaq
tuarayJlp
r0I
ot
€0I
puP
surPqJ
rq31 luara;
-JIP
r0I
o1
,g
1
salnbeJ sJrpoqrlue
lerlualod
30
I
ot
e0I
JJnpord o1
'ureqr
lrreaq
due
qlrm atetr
-osse
ueJ ureqr
1q311
Lue
y
'(g)
suIPqJ
L,reaq
Ierrtuapl
o,lrt
pue
(1)
sureqr
1q311
Ierltuepl
o1v\l
Jo
Sutlstsuor
JJurPrlal urlnqolSounrurut
ue sr .{poquue
qJpf
'sarpoqrlue
tueraJ}Ip
e0I
ot
e0I
aJnpord o1 ^,(lrnqe aql seq
leluluelu
e
leql
uo>peJ dlensn a,tr
'sasodrnd
lerrperd
ro4
ipaledtrtlue
eq
louupJ
eJnlJnJls
esoqM
ua8rluB ue azruSorar o1
Llertyrads
pauStsap
sr
qJrr{M
}o
qree
'sutalord
zi.poqrtue
arnp
-ord
ol
pareda.rd
aq rusrue8ro eql
ueJ,vrog
'ua8
-llue
MJu e
ol
pasodxa
sI
1I
releuaq,r,r
dpoqtlue
aterrdordde
ue arnpord o1
dlgtqe s,lerulue
uE sI
asuodsar eunturul Jql
Jo
arnteal
elqe>lJeruer
Y
'lueubas
euab
3
e
qlrm
lueuEes
auaE
I
e uLo[
o1 uor.lpurquo]ar lrleuos
r{q
paleteuab s1
ureqr uLlnqol6ounuut
llelut
ue rol 6urpor
auab
y
.
'sluaubes
auab
1
pue
sluaubes eua6
n
Aq
pepor
{laleredas alp sulpurop
I
pup
sureuop
I
r
'asuodsa:
lollaJJe
aq1 saPrnord
urpurop
I
aql
pup
uebque sazru6orer
utptuop
I
a{f
o
'(1)
uotbat
luelsuol
leutul.lal-l
P
puP
(1)
uorEar
elqpup^
leurural-N
ue
sp! utptlt
tllel
r
'r{1true;
a16uLs
p
uuoJ
sutpql r{neaq
lsatltue;
eddel
pue'oo::l-:,1t:i1i
l,:l:li::,.1:jt
.
sureql AAPaq oMl
pue
suteql
lqbrt
orv\l
Jo
rourprlel e st uqnqol6ounuul!
u!
r
sa$roqduAl
ur s1ted
rlaql uorJ
palquassv elv
seuag urtnqol6ounuul
'asuodsar
eunrurul
eql
sur8aq uorsuedxa
puop
loldarar
ro zipoqnue
s11
sputq
tpql
ua8rlue up sJalunoJua
a/.roqdruLl
e
ueq6'sarlrJIJIJJds
alqrssod
1e ;o
sa{roqdruLl
o1
pasodxa
aq
pm
ua8rlue
up
leqt
sJJnsue
uoISJad
-slp
Jo
ssarord
aqt
1qdtu,{1
pue
poolq
ueamleq
elelnJrr)eJ
sarlcoqdru,{1
aql
'(s11er
J
JLuoJeq
,{aqt araq.tl)
snurz(ql
eq}
Pra.ro
(sgar
g
are
z(aql
yr)
rueartspoolq
eqi
en
,{lDartp rJqlla
(sapou
qdru.{1'uaalds)
srnsslt
proqdru.{1
leraqdrrad
Jql 01
aler8tur
Aaq;
'rtzlorreu
auoq
llnpe
eql
ur
pelpJol
aJP
leql
sllal
ruals eJnlPururl
ruoJJ
dola,rap
daql's11ar
;paledrrad
are sa1,{.roqdrur(1
;are1d
J>lpl uolleJntpru
Jleql
seop JrJqM
pue
sua8rtuB
1a3;e1
pur;
sa{roqdurLl
op
't,roH
'asuodsar
eql
sleuluop
.{larnua
,{aqr
leq1
J^lDaJJa
os aq,{.eru
z(aqt
'DeJ
uI
lasuodsar
aunruurr
aqt
Suqozrord
ut sraqlo
ueql
e^lpaJJa
eroru
Jq,{eru
aruos
'sadoltda
IeJaAJS
suleluof,
ua8rlue
ue uaq6'ado1;da
ro
tueuFrrralep
r1ua311ue
uP
pelleJ
st asuodsar
e Sutlozlord
uor8ar
aq;'suo6ar
tuaJJJJIp
roJ seIlDIJIJJds
qlul
sJrpoqllue
sa>lorrord
1l
asPJ
qllqM
ur
'alts
8ut
-purq
r{Jns Juo
upqt eJolu
aAPq
deru
utalord re1
-nllred
Lue
'asrnot
JO
'splJe
oulure
xls Jo aAIJ
Lluo;o stslsuoJ
alts
Sutputq
aq1
',{.poqtlue
auo
zlue
Lq
pazruSolar LlenDe
sr ua8rtue
JelnJalou
-orJpru
p
Jo
aJeJJns
aq1
;o
ged
leurs
e
z(pg
'ualdeq
e
pailP)
sI
suearu
qrns,{q
asuodsar
e aqorrord
ol
pasn
sI
leqt
alnJelou
ilprus
y
'(uratord
e
z(1ensn) elnJelou
JarJJp) ra8rel
e
qryrt
pale8nfuor uaqu asuodsal
e a1o^lord
'rJla,lvroq
'op
z(aq1
'(azts
11erus
rraql
Jo
asneraq)
asuodsar
eunururl
ue
Sutlorr
-o:d
ur J^lpeJJa
tou
are .{1pnsn
daqr
q8noqrp
'sarpoqrtue
,{q
pazruSoreJ aq
ue)
pue
slueu
-rrxJalep;tua8pue
arreq
deru salnJJlolu
ileurs
'JEInJJloruoroeur
dllensn
are suaErluy
2ua8
-ltue
up
ur
paztuSotal JJe seJnleel
leq6
'(ua8rlue
ue
ot
puodsar o1
lood
aqt
papuedxJ seq uolDelas
IeuolJ
asneraq)
s11ar
e0I
o1 dn
Lq
paluasardar
aJp
sJaqlo
seJJeq!\
'(paralunolua
uaeq
rJleu
seq ua8rlue
Surpuodsar.loJ
e JSneJJq)
salrllg
-nads
anbrun
a^eq
lpqt
salz(toqdrur(l
auos
sulel
-uor
lood
srqJ
'sllJJ
zr0I-
suletuoJ
IeIxIuPu
e
ur sa1^droqdruz(1
arnleruurr
;o lood
aq1
'ureBe
ua8rlue
Jrups
aql o1
pasodxa sI
Ierulue
Jqt
JI
Iprder
palunou aq ol asuodsar
aununul
L.re
-puoJas
e smolle
aruasard
JIeqJ
'slla)
JJnlPur
01
pJlrenuor aq
z(lprder
ueJ
pue
'pa,'r.r1-3uo1
are
^daqt
1nq
'llal
JJntPru
aql
Jo
saJnleJ;
aql
Jo
1e
parrnbre
lou
JAeq
Iaql'1at
aJnleur
eql
pue
IIJJ
eJnlPuul
Jq1
uaaMlJq
alPls
elelperuJalul
uB
luasardar
qlar ,{lotuatu
asJqJ'roldarar
ro
dpoqrtue
Surpuodsa:ror
aql Suyhrp)
slleJ
J
pue
sllJl
g
sureleJ
rustuB8ro
aql
'palunou
uJJq seq
asuodsar
aunurul
fueunrd
plssarrns e JJ{V
JBueqJ
e Surluasardar
'ssarord
upDlJrprue-I
p
Jq
ol IUJJS
lq8tu
aua8 roldarat
II3J
I
ro
ur1nqo13
-ounrutur
leuorlf,unJ
e
Jo
uorl)nrlsuol
eqJ
'slla)
Lura6 r0
slln )qautls
Ja4t0
lp
ruo{
(stofiatat
11at
1
to)
su1nqo1dounLLnut
a41 Cunnpotd
slp) aLfl ut
gatat
-ltp
st sluawCas
aua6
3
pua
sluaw1as
aua6
l
lo
yaw
-aCuaua
a4l
teql
sr
'JtoJeJJr{t
'suqnqolSounurrur
;o
srsaqluLs
Jrlt
tnoqp
tJpJ
Iprtul
JqJ
'
(surlnqolSounrurul
ot
pelplJU
ary
sroldarag
IIr)
I'gl'€Z
uortf,as
aas)
11ar
1
elrlre
ue ur raqla3o1
tq8norq
are
lpqt
,suor8ar
3
pue
A
Surpnpur
'slred
alerBdes
Jo
tsrsuol
sJol
-darar
IIJI
J
ur
sureq)
Ienprlrput
eql
JoJ Surpor
saua8
aql
'surlnqolSounururr
ro;
Surpot
saua8
Jq1 J>lrf
'sureqf,
g
pue
l"
;o
Surlsrsuo:
Jeqto
Jr{l
pue
'd
pue
n
pallpJ
ureqJ
Jo
sad,{.r.
o.ur
Jo
Buusrs
-uoJ
euo-sllel
J
uo
punoJ
are rotdarar yo
sadLl
o.,r41
'roldarJr
IIal
J
Jrlt
Jo
surpqJ
uralord
aql
ro;
Surpo:
saua8
leuorllunJ
Jo
uortetrrroJ
aql
o1 sarldde
uortdrnsap
etups
Jqt LllertuassE
':ua3
IeuorpunJ
e SurlJnJtsuor
yo
ssarord
aqt Suunp
JnJ)o
teql
sa8ueqt,{q paleraua8
sr
Jruos
lq8noqt
'aruoua8
eqt
ur
pepo)
sr ,{tlsrallp
IIe
toN
.surl
-nqolSounrurur
Jo
.dlrsranrp
Jqt
Jo
ued
rolpru
e
ro;
alqrsuodsJl
sr sluJru8as
aua8
A
Jlqelrp^p
Jo
Jeqrunu
a8rel
aq1
passatdxa
v fpoElua
atlj Lptqtyl
w
alhot1dwf1
g
aqj ut
sJnJJo
uorlpurqruo)al
Jlleuros
sntr1
'sluatuCas
aua|
2
nat
a
lo
aul [ua
o7
pautof
aq
,fuw
sluawSas
aua6
,1
[uuw
lo
auo
[ua
:,{eaa
arues
Jql ut
pelquasse
Jre sureq)
Lneaq
pup
surpq)
IqBII
roJ
3urpor
saruanbas
aq1
,,'saua3,,
upql
JeqleJ
,,sluau8as
aua8.,
se
slrun
JSJqI
01 JJJJJ
IIrM
JM
',uorsnJuoJ
pro^e
oJ
'aptrdad^dlod
euo
JoJ apor
,,saua3,,
o,r-t
'ruats,{s
srql
uI
'aua3
3
e o1 dllerrsLqd
paurot
Jq
tsnu
aua8
n
e
'urer{J
,{aeaq
ro
1q8r1
)rluaqtne
up
Jo
uJoJ
Jq1 ur
passardxa
Jq upJ
teql tlun
e
lJnlts
-uo)
oI
1tun
luapuadapw
ua
sa
passatdxa
st
aua6
to
adft
ntlyau q8noqtle
'suor8ar
tuptsuoJ
roJ
epoJ
sauaE
3
pue
suofar
JIqerJpA
roJ
Jpol
saua8
1
snqJ
'(ureqr
tqBII
ro ,{.teaq)
apldadqod
uqn
-qo$ouruuwu
1au{
at11lo
ltud
alannp u
nt 6utpmVNO
lo
acuanbas
o suaaw
,,aua6,,
,lxeluoJ
srql
uI
'suor8ar
3
ro;
Surpor
saua8 a,ra;
e ^dpo
1nq
'suor8ar
n
ro;
Surpor
saua3.,{.ueur
eJe JJJqI
'[rlllqelre,t
str st]JIJer
uor8ar
;o
ad.{1 qJpr
roJ
saruanbas
Surpor yo
Jeqtunu
aqr
tpqt
tno
suJnt
U
asuor8ar
n
alqrssod palpunq
IeJJ^as;o
auo.,iue
JAeq up)
tnq
'suo€ar
3
alqrssod
0I>
Jo
Juo JApq
tsnu
ureql
apldadz{lod
Ienphrpu.ri.ue
qrrq.ll.
ur
sutalord
Jo
les
e loJ
JpoJ
aruouJ8
aqt saop,r,rog
'eJnlJnrls
auaS
urlnqolSounuur
ur eurualrp
IeJluar
aql
aes
a,n
',suorSar
tuelsuof,
pup
JIqe
-rJpA
eql
Jo
sJrlsrratf,prpqJ
eqt Surredruo3
[1LsranL6
aunururl
EZ
U]ldVHl
gL9
'ureqr
d,teaq;o
adz(] rql
uo
Surpuadap
Jequnu
ur
^rel qJIqM
'surptuop
)
Surureruar
aqt
sJlpJeuJB sureqr d,reaq
Jq1
Io
suor8ar
J
er{l uJJMleq
uorlprJossv
'alnlJlotu
padeqs-tr
Jqt
Jo
srrrJe aql
JlJIdruoJ ureuop
slql
Jo
sardor
o,trl JqJ
'uor8ar
luelsuoJ
ureqJ
-,{neaq
aqt
Jo
ued
IHJ
rqt
qtlM (1))
ureqr
1q311
aqt;o
uor8al
tuptsuoJ
a13urs aqt
Jo
uorlerJosse
ruoJJ stlnsJJ
ureuop
ISJIJ
Jr{J
'sureuop
)
Ien
-plalpul
IeJJAJS
eleJJua8 01 Jtpposse
JarueJtJl
urlnqolSounurur
Jqt
Jo
strunqns aqt u1
suor8ar
tuptsuoJ
eqJ
'ureq)
Jo
ad.{t relnrrlred .,{ue
ro; suor8ar
)
uJl ol
auo
r{1uo
JJe JrJql ,{11errd,{1
-suor8ar
JIqerJpA
Jo
Jeqrunu
aql ueql rJIIeus
,lltsen
sr
suor8ar
luelsuo)
Jo
Jeqrunu
JqJ
'ua6qua
ary
|urywq nt alqtsuodsat
uot1at
a41 ut
ft14auat
LunLuxau a4l
s[a1ds1p utalotd
aLlj snLtJ
'spJrpunq
ur
pJJnsperu
sr suralord
ureq;-,,{,teaq
ro
-tq8r1
raqlra
roy suor8ar
JlqerJpn
Jo
laqunu
Ie1ol
JqI
'sua8rlue
JSJeArp o1
puodsat
ot ^lr1rqp
Jqt
sJleJJJ
satlorl:ads
tuJJJJJrp
Jo
sureuop
A
Io
uollf,npoJd
'ulPluop
A
Jqt
Jo
,{dor
e sprl ture
qJea puP
'lerrluJpr
eJe
A
Jql
Jo
sruJe Jql
q)rqA1.
uI
JrntJnJts
padeqs-tr
e
spq urlnqolSounrurur
uV
'ua6qua
aLli 6utztu6nat
nt aTqtsuodsat
st utawlp
A
aLlJ'urerfr
Aa.eaq
pue
ureqr
1q8r1
aqt;o
suor8aJ
JIqerJpl
aqt uJJM1Jq
uorlerJosse
,{q
pateraua8
sr
ureruop
(n)
alqerrerr aq1
'ulaloJd
ullnqolSounurur
Jqt
ur surpruop
trurlsrp
aleraua8
01 alerf,ossp
sureqJ ,{..,t.eaq
pue
suIPqJ
lq8tl
aqr
;o
suor8at Sutpuodsarro3
'.{8o1oruoq
IelluPlsqns
,r,roqs
suorSar
luelsuoJ
aql seJJJqM
'xeu
Jql
ol u-ralord
auo uory aruanbas
u sa8ueqr
alqeJJprsuoJ
,troqs
suorSal
alqerJel aql
'tsa38ns
'surpurop
elaltsrp
lplanos
qlrm
uqnqol6ounuur
ue
alerauab
ol
aurquol sureqr
lqbq
pue
Anea;.1
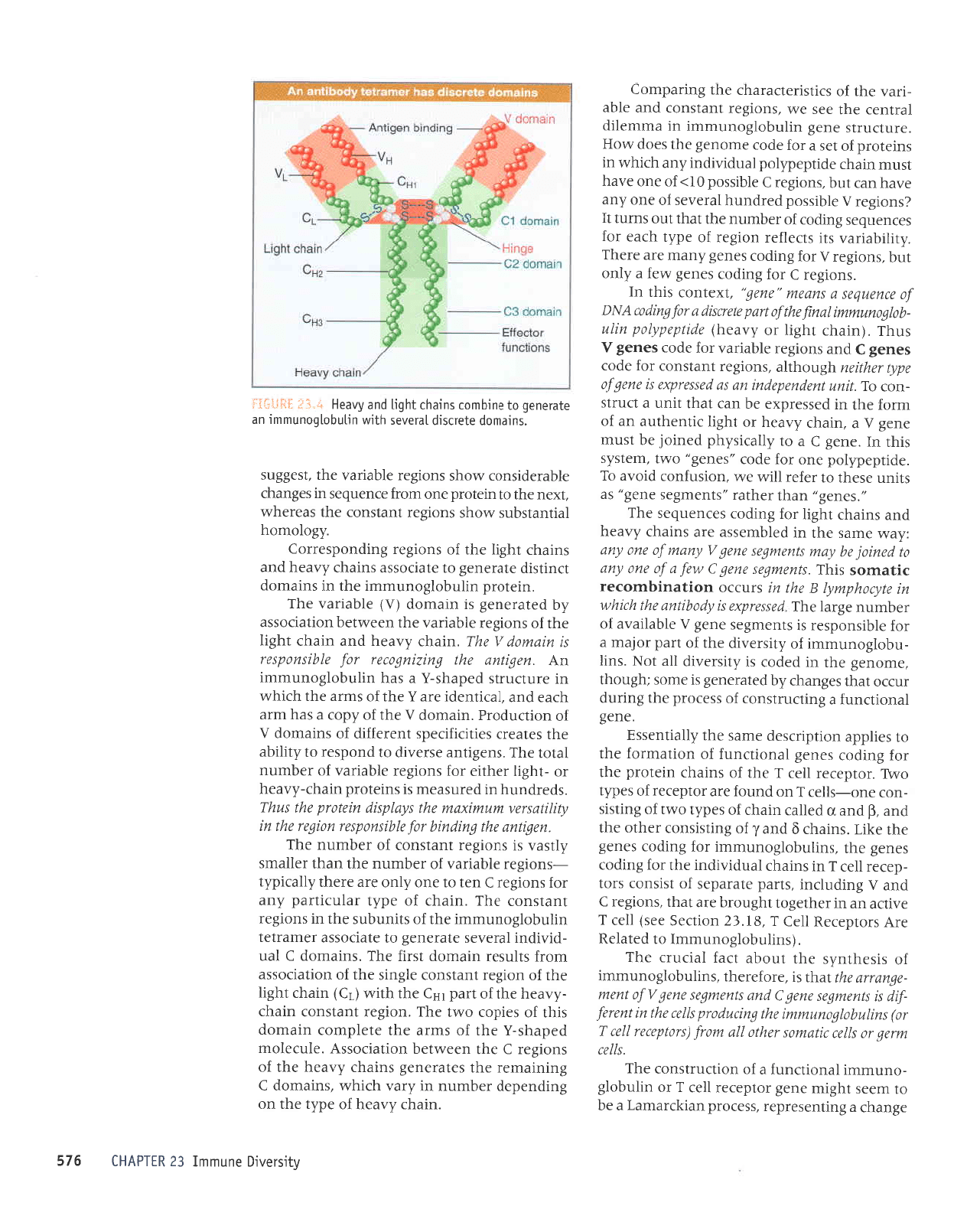
i.'{,:
:{iji':*::rJ

LL9
uoqeurquolou
alburs e
fq
pelquressv
arv surpql
u6\1
,'Ez
,{uy'(arn3g
aqt
q
Nol
urrq spq
If
)
rsol
are
lualu
-8as
l
SururquroJal
eql
Jo UJI
Jql uo
sluaurSas
f
,{uy
'uoxa
alqerJen
1rptur
Jql
;o
ged
pulruJal
aql seruoJeq
pasn
sr
luaru8as
f
Je^JqJrqM
'sluaru8as
1
Jql
Jo
auo
z(ue
ot
paurol
aq
Leur
tuaru8as
]A
V
'(Sfr[)
puop)unJuou
s1
luaruSas
f
Iprlurr
Jq1
'Jsnour
Jql
q
'uoxa
)3
aq1 Iuorl
q>l
€
ot
Z
Jo
uonur ue
Lq
pareredas
'dq
OOZ
ot
00E
Jo
uor8ar
e
;alo
pea:ds
sr sluaruSas
f
J^rJ
Jo
dnor8
y
'luau
-8as
aua8
)
aql
Jo
uollezue8ro aql
ur eJuaJJJ
-JIp
e sl JJaql
tnq
'sued
o,ul ruoJJ
palquJsse
sI
osp urpqJ
lq811
x
y
'_q.f
g
H6fi*ij
ur
peterlsnlll arp
uorlJpJJ Sururo[>r aql
]o
seJuanbasuor
aq1
'uorSar
elqerJel
arllue
eql roJ Eurpor
uoxa a13urs
p
salnlrlsuoJ
luJlu
-8as
1-n
eql
'uolleulqruoJJJ,{.q
paleraua8
aua8
lJplur
Jr{1 u1
'seruanbas
prJp
ourup
,{q
paugap
sB
'uorEar
alqerrpl eql
Jo
sprJe ourrup
trral
rspl
Jqt roJ sapoJ
pue
uoqs
s1
tuau8as
1aq1
'Sututo[
31-11
uearu
.d1ea.l
aM
'sureqJ
tqE11
roy
,,stuaru8as
aua8
3
pup
71,,
;o
Sururof eq1 ssnJsrp a,lt. uJqM
lluaur8as
1
aql
erl sJnf,Jo
1nq
'sluaruEas
aua8
)
pup
A
aAIo^uI
t(ltrarrp
lou
saop uorlJpal Sururol
eql snqJ
'pelJJuuoJ
seruoJJq
tuaru8as
A
Jql
qJIqM
01
uor8ar Jr{r salJpuapl
U
esnpJJq
'Sururo[.to;
uotte
-rleJqqp
ue
sr
luaurtas
f
Jql
Jo
Jrrrpu eq1,
'uoxa
())
luelsuor
Jq1 luorJ uoJlul aputs
e ,{q
palere
-das
tuaruEas
t
Jqt
Jo
stslsuo)
luaru8as
aua8
3
aql
'tuaru8as
(n)
alqerrel aql
rxorJ uolut
al3
-urs
e,{.q
paleredas (1)
uoxa JJppal aql
Jo
slslsuoJ
tuaru8as
auaS
4
eqJ
'g'flfr
Tsfl*gj{
ul
patertsnlll
se
'sued
o,ll,l rxoJ;
palqruessp
sr ureqJ
tq8q
1V
'aua6
3
aq1 6uLparatd
sluaubas
C
a^!J
Jo
auo
pue
luaubas
auab
1
e uaamlaq
uotlpurqulolal
alEurs e
fq
palqurasse
sl ureql
1qbrl
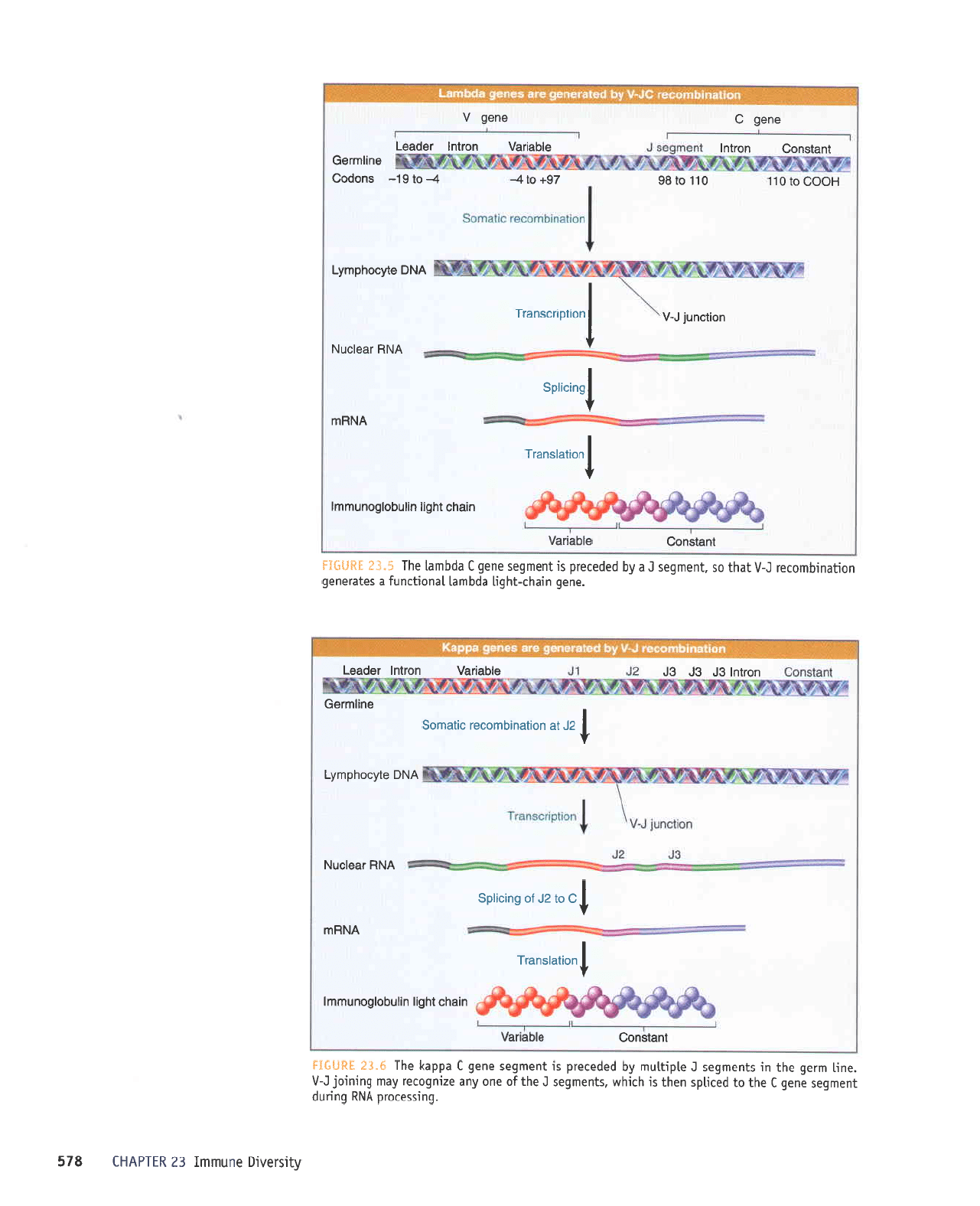
edde>1
y
r
'uotbar
6utpor-l
pue'uollut
'uoxa
6urpor-g
iloqs
p
seq
luau6as
auab
1-g
aq1
o
'uorbar
6urpor-olqPue^
pue
'uollur'uoxe
leppel e seq
lueubas
auab
I
aq1
r
'lUeUbas
auab
1-1
e
pue
auab
A
p
uaaMlaq uoqeulquolal
a16u1s e {q
palquasse
sr urpr.lr
lqbLt
epquel
y
.
uorlPurquolau
alburs e fiq
palquassv
arv sureql
tq6!l
.IJOI
)I1PIIIOS
IpuorlJunl
Jql uI
pJpnlJur
are
(sluaru8as
q
pue
sluaur8as
l
Eurpnpur) saruanbas
yNO
uoqs
raqto
'sluaruEas
aua8
3
pue
sluaru8as
aua8
n
aql
01 uoulppP
q
'ureql
ueaMleq
uolpPal uolleulq
-ruoJal
aqf
1o
z(lSurpuodsarror
pue
'sluaurSas
aua8
3
pue
sluaru8as
eua8
A
eql
Jo
uopeaue8ro
eql
Io
slrelep
Jql
q
saJueJaIJIp
arP
JJJqI
lnq
u(lrue;
qJea
uI aurps
aql ale
palquassP JJP
saua8
leuo4JunJ
qJlqa,(q
saldnutrd
aq;
'PeutoI
uJaq seq
leql
slueru8as
auaE
)
pue
stuJru
-8as
aua8
A
Jo
uollpJn8r;uor
relnrpred
aqt
.{q
pJurrrJelap fl
IIaJ
Juo zlue,{q
parnpord roldarar
IIaJ
J
ro
.{poquue aq1
'aua8
l,
auo
pue
aua6
g
euo Jo
'aua8
5l
e
pue
aua8 n
ue
sa8uBrrear
dlazrrpnpord
arboqdru,{1
;
e
,,{pepruts
'sluau
-3as
aua8
ureqr-zheaq
Jo
euo
pue
(1
ro
I
raqtta)
sluaru8as
aua8
urcqr-lq8q
yo
luarua8uertear
azrllrnpord
auo
z(1uo seq
u(lensn af,{roqdru,{.1
g
V
'sailroqdrudl
arnleurun
;o
uoIlPIn
-dod
e ur srnJJo
IJol
IEuoIlJun;
arrr8 ol sluJru
-8as
aua8
f,
pup
A
ueeMlJq
uolleulqluoJeU
'3un11ds
VNU
^q,{e,u.
pnsn
aql ut
pJlorrral
eJp
suoJlul aq1
'ugatord
Jql
Jo
suleruop
Ipuort)unJ
eqt
qllm .{lasnard
puodsarrol
teql
suoxa;o
Supstsuor
aua8
azrlDe ue seleJJJ
luaurEas
aua8
3
e raulred
o1
luaur8as
auaE
n
e
s8urrq
leql luale
uolleurquorar
aq1
'aua8
Detur
a18uts e,{q
papot sr-ad^&
Ineaq auo
pue
(1ro
x
raqtra)
ad&
tqSl
auo-sulPq)
sll
Jo
q)Pe
'q8noqr
'z(poqlrue
ue
EutssardxJ
IIa)
p
uI
'ruals^s
eunuruI
aq1
ueql
raqlo saEeaull
11p ]o
slleJ
Jlletuos
rn
pue
aurpura8
aql
ul
punoJ
sI
puP
'utailud
autlwa|
at4tpJIIpJ
sI srql'sluaruEas
auaS
3
pue
sluaru8as
aua8
n
qloq
Jo
les
uMo sll
Jo
slslsuo)
pup
auosoluoJqJ
lueralilp
P
uo saprsar
,{.1t
-r.upJ
qreg
'(g)
ureqr.{neaq;o
sadzb
eqt
1e
3ut
-ureluoJ
,{lrue;
Juo
puP
'Y
pup
>t
'sureqr
1q311
ullnqolEounulul
Jo
saIruPJ
o,lrl
JJB alar{J
'ursruBBro
eql
Jo
,{.ua3ord
,{q
patrraqul
lou
sI ua8tluB
ue
o1 asuodsal
aql snql
lauquua8 eq1
DaJJe lou
saop
pup
sllJJ
Jllpruos
uI sJnJJo
ssarord
artlua
aq;
'ua8rlup
Jql
pulq
01 elqP
utalord
e Eur,{r
-rPJ
slle)
JSOI{I Jo}
u1tpapsu\
slFsar
uaql
qJIqM
'ua6tluu
o1 atnsodxa
a41
atolaq
JnJJo
YNO
Jo
luelu
-a8uerrear
srql
leq1
sarrnbar
zfuoaqt uollJelJs
IeuolJ
aq1
'ua8pue
eql
qllM
DPaJ
01 slqPIIeAe
sI
11
lPql
os
pJZISJqlu,(s aq ot
rotdal:rTzlpoqllue
eql srllolle
stred
asaql
ruor;
aua8 JAIlre
ue
Jo
uorDnJlsuoJ
luanbasqns
aql'slueru8as
aua8
3
Jo
Jaqrunu
Jallpurs
E
pue
sluaur8as
aua8
n
Jo
Jaqrunu
a8rel
e sassassod
11
'roldarar
1ar
1
ro
,{poqllue
relnrtlred
e Sunnpord
ro;
aua8
IeuolDunJ
aql
ssassod
lou
sJop tustue8ro
aql
'qrrlq
lt7
'(ua8rlue
aqr)
ad,{rouaqd
aqr
}o
rrnl
-eeJ
JelnJrued
e ol
spuodsal
lPql
aruouaS aql
ut
[1rsranr6
aunuul
€Z
U]IdVHl
gf g
'6urssarotd
ypg
6uunp
luaurbas
aue6
3
aq1 o1
parnds
ueql
sr
qllqM
'quau6as
C
ar.ll
Jo
auo Aue
azruEolat feur 6ururol
q-1
'aurt
turab
aq1 ur
sluar.ubas
g
aldqlnur r{q
paparard
sr
luau6as
eue6
3
edde>1 aq1
g'IU
IU{.ltIJ
'auab
ureqr-1q6rt
epqurel
leuorllunJ
p
salerauab
uotlpurquoler
C-A
lpql
os
'1uau6as
g
e r{q
paparatd
sr
luau6as
aua6
3
epqurel
aql
S"tg
H&fl*Il
luBlsuoc
olqBuEn
u;eqe
1q6;1
urlnqol6ounuur;
I
uo,,",.r".,,
VNHUI
VNH
JEEICNN
yp6
e1tuoqduI1
I
tt
"
uo.eurqurocor
c'euos
ou!lruJee
elqP!Pn
uorlulrepea'l
I
t
ot
-r
1o
ouqqlds
uo4ul
gf
er
el
luElsuoc
alqEuen
u;eqc
1qOr1
urlnqo;6ounrlur;
luo*"'""','
VNUTU
vNH
realcnN
uoglcunI
p-1
uorloucsuell
VNO
ev{cor.ldr.uIl
HOOC
ol
0il
26+
ol
t-
t-
ol
6l-
suopoc
ourluJJ90
luelsuoc
uollul
alqeuPn
uoJlul
Jopee-l
fo'n"os
eueb
4
eue6
3
0tt
ol
86

619
suorlpurquolau
oml [q
palqurassy
arv sulpq] Aneag
E'g7
'luetubes
0
aql
ol
pautof
st
luaubes
aua6
n
e uaql
puP
'1ueu6as
1
e o1
paurolsLlueubas0plslrl'suorl]pa.l
6uLuLo[leLluanbasAqpalquassearesaua6finpaH
{'i.i::i:$*1]i:i
'(sa1grue1
rrrqt
IIe
ur)
tuaru8as
1
e
Lq
pue (zlpo
z(puey
g
aql ur)
luaru8as
q
aq1
.{.q papurord
sr uorSar
elqprrel eql
}o
rJpurpruJJ
aq1
'(8uo1
suopoJ
OOrtl
y1asll
uorSar
alqe
-rJpA
JrIl
Io
ued
Joleru Jrll JoJ sepoJ uoxJ
puof,as
eq1
pue
'(luaruqrette
auerqruJru
ut
pa,L1o,tut)
aruanbas
leu8rs
Jqt roJ sJpoJ uoxJ
tsJrJ
JqJ
'uorlezrueSro
ur Jelrurrs erp sJrFupJ ur1nqo13
-ounrurur
JaJr{l
ile
yo
sluaru8as aua8
11
aq1
(.SUOIlJPJJ
Sururof
f-O
pue
C-A
^q
pataldruor
uJaq
ser{
ssarord
Jql arunsse eM
'sureqJ
,{,teaq roJ sluJru
-3as
aua8
)
pue
A;o
Sururof ssnJsrp a.l,r
uJqM)
'suorlJeeJ
Sururof
O-A
pue
Sururof
f-O
Jq1 ul
pellolur
sr
luaurSas
qauus
erll
leql
eJnsuJ
lsnu
usrueqJeu uMou>lun auo5
'(a,rrtre
f,lrressa
-Jeu
IIp
lou)
stuaruSJs
q1
0€-
seq sn)ol
uelxnq
aqr
lqlSual
alqerJel
;o
sluaru8as
( JAIaMI
sureluoJ snJol ureqJ-d,teaq
Jsnoru
aq1
',{erre
uapuel
e ur
pazrueSJo
are slueur8as
q
aq1
('tuaru8as
aua8
H)
e aparard
leql
stuaru8as
f
IerJAes
]o
euo 01 uorl)auuor eql
Jo
suJel
uI
uoIlJeJJ eql reprsuoJ
tsn[11,r,r
J,tr
ilrou ro]
luoll
-eurquoJJU
yNq1
^q
prsne)
s1 Surqrtlu5
ssel)
'eI'€Z
uortJJS ur stuaru8as aua8
HJ
luJJeJJIp
Jo
JSn aqt ssnJsrp a14)
'(suoxa
IErJ^es
Jo
stsls
-uor
qlqm)
luaruSas
H3
tuarelpe
Jql
Jo
uoIS
-sardxa
01 speJl uorlJnrlsuoJJJ eql
'luaru8JS
pJurquoJ
HfO
eq1
qllM
saurquof,Jr
uaql
tuau
-8as
HA
B
jluaru8as
u1
e
qtl,r,r
sJurquroJer
stuarrl
-3as
q
Jql
Jo
Juo
lsJld
'
r i:
:i
;i
:j:
i
;r.i.ii
uI
pJlPJlSnllI
se
'sa8els
omt ur areld salel Sururo[
1-q-n
'sluaru8as
Hf
JnoJ Jql
pue
sluaru8as
HA
aqt
uaaMleq auosouoJql
aql
uo serl sluaru8as
q1
0I<
Jo
,{erre
uy
'luaru8as
f
eql
pup
luaru8as
n
aqt
z(q
pepo)
saJuJnbes
Jql
ueJMlJq sprJe
oullue
uJeulql
01 oM1 PJlxe
ue
yo
uratord
Jql uI
eJuesard
aql [q
para,rorsp
se,vr
(.{lrsranlp
ro})
luaur8as
q
JqJ
'luaru8as
Ipuourppe
u€ sJAIoAuI
uollJnrlsuoJ
uteqr,{.leag
'suoxa
lplanos Jo
slstsuol
lueubas
I
otll
.
'l-t-C
ol
I
sutol uoqeutQtuo]ar
puolas
aLlf
r
'l-C
ol
0
sutol uotleutquo]ot
]stQ
eLll
o
'luaubas
auab
3-g
e
pue
'lueubes
6
e
'aue6
n
p
ere uoqpurqulolal
uleql
r\neaq ro; sltun
oqi
o
su0rlPuLquolau
oMI
^q
pelquassv
uorlul
uoJlul
uollul
I
oullruj€e
luelsuoc
uorlul
tf-l.l
olo-to
arv sur.Pr.l3
fneag
'Surssarord
VNU
ul
leu8rs
3un11ds
str
sasn SururoIf-A
VNo
uI
paztuSora:
s1
tuaru8as
1
re^JqJrqM
'uoxa
J
eql 01 Sunrlds
JoJ
pJSn
aq
ueJ
leqt
,Lepunoq
rq31r
aql
1e
1eu31s
e ssas
-sod
osle
[aql
lluaur8Js
A
Jq]
qlIM
eulqluotJJ
o1 alqrssod
ll
sr>leu
teql
zlrepunoq
Uel
aql
1e
pu8ts
e ssassod
stueur8es
f
leuop)unJ
IIV
'(a:n8g
eql ur
tno
paxds sI
teqt
uorlul eql ul
papnlJur
st
g1)
suoxa
tuelsuoJ
pue
elqPIJeA
Jql
uJeMleq
uorlul
eql
Jo
lred
se
pelPJJl
sI
lueru
-8as
l
SururqruoJar
Jql
Jo
tq8r.r
aql uo
tuaru8as
I

JaAo
ppJJds
JJe sluJuSes
o
JqJ
'tueruSJs
(
lsrrJ
eqt
urorl
q>l
0Z
^luo dq
pateredas
sr rJt
-snlJ
un
Jql
Jo
Jeqrueru,€
eqI
'sluaur8as
aue8
)
Jo
uorlezrup3ro
pue
rJqrunu
Jql
ul eJuaJaJ
-JIp
1q3[s
p
pup
'stuatu8as
l
pue
C
raMJJ
's]uJrrr
-8as
aua8
HA
aroru Jle
JJJqI JrJqM
,esnou
aq1
uI JeIIIUIS
SI
1I
'i:'i'f
,: +i{r:,*};.j
ur
pJzrJeurrxns
sp
'suorlJJs
JIJIJSTp
IeJaAaS
Jo
slsrsuof, upur ur uorl
-rnpord
ureqr ,{rreaq
loJ
snJol a13urs
aq1
'sluaru8as
aua8
1
Jo
laqunu
peJnpJJ
Jql
Jo
esneJeq Llqerunsard
'ureqJ
tqSll
Jo
adLr
x
aqt
ssardxa slleJ
g
Io
./"56'Jsnoru
uI'\
erc
o/o17-
pup
y
aJe
sureqJ
tq8rl
aqr
p
./r09-'uetu
uI
'urcqr.dneaq
aqt
qll.ta
Jterlossp
ot ureqJ
tq3{
y e
J0
>r
e
JaLllta
saleraua8
a1,{toqdru,{1
ua,r13
y
'surlnqol;ounururr
aterauaS
ol
pesn
aq o1 d1a1rl
JrE
0E>
pup
'q8noqr 'sauaSopnasd
anrlreur
are sluaruSas
n
aqt
Jo
,{.uepsJJqrueru
IpJlseJue
Ipnprlrpur
yo
aruaSra,r.lp pue
uorlerrldnp
,{.q
pateraua8
Jrp r{trqM
'srJlsnlJqns
uroJ sluau8as
aua8
n
petelar
'saua3
pJtelJJ
Jo
sJrlrueJ
rJqlo
J>lrf
'sJeqruJtu
00 I
ot
Z
ruoJJ
JZrs ur ,{ren
teql
s:r1r
-rueJ'A
8I-
Jre errqt
pup'(saua8
0OOt-)
a8rel
z{11ensnun
sr ,{.gue;
Jsnotu
eqJ
'^tltuepl
pIJp
ourure
%09<
J^pq,{gure;
e
Jo
srJqruJru
leqt
uou
-JlrD
ar{t,{q
paugap
erp
qJrqM
'serlrrupJ
olur
pJprlrpqns
eq uet
stuaur8as
aua8
)r1
aq1
'stuJru8Js
auJB
)n
9g
;o
dnor8
Jeqloup
tuoJJ
uor8ar srql
saleredas
q{
009
yo
de8
y
'sluJu8es
euJB
)A
07
pue
stuJruBJS
,f
aAIJ
Jql surptuoJ
q{
009
;o
uor8ar
e
'tuaru8as
aua8
"3
aql 8ur
-parard
lsnl
'suor8ar
out
seq JJtsnlJ
ueunq JqJ
'uor8ar
luelsuoJ
eql
Jo
ueJJlsdn
'aruosoruorqr
eql uo relsnlf,
a8rel
e .ddnno
stuJrrrSes
eueS
'A
aqJ,
'(annreur
tuaqt
Jo
auo)
sluaru8Js
f
aArJ
,{q
paparard
sr
1r
q8noqlle
'tuaur8as
aua8
3
auo
Lluo
seq
sn)ol
>t Jql
teqr
s^{oqs
ia'f ;j
;ts{:=:j
'sluJruSJS
aua8
xA
aurlruraS
slr
Jo
tsotu
Jo
uouelap
rrqdorls
-eteJ
e
peJJlJns
Jsnoru
aql
'tsed
eqt ur aurl
eruos
lV
'JnrlJeur
sr Juo
'sluJruSJS
JueB
Y)
JnoJ
Jqt
JO
'suor8ar
)-f
out
ol
pJ>lurl
sr
qJeJ
lslueru
-8as
aua8
1A
oul
^luo
are areql
esnoru
p
ur
lpql
sr JJUJJJJJTp
ureu
aqJ
'snJol
ueunq aql
ueql
esJJArp
ssel
qlnlu
sr
asnorx ur
snJol
y JqJ
'lueu
-3as
1
ulro
slr Aq
paparard
qrea
'sluaru8as
aua8
)
g-
seq
snJol
y eqt
]eql
sMoqs
i+'ii
.if
:iijij
'stuaul8as
aua8
3
Jo
requnu
JJIIeus qJnu
e ot
pJlull
are sluau8as
aua8
n
Lueu
.{lruey
aua8
31
1q8r1
qJea
uI
'aur1ruraS
eql ur
parJJer
suor8ar
Surpor
eqt
Jo
^tJlrp,r
aq1 dq palepoutuoJ)e
aq
uet Alrs
-relrp
qf,nru
Moq
aas
ol sluaur8as
aua8
3
pue
n
yo
saddl
tueJaJJrp
eqt
aururexe
tsnru
JM MoN
^]lsle^ro eunuul
€z
ulldvHl
'[lquasse
euab
uteqr-fineeq roJ
uorleuloJul aql
llp
sutp]uol uput ur lelsnll euab
e16urs
V {t{,tl:
;***I*
09 00r 09r 002 092
00e
Ql
,n
e
,L
rL
,tA
,o
vh
,1" 61,
g
rl
9 0z-
00e-
slueurbes
aue6
g
I
slueubes
O
<seua6
HF
'luaubas
aua6
3
e16urs
p
ol
pelleuuol
sluau6es
C
a^g ol
pa>1uq
sluaubes eua6
A
Jo
lstsuol
sarlrueJ eddel
asnou
pup
upurnq aql
*'€* SHit$g.{
"c
n-tr
'n
of
"n
gg
'sluaLubes
auab
3-1;o.laqunu
lleus
p
ol
palurt
sluaubas
aua6
1Jo
stsrsuol [1Lure1 epquel
oq1
.*'gf
S]*f,:*ij
ueur ur
slueu0es euab
C-l
g<
ueu ul
00e-
osnour ur
slueur0es eue6
3-p
t
esnour ur
Z
:l :l
-:'[
r+i!:!'!!'rt:
e\ce\f
z\cz\n t\ct\l
sluaubes auebYl
'sluauDas
c
rnoJ
pup'sluaurEas q
67'saua6
n
ggg
6ururquor
Aq
sureqr
gggT<
arnpord upl snlol
H
uV
.
'seue6
3
a^q 01 lnoj
q1r,lr
sauab
n
ggE
6uLurqruor
fq sureqr
6991<
arnpord uel sntol ureqr
lqbrt
y
r
[1rsrenr6
anrsua]xl
salPraua9
uorlPu rq
uolau
'(dq
OOE-)
qerus
LlanrtelJJ
ere
z(1ensn
suoJlur
ar{J
'Jagoqs
sr
a8urq Jql
13uol
suopoJ
00I-
sl uoxJ
HJ
qJeA'tH)
pup
'zH) 'a8urq'tH3
suor8ar
aql JoJ apoJ
suoxJ JleJedas
'V'67
an?q
ur uMoqs
ureqf,
uralord aqr
qlIM
SulpuodsJJ
-roJ
lsuoxJ
IeJeAJs
,{q papor
sr uor8ar
luelsuof,
aql
'surpqJ
g
ro4
'(aua8
J^lDp
'papnrtsuoJar
aql
Jo
uoxe
pJrr{t
aql
sJruoJeq
qrrqazr)
uoxa
a13urs e,{q
papor
sr uor8ar
tuelsuoJ
eqt
'suleqJ
IqBII
y
pue
>r
qloq
rod
'ureqr
yo
addl
rql uo
spuadap
uor8ar
tuetsuoJ
Jqt
Jo
aJnlJnJts JqJ