Lewin Benjamin (ed.) Genes IX
Подождите немного. Документ загружается.

19,
sauosorxolql
posdeu/is
uoaMlaq
srnllo uorlpurqulolau
snoboloruoH
z.6I
ur spuerls
aFurs
Jo
lueluenlo^ul
Jql JoJ
Iapou
IeJauJS
p
sMoqs
S'6I
JrnSrd
'spuens
JISurs
uaeMlaq,{.lrreluauraldtuol
:aruanbas yo
stseq
aql
uo JJr{loue
auo ezruSorJr
o} sprJe
JrelJnu
JOJ rusrueqf,Jru
Juo z(1uo
1o-
Mou>l JM
'rrPo
JSeq
Ienpr^rpul
erll
Jo Ia^el
eql
1e
uorsrlard
qlr.nr pa8ueqrxa
aq
01
IPrJJlerx
s^^ollP
leql
Jeuueu f,rJr)
-ads
z(p8q
e ur salnJJlorrr
o.trl eql ur sJJuJnbJs
SurpuodsarroJ
Jo
rred .r(ue
uJJMlaq
rnJ)o
ol uoIlJPeJ
srql JoJ
alqrssod Jq
lsnlu
1I
'Jruosolu
-orq)
Ieqlo eql uor]
pllPruoJqJ
Jetsrs P
Jo
vNQ
xaldnp
eql
qlrM
1)eJetur
ol
prtpruoJqJ
Jalsrs
'sypq
xaldnp rno1
eql
JO
OMl A^lo^ur
luana
rarro-6urssoll
lenpnrpur
/{ue
1o
suoqlelalur ielnlaloul aql
'pua
eql
1e
fluo
elqLsLn seuroloq
a1e1s
palerrldnp
eq1
q6noqlle'(sprleuorqr
telsLs) serqdar orvrl
Jo
slsrsuol
qlrqm
J0
qtpa
'seuosoutolql
aql
1o
arueteadde
aql
fiq peugap
ere aseqdord
1o
sa6e1s aql'aseqdotd
lrloreul
lslg
oql 6uunp srnoro
uorlpurquolou
?.i:
;.
:liJillt:*
ruo
Jo
y51q
xaldnp aqt s,lrolle
leql
ruslueq)eru
p
serrnber
uorlpurqruoJeg
'VN11
Jo
sJlnlelou
xaldnp JnoJ suretuoJ
tuJlelrq
qJpa
os
'xaldnp
ygq
a18urs e suretuoJ
plletuorqJ
Jalsrs
qJeA
zsluJ^e
JSJqI JoJ srseq rPlnralolu aql
$
leq
a
'luapr^a
aruo)Jq
spueuroJr{J JnoJ
IIe
pue
esuapuoJ
sJruosoruoJq) aql
uJqM alqISIA rnPruar
eleursprqf, aqJ
'(s'*E
Isfi*Is)
luJ^a
JaAo-3uI
-ssoD
aqt
sluasa.rdar Bruser-q)
p
reql
sploq ssLpue
Ipuonpe{L
'rJAo
Sursson
rpauaS;o seJnlpal aql
lalpred
eterusprqf,
Jo
uoltnqlrtslp
pue
JJqrunu
Jt{J'Blprusp1qJ
pJIle)
satls
JlJrJsrp
1e
raqlaSot
pleq
eq 01 uaas
eq ueJ ,{aql
'aleredas
ol
u6aq
srruosoruorqJ eql
uaq
a'rJqlo
qJee
qllM pe>luu
uaqt
pup
ua>lorq
ureq enpq
(y1qq
1o
xaldnp e 8ut
-uretuoJ
qrea)
sprleruoJqJ
relslsuou o,lrl
q)IqM
ur
/uolunar
pup
a8p>IEeJq
e se
paluasarder
^dlensn
'sged
;o
a8ueqrxa
prrs,{qd
e seAIoAuI
seruosoruorqJ
uaeMleq
uollPulquo)ed
'saDads
uJJllrleq slrelap aql
uI uoIlPIJeA
aprM sr araqr
q8noqlp
'sanads
qJeJ
uI aJnlJnJls
f,rlsrJelJeJpqf,
e seq
q)lqm
'xalduror
leur
-auoldBuIs
p
Jo
ruJoJ
Jqt ut
palplJosse
d11era
-lpl
eJe sauosoruoJqr
aqr
'paraldruof,
sI ssaro.rd
Jq1 uaq6'8u1rpd aurosotrroJqr
ro
qsdeu,{.s
paller
sr ssarord aq1
'3o1oruoq
sll
qtlM pasodde
sr JruosoruoJqJ
qJee yo
qr8ual
eJIluJ Jql
Illun
spualxa
Surrte4
'sluelp^Iq
Sutruroy
'suorSar
Jrour Jo euo
ur rred o1 urSaq
pue
rJr{loue
auo
qreorddp
saurosoruorqt
snoSolouoq
aqJ
'y1qq
xaldnp
p
suretuo)
q)lqM
Jo
qJpe
'splleru
-oJqJ
JJlsrs oMl
Jo
slsrsuoJ
pue.dlsnoulard
paler
-qdar
seq sauosouroJqJ
asJqt
Jo
qJpf
'alqlst,r
aruolJq
seuosoruoJqJ
IenpIAIpu
q)tq,r.
1e lutod
aqt
dq
pJ>lJpru
sI slsoleru
yo
Suruut3aq aq1
'VNo
Jo
saxaldnp
uJJMleq
leualeur
3ut
-Sueqrxa
ur
pellolur
eJe
leql
suollJpJelul
reln
-Jelou
Jql
qlrM
aseqdord
rrloleru
Jo
sa8els a.tq
aqr
q8norqr
saurosoruoJqr
;o
ssarSord alqrsrn
aql saredruoJ
t"*T sHili*x;i
'slsolaur
;o
aseqdord
paperlord
aqt
Suunp srnJ)o
uolteulqruoJeU
'uppruoJq)oJelaq
Jo
suor8ar ellpern
pue pJsuep
-uoJ
eql
;o
LlruDrrr aql
ur
passarddns
sI JeAo
-Sursson'aldruexa
JoJ
jeJnlJnJls
euosotuoJqJ
uodn spuadap
rbuanbar;
str
'aruouaS
eql ulqll6
'suerunq
Jlpru ur
se apruJ] ut
trltuanbarJ sp JJIM1
s.rnJJo
uolleurqruorar
lrurads uI
pue
salu(roo
ul
luJJJJJrp
aq
,(eru .{ruanbar;
IIeJe^o
JqJ
'spaJJe
le)ol
pup pqop qtoq z(q
paruangul
sI
tnq
'aruouaS
aql
tnoq8noJlp luelsuoJ
lou
sI uoueulq
-ruoJJJ;o
l:uanbary
aq1'(sraqro
JeAo
peJo^e;
eq z(eru saruanbas
;o
sad,{.1
aruos
q8noqtle)
sJteJlsqns se saluanbas
snoSoloruoq;o
rted
Lue
esn
ueJ alqrsuodsar
saui.zua eql
leql
sI JJnl
-pel
IeJrtrJJ
sll
'VN11
;o
saxaldnp
o,rtt uJaMlaq
uorlJeeJ e
sr uorlpulqruoJal
snoSolourog
uor]€ruul
SplleuojLlc lolslS o/vu
lo
slsrsuoc
puP
'polecrloor
seq
aurosou.roJqc L.lcPf
suor6er
ro uor0el
payu,rr;
ur 6uured
urbeq seurosouotq3
aue1o6{7
suorlceralur relncalon
srsorour
l.lono.r
Lll ssoroord

qree
uorJ
puprls
Juo
Jo
Surlsrsuol
uorSar e seq
xaldnp
qtpJ
'uorleurqruoJJJ
Jo
atrs eqt
tV
'1u1oI
lueulqruo)ar
Jqr
prllpJ
sr
rJqto
eqt 01 xaldnp Juo
ruorJ sJSSo-r)
VNC
Jo
pupJts
Ienpr^rpur
ue
q)rqnl.
lp lurod
JqJ
'alnJalotu
lurol
e
pJIIe)
sr saxaldnp
;o
led
pJtJJu
-uol
JqI'saxaldnp
VNC
o..lr1 eql ueeMtJq uorl
-)auuoJ
e saleaJJ a8ueqrxa
lerordoar
aq1
'xaldnp
Jeqto Jqt ur
tuJrueldruoJ
slr
qllM
Jred
01
rJAo
sassorJ
pue
rauued su sJlpel
pueJls q)eE
's>lJru
aqt ,{q
palEJrJ
spuJ JJJJ Jql
Jo
tuJruJloru
smollp
a8elearq aq1
'saxaldnp
ypq
parred
oMt
Jo
spueJts snoSoloruoq
aqt
;o
slurod
SurpuodsarroJ
Jql
te
a8elearq
qtru
suels
teqt
ssarord e
sJlpnsnllr
* +=i
igi:1:ij
'uoIleuIqruoJel
ur
pe^lolur
sassaro,rd
aql SurpuelsJJpun
ol
IpJluJJ
are deazr
srql ur
pJleeJ)
saln)Jloru aql
;o
sarlradord
aql asneraq
'(palerlrur
L11en1re
sr
tr
aroq dUressarau
lou
sr srqt
tpql
JJS
lleqs
a,vr
q8noqlp)
sa8ueqrxa puerts-alSurs
Io
stuJJl
ur uorllear
uorleurqruoJer
eql eur8erur
ot
1n;
-esn
sr
lI
'uorlpurquo)al
ur
spueJls a13urs Lrel
-uarualdruoJ
uaJMtJq
Surrred eseq
Io
Jsn Jql
Jo
MJrA:no
Surpuedxa
.dq sgels
JroJeJeql uoll
-eurqruoJJJ
;o
srs.dleue
Jpln)aloru
rn6'ssarord
uorteurquroteJ
ar{]
Jo
ueeq
Jql
]p
sl
vN(I
Jo
seln)Jloru xaldnp
o,trl SurtrauuoJ
Jo
lJp
JqJ
'uorlnlosar
6uunp
pellru
etp
spup.lls
Jo
tLed taqlo eq1 ro
abueqrxa
leurbuo
oLll ur
pello^ur
spuells eql leqlaLlM
uo spueoep
poul.loJ
a.re
sluPutquola.r leqleqfi
.
'spu
Pjls
burlrauuor eqlJo
oml 6uLlrru
fq salnralour xaldnp
elplpdas
o/v\l olur
pe^losal
sr elnleloul
luLol
eql
.
'alnlelou
lurof
e aletaue6
o1 firessarau
ere sa6ueqrxa
(lerotdtret)
orvrl
o
'luared
qree
uol1
puplls
auo
1o
buqsrsuoo
y116
xoldnpolalaq
Jo
qtlolls
e saleteuab
ebueqrxa aql
.
'arnllnrls
paqluPlq
e soleait
1r
'xaldnp
laqlo aql ur
ledtelunor
s1r sareldsrp xaldnp
auo
ulolJ
puells
e16urs
e uo{/!\
r
'spuPtls
a16uLs;o a6ueqrxe
sr selnlalout
y116
xaldnp
oMl
uoaMloq uoqpurqurolel
u!
lua^a
fie1
aq1
r
VNO
xaldnporalaH
sa^lo^uI
uoqeurquiolau l$Dads-a]l5
pue
snobolor.uoH
6I
ulldvHl
uee^l.leq uorlJPJJlur seAIoAur uorlJPeJ
l€rJel
-Jpq
aql
'seln)Jloru
xaldnp uJJ,tlrtJq a8ueqrxa
)IlEUE8 JSNE)
lPql
UMOU>I JJP SJIlIAIlf,E
IEIN
-Jelou
qJIqM
ur
'PrJJlJPq
ur alqelrenP sr ssarord
uorleurquoJJr aql
lnoqe
uorleruJoJur
pJIrPtJ(
'srsorJru
;o
sa8els
aterurxordde
qllM
sdals reln
-JJIoru
Jqt
Jo
euos elplJJJoJ ot alqrssod
tr
apeu
seq
lsea,{
ur sluelnru
Jo
uorlelosr eql JaAJMor{
Llluarau
'(panresqo
z{1aso1;
tsoru
uJJq seq srs
-oreru
qJrqM
ur) s11ar JltoIJe>lna
raq8rq ur uoll
-PurqruoJJJ
ur
pJAIoAur
slue^e JPlnrelolu Jql
lnoqe
uoIlPuJoJuI
pelelJp
ou sI aJaqJ
'sJluos
-oruoJqJ
Jql
Jo Ie^el
Jql
le
pJAJJsqo
3Je
leqt
sa8ueqr Jqt
qll,lt,{.1sno:o3rr
stuene
rpln)Jloru
eseql elelal arnlrunI srql
le 1ouuef,^e14
'uorunJr
pue
atsP>leJJq
e
Jo
spueuep Jqt ot spuodseJJoJ
leql
JJAossoJJ
p
Io
sueJu
[q
salnralou
xaldnp
letuared
Jqt
tJauuoJ
ot alqrssod sr
1r
'uJql
3u1unr
relpl
pue
spuerts
qtoq
Sur8upqrxa ,{g
'saxaldnp
Jql UJaMlJq
UOII)aUUOI
JAISUeJXJ eroru
Jq uer JJeql
'pJpuelxa
sr a8ueq)xe
pueJls
aqt
;1'saruanbas
Surpuodsauo)
te
pat)Juuor.dlerr
-;nads
Jq
IIIM
sJInJJIoru xaldnp o.trl eql
'xaldnp
JJqlo Jql
ur
puerls
SurpuodsarroJ
Jql sareld
-srp
puprls
auo
(tsea1
1e)
;1
'pasealeJ
Jq ueqt ueJ
xaldnp
teqt
Jo
spuprts rqt
Jo
qtoq
ro aug
'xaldnp
vNO
q)pJ
ur
>leeJq
e J>lpur o1 sr spuerls a13
-urs
Surprnord
ur dats
tsrrJ
JqJ
'uorteurqruoJJJ
'sy116
leluared
oMl eql
Jo
spup.lls Arelueuald
-ulor
uaamlaq 6uured sanlonur uorleurqurolau
11"*i
3*i"i*Ij
sluPurquloceu
I
alerpor.!Jelur uorleurqurocou
t
salnsalour
vNo leluaj€d
uorunau puP
e6e)Par8
@
'leAJI
Jeln)eloru
eq1
te
ureldxa
ol a8ets
tlnJrIJrp
lsoru
eql
surpureJ sJurosoruoJqr
rpofuelna;o srs
-deu,{s
aqJ
'seuouaS;o
Surrred arrluJ
ue ueql
JJqlpr
'q8noqr
'auroua8
aql
yo
suor8al
paDrJlsar
DNA
duplexes
parr
Homologous
strands
are nicked
Broken
strands
Crossover
point
moves oy
branch migration
Nicks are
sealed
I
I
exchange
between duplexes
11
Second nicks
/ \ Second nicks
made in same strand
made
in other strand
|JlTio"*
I
Genomes
are not
recombinant, but
coniain heteroduplex
regron
Reciprocal recombinant
genomes
are
generated
fl**R*
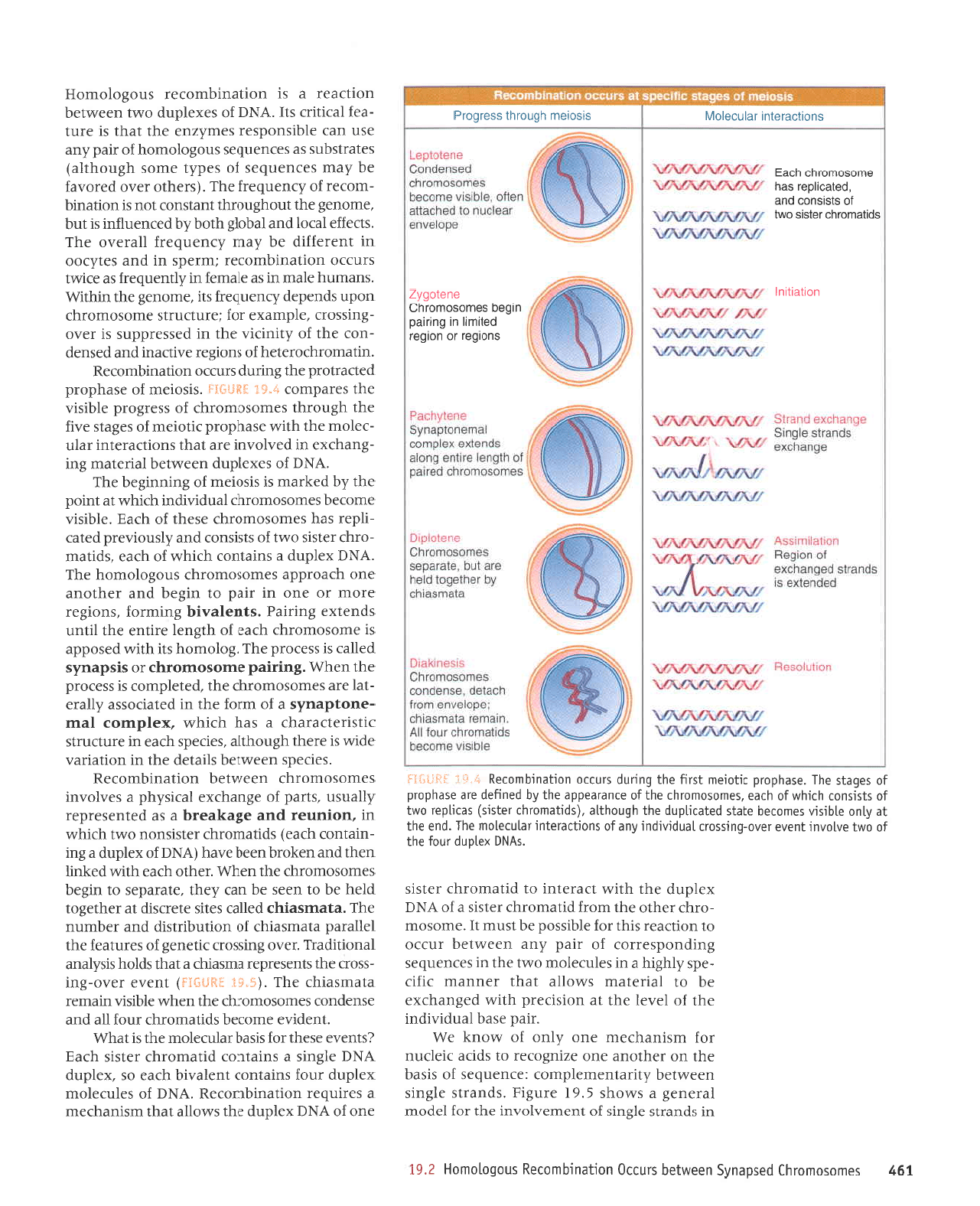
3ll.S Recombinatjon
between two
paired
duplex
DNAs
coutd
invotve
reciprocal
singte-strand exchange. branch
migration,
and
nicking.
of the
parental
DNA
molecules. This region is
called hybrid DNA
or
heteroduplex
DNA.
An important feature
of a recombinant
joint
is its ability
to move along the
duplex.
Such
mobility is
called branch migration.
Fi*13fi[
1*.;
illustrates the migration
of a single
strand in a duplex. The
branching
point
can
migrate in
either direction as
one strand is dis-
placed
by the other.
Branch migration is important
forboth the-
oretical and
practical
reasons. As
a
matter
of
principle,
it confers a dynamic
property
on
recombining structures.
As a
practical
feature,
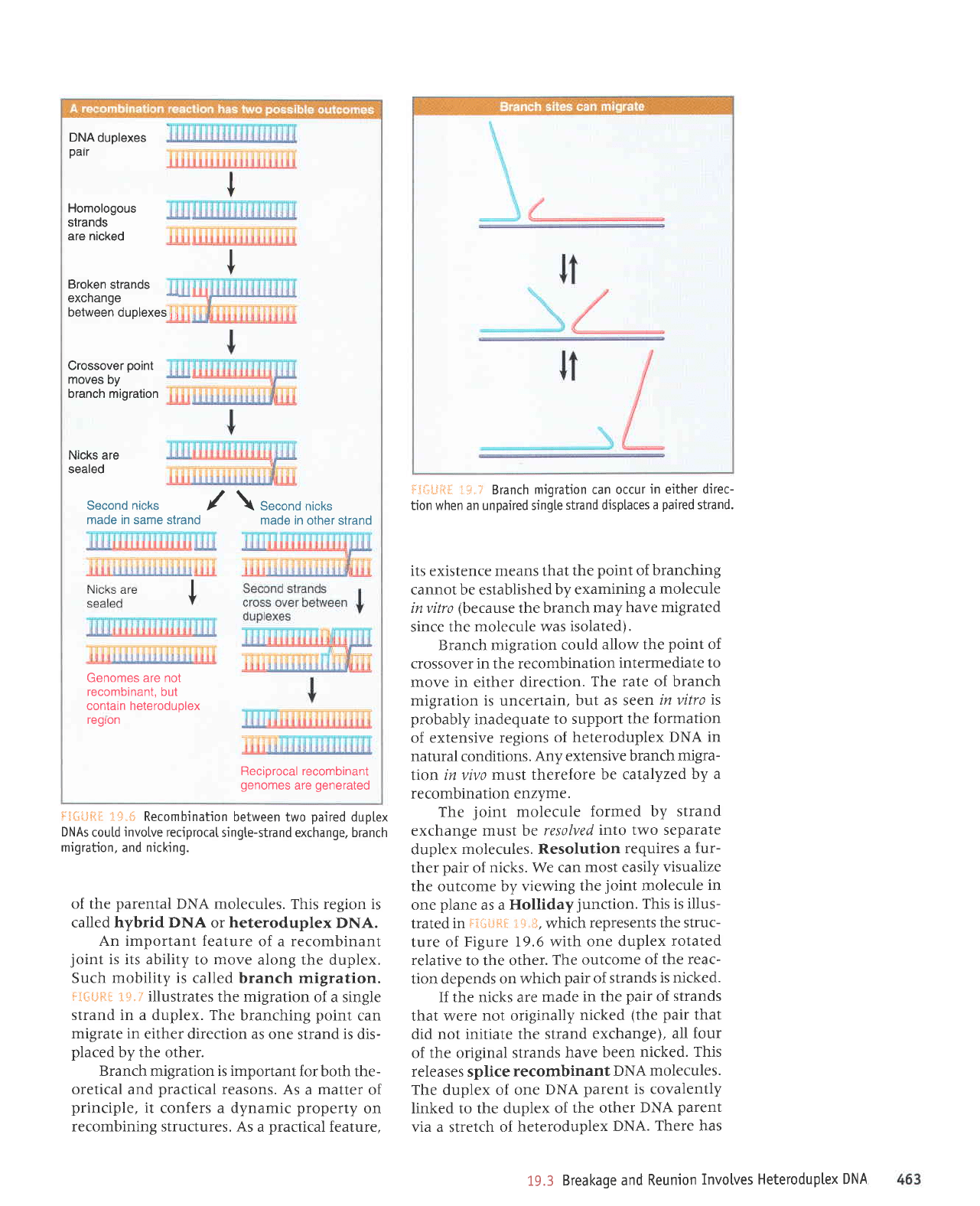
flEGUft$.
t#.? Branch
migration can occur
in either direc-
tion when an unpaired sing[e
strand displaces
a
paired
strand.
its existence means that
the
point
of
branching
cannot be established
by examining
a
molecule
invitro
(because
the branch
may
have migrated
since the molecule
was isolated).
Branch migration
could
allow the
point
of
crossover in the recombination
intermediate to
move in either direction.
The rate of branch
migration is uncertain,
but as seen
in vitro
is
probably
inadequate
to support
the formation
of extensive regions
of heteroduplex
DNA
in
natural
conditions.
Any
extensive branch
migra-
Iion in vivo
must therefore
be catalyzed
by a
recombination enzyme.
The
joint
molecule
formed by
strand
exchange must be
resolved
into two separate
duplex
molecules. Resolution
requires a {ur-
ther
pair
of
nicks. We can
most easily
visualize
the
outcome
by viewing
the
joint
molecule
in
one
plane
as a
Holliday
junction.
This is illus-
trated
in
Fi*{Jltf
ii}.i}, which
represents
the struc-
ture of
Figure I9.6 with
one duplex
rotated
relative to the other.
The outcome
of the reac-
tion depends on
which
pair
of strands
is nicked.
If the nicks are
made
in the
pair
of
strands
that were
not originally
nicked
(the pair
rhat
did not initiate
the strand
exchange),
all four
of
the
original
strands
have
been nicked.
This
releases splice
recombinant
DNA
molecules.
The duplex of one
DNA
parent is covalently
linked to the duplex
of
the other
DNA
parent
via a
stretch
of heteroduplex
DNA.
There has
19.3 Breakage
and Reunion
Invotves Heteroduptex
DNA
Nicking controls
outcome
Nicks in
same
strands release
patch
recombinants
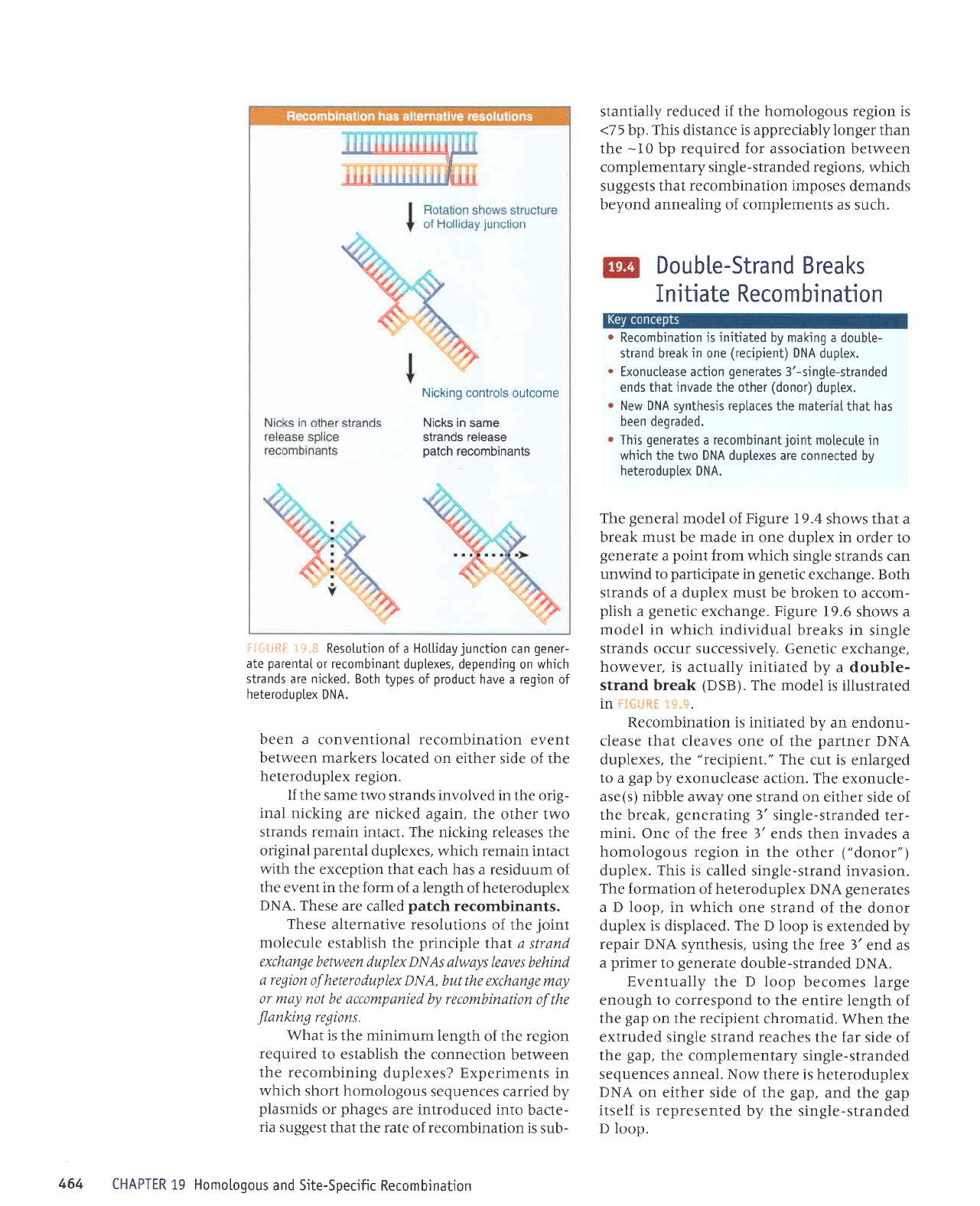
i:+ij*lF 1*.*
Resotution ofa Hotlidayjunction
can
gener-
ate
parentaI
or
recombinant
duplexes,
depending on which
strands
are nicked. Both
types of
product
have a region of
heteroduotex
DNA.
been a conventional recombination
event
between
markers located
on either side of the
heteroduplex
region.
If
the same two
strands involved in the
orig-
inal
nicking are nicked
again, the
other two
l'",:l*,;xniT,1il;lllff
[T?.;T,ff :,'l;
with the exception
that each has a residuum
of
the event in the form
of a length
of
heteroduplex
DNA.
These are
called
patch
recombinants.
These
alternative resolutions
of the
joint
molecule
establish the
principle
that a
strand
'
:'l
;,:!;
:; #;:: !:; l
:
:
: tr i' ;y #: :::;';:;'#l
';"7';':'::'accompaniedbvrecombinationofthe
What is the minimum
length
of the region
required
to
establish the connection
between
the recombining
duplexes? Experiments
in
which short
homologous
sequences carried
by
plasmids
or
phages
are introduced into
bacte-
ria suggest
that the rate
of
recombination
is
sub-
CHAPTER
19 Homologous
and Sjte-Specific
Recombination
stantially
reduced if
the
homologous
region is
<75bp.
This distance is appreciably longer than
the
-I0
bp required for association between
complementary
single-stranded regions,
which
suggests that recombination imposes demands
beyond annealing of complements
as such.
Double-Strand Breaks
Initiate Recombi nation
Recombination is
initiated
by making a
doubte-
strand break
in
one
(recipient)
DNA
duptex.
Exonuclease action
generates
3'-single-stranded
ends that invade the other
(donor)
dup[ex.
New DNA
synthesis
reptaces
the material
that
has
been degraded.
This
generates
a recombinantjoint motecute
in
which the two DNA duptexes are connected
by
heteroduolex
DNA.
The
general
model of Figure 19.4
shows that a
break
must
be made in one duplex in
order to
generate
a
point
from which
single strands can
unwind to
participate
in
genetic
exchange. Both
strands of a duplex must be broken to
accom-
plish
a
genetic
exchange.
Figure
I9.6 shows a
model
in which individual breaks
in single
strands occur successively.
Genetic exchange,
however, is
actually
initiated
by a
double-
strand break
(DSB).
The
model is illustrated
in il5$l.JR* t$.*.
Recombination is initiated
by an endonu-
clease that
cleaves one of the
partner
DNA
duplexes, the
"recipient."
The
cut is enlarged
to a
gap
by exonuclease action. The
exonucle-
ase(s) nibble
away one strand on
either side of
the break,
generating
3'single-stranded
ter-
mini. One of the free 3' ends
then invades
a
homologous
region in the
other
("donor")
duplex. This is called
single-strand invasion.
The formation of heteroduplex
DNA
generates
a D loop, in which
one strand of the
donor
duplex is displaced. The
D loop is
extended by
repair
DNA synthesis, using
the free J'end
as
a
primer
to
generate
double-stranded
DNA.
Eventually
the
D loop
becomes
large
enough to
correspond to the entire
length of
the
gap
on the recipient
chromatid. When
the
extruded single
strand reaches the far
side of
the
gap,
the complementary
single-stranded
sequences
anneal. Now there is
heteroduplex
DNA
on either side
of the
gap,
and the
gap
itself is represented
by the
single-stranded
D
loon.

The duplex integrity
of the
gapped
region
can
be
restored
by repair synthesis
using the 3'
end on the left side
of the
gap
as a
primer.
Over-
all, the
gap
has been repaired
by two individ-
ual rounds of single-strand
DNA synthesis.
Branch migration
converts
this structure
into
a molecule with two recombinant
joints.
The
joints
must
be
resolved
by cutting.
If both
joints
are resolved
in the same
way,
the original
noncrossover
molecules
will be
released, each with
a
region
of altered
genetic
information that is a footprint
of the exchange
event. If the two
joints
are resolved in opposite
ways, a
genetic
crossover is
produced.
The structure
of the two-jointed molecule
before it is
resolved
illustrates
a critical differ-
ence between the double-strand
break model
and models that invoke
only single-strand
exchanges.
.
Following the
double-strand break, het-
eroduplex DNA has been formed
at each
end of the region
involved in the
exchange. Between
the two heterodu-
plex
segments is the region
correspon-
ding to the
gap,
which now has the
sequence of the donor DNA in
both
mol-
ecules
(Figure
19.9).
Thus the arrange-
ment of heteroduplex
sequences
is
asymmetric, and
part
of one
molecule
has
been converted to the sequence of
the other
(which
is why
the
initiating
chromatid is
called the recipient).
.
Following reciprocal
single-strand ex-
change, each
DNA
duplex has heterodu-
plex
material
covering the region from
the initial site
of
exchange
to the
migrat-
ing
branch
(Figure
19.6). In variants of
the single-strand
exchange
model in
which some DNA is degraded and resyn-
thesized, the initiating
chromatid
is
the
donor of
genetic
information.
The double-strand
break model does
not
reduce the importance of the formation of het-
eroduplex
DNA,
which
remains
the only
plau-
sible
means by which two
duplex
molecules
can
interact. By
shifting the
responsibility for initi-
ating recombination from single-strand to dou-
ble-strand breaks, though, it influences our
perspective
about the ability
of the cell
to manip-
ulate DNA.
The involvement of double-strand breaks at
first
seems surprising. Once a break
has
been
made right across a DNA molecule, there is no
going
back. Compare the events of Figure
19.6
and Figure 19.9. At no
point
in the single-strand
exchange model
has
any information been
lost.
lilirliill.
.L',r,ii Recombination
is initiated by
a double-strand
break, fotlowed by
formation
of single-stranded
3' ends,
one
of
which m'igrates
to a homologous
duplex.
In the double-strand
break
model,
though,
the
initial
cleavage
is
immediately
followed
by
loss
of information.
Any error
in retrieving
the
infor-
mation
could
be
fatal. On
the other
hand, the
very ability to
retrieve
lost information
by
resyn-
thesizing
it from
another
duplex
provides
a
maior safetv
net for
the cell.
Recombining
Chromosomes
Are
Connected
by the
SynaptonemaI
Complex
.
During the early
part
of
meiosis.
homologous
chromosomes
are
paired
in the
synaptonemaI
comotex.
.
The mass of chromatin
of each
homotog
is
separated
from the other
by a
proteinaceous
co m
p
lex.
A basic
paradox
in
recombination
is that the
parental
chromosomes
never seem
to be
in close
enough
contact
for
recombination
of
DNA
to
19.5 Recombining
Chromosomes
Are Connected
by the SynaptonemaI
Complex
465

iia::-:rii
::-:. -::: The
synaptonemaL
complex brings
chromosomes into
juxtaposition.
Reproduced
from D. von
Wettstejn. 1.971.. Proc.
Natl.
Acad.
Sci.
USA. 68:851-855. Photo
courtesy
of D. von Wettstein,
Washington
State University.
ir
;:.ii:jiii
.i*-
j,;.i
Each
pair
of sister
chromatids has
an axis made of
cohesins.
Loops
of chromatin
project
from
the axis. The
synaptone-
mal
complex
is formed
by tinking
together
the axes via zip
proteins.
occur. The
chromosomes
enter meiosis
in the
form
of replicated
(sister
chromatid)
pairs,
which
are visible
as a mass
of chromatin.
They
pair
to
form
the
synaptonemal
complex,
and it has
been
assumed for
many
years
that this repre-
sents
some
stage involved
with recombination-
possibly
a necessary
preliminary
to exchange
of DNA.
A more recent view
is
that the
synap-
tonemal
complex is
a consequence
rather
than
a cause
of recombination,
but, we have
yet
to
define
how
the structure
of the
synaptonemal
complex
relates
to molecular
contacts
between
DNA
molecules.
Synapsis
begins
when
each chromosome
(sister
chromatid pair)
condenses
around a
struc-
ture
called
the axial element,
which is appar-
CHAPTER
19 Homotogous
and
Site-Specific
Recombination
ently
proteinaceous.
The
axial elements
of cor-
responding
chromosomes then become
aligned,
and the synaptonemal complex
forms as a tri-
partite
structure,
in
which the axial
elements,
now
called lateral elements,
are separated
from
each other by a central
element.
F:*iififl
:il"3i:
shows an example.
Each chromosome at this
stage appears
as
a mass
of chromatin bounded by
a
lateral
ele-
ment.
The two lateral elements
are separated
from each
other by a
fine,
but dense,
central
element. The triplet of
parallel
dense
strands
lies in a single
plane
that
curves and twists
along
its axis. The
distance between the homologous
chromosomes is considerable
in molecular
terms, at more
than
200
nm
(the
diameter of
DNA is 2
nm). Thus a major
problem
in under-
standing the role
of the complex is that,
although
it aligns homologous
chromosomes,
it is far from
bringing homologous DNA
molecules
into
contact.
The only visible link
between the
two sides
of the
synaptonemal complex
is
provided
by
spherical or cylindrical
structures observed
in
fungi
and insects. They lie
across the
complex
and are called nodes
or
recombination
nod-
ules; they
occur with the
same frequency
and
distribution as the
chiasmata. Their name
reflects
the
hope
that they may
prove
to be
the sites of
recombination.
From mutations
that affect
synaptonemal
complex formation,
we can relate
the
types of
proteins
that are involved
to its
structure.
IItuiJiqil
i.*"1: presents
a molecular
view
of the
synaptonemal
complex. Its
distinctive
struc-
tural features are
due to two
groups
of
proteins:
.
The
cohesins form a
single linear
axis
for each
pair
of sister chromatids
from
which loops of chromatin
extend.
This
is equivalent
to the lateral
element
of
Figure I9. 10.
(The
cohesins
belong
to a
general group
of
proteins
involved
in
connecting
sister chromatids
so
that they
segregate
properly
at mitosis
of
meiosis.)
.
The
lateral elements
are
connected
by
transverse
filaments
that are
equivalent
to the central
element
of Figure 19.10.
These are formed
from
Zip
proteins.
Mutations in
proteins
that
are needed
lor
lateral
elements
to form are found
in
the
genes
coding for
cohesins. The
cohesins
that are
used
in
meiosis include
Smc3p
(which
is
also used
in mitosis)
and Rec8p
(which
is
specific
to meio-
sis and is related
to the mitotic
cohesin
Scclp).
The
cohesins appear
to bind
to specific
sites
along
the chromosomes
in both
mitosis
and
466

L9'
s)earg
puprls-alqnoo
reue
surol xalduol
lpLuauoldpufs
eql
g'6I
ur
Suured JurosouroJqJ
r{srloqp
lpql
suoll
-Elnru
[e
leql
uMoqs seq >lJoM
tue]ar
pue
'paqs[
-qelsJ
IIJM
sr uorteuloJ xJlduo)
Ipuauoldpu^s
pue
uoleurquroJJJ
ueeMlJq uoueleJJoJ 3qI
vN(
xJldnp
snoSolouoq
Jpenur ol spuJ
pJpueJls-elSurs
aql selqpuJ
uaql
dnorS JJqlouV
'spuJ
pepue-rts-alSurs
Ho-,€
Surpnrto:d
olur s>lpJJq
pueJts-JIqnop
JI{t
ueluoJ
o1
parrnbar
sr suralord
yo
dnor8 aug
's>leeJq
pueJts-elqnop
aql ssarord ot
parrnbar
are
suralord
raqlo auru
tspJl tV
'uortrp
aseapnu
Lq
peMolloJ
urqt
sr
IIods
Jo Ielorua5
'xaldruor
I I
ods Jql salerJossrp
teqt
uralord rJqtoup
qlIM
uorlf,eJJlur ue dq
arnlrnJls
lueuelu:ad
e olur
peue^uoJ
sr
>leaJq
eqt
1VNC qtrm ,ilqrsranar
st)prelur
I I
ods
leql
stsaSSns
ij
i.'
i:r i
,it
ai1"ri
j
ul
uMoqs uorDeJJ
slql JoJ
Iepou
JqJ'$lPerq
puPns
-Jlqnop
eql setpJJuJB
lBql
au.,i.zuJ J>lrl-aspJeru
-osrodot
e aq ,{.eru
I Iods
teqt
slsaSSns sIqI
'seseJeruosrodor
I
ad,{r
;o
Llue;
e
Jo
strunqns
rr1L1e1er
Jqt ol snoSoloruoq sr
qlrqM
'I
Iods
utalord
aql o]
palJauuo)
JJe s>lparq
pueJls
-alqnop
rql
Jo
spue
,E
eql
'sluelnru
)spat
uI
'>leaJq
pueJls
-Jlqnop
eql
Jo
alrs
aql uoJJ saseeJrur
eJuelslp
se
paleraua8
aq
IIIM
uor8ar
papuens-a13urs
e
leqr,(rmqeqord
Suruqrap aqt Lq
prurruratJp sI
luarper8
eqJ
'uorleurqruoJeJ
ro;
Lressarau are
s>learq
pueJts-Jlqnop
leqt
stsaSSns srql
'uorl
-eurqruoJar
ur JArpelJp sr uorsnnord
pJpuerls
'slPa.rq
pu
erls-olqn0p
Jo
spua,9 aql ol
peuLo[A11ua1enor
sL
11od5
ri
i'i; i
;]i,:jit:ii;r
-a18urs
eql o1ut
pua qsnlJ
aql
Jo
uolsre^uo) eql
$lrolq
leql
(ospat)uouelnru
1sea.d
y
'6'61
arnSrg
Jo Irporu
eql
q
umoqs
sp
'spue
papuBrls-al3
-urs
,g
3uo1
otur sepls
qloq
uo
pJuanuo)
,{ptde:
Jre >leJrq
puPJls-elqnop
Jq]
zi.q
palea;r
spua
lunlq
JqI
'sruJel
JelnJelolu
uI s>lEeJq
pueJls
-Jlqnop
Jo
JIoJ aqt
tardlalut,uou
r{eu a14
'1I
tuoJJ
peards
11tLr
stuene uortpurqruoJeJ
eqt
leql
z(itlqeeord aql
spalJeJ
luarper8
eqt
pue
'pateppl
sI uoltpulq
-uro)Jr
qJIqM
le
etIS Jqt
sJIJItuepI
lodsloq
aq1
'1ods1oq
aql
Jo
sJpIS
qloq
Jo auo
uo
luatper8
e ut
seullJap uopeurqrxoJar
;o
trruanbJJ] JqI'uperu
-oJqJ
Jo
suor8ar
alqrsse)le arou
qll,lr
apI)uIoJ
o1
leraua8
ur
pue
suor8ar
raloruord
uI rnJ)o ol
pual .daq1
'rqoads
aruanbas
tou
eJp suolleJol
rraqJ
'uorleulqruo)JJ
ro; slodsloq
apr,tord
leqt
sJlrs
lP
Srsoraru
ur,{.Fea JnJf,o
oslP s>leeJq
puPJls
-elqnog
'(snto-IJVW
luatdttag
aqt
.dq
paterlrul
sI uorlrsodsue{
IeuoItJJrIpFn
'€Z'6I
uolttes
aas)
raqloue
.{q aruanbas euo
Jo
luaruareldar
Jqt sa^lo^u
r{JIqM
'adrb
Suueru
yo
a8ueqr
aqt ur
palerrldrur L1er1ru1 eJe,lr s>leJrq
pueJls-elqno(
'uorleurqurorar
rt;nads-elIS
pue
snoSolouoq
qloq
ur
uorleurqruo)er
elPIlIuI s>lealq
puerls
-Jlqnop
teqt lseau(
uI af,uJplna
poo3
sr eJeql
'uuo]
louuel
xalduror
leuauoldeufis
aql
'pelrolq
st uot]PutQtltolet
Jl
r
'sLuio;
xalduor
leueuoldeuAs
aql
aloJaq tnllo
uotlPUtqu0l0l
aleqtut
lPql
s)Pelq
puells-alQn0[
r
slParB
puPrls-alqnoo
JauP
sruJol
xalduol
rL rL
a .
lcPllE
I
osealcnu
Aq
pa/v\ollol
I
ttods lo le^ourag
r-..J
I
f
eoeneerc otqrsre^eu
lPruouoldeufi5
aq1
'srrcd
prleurorqr
JJlsrs
Jq1
Jo
sluJruele
IeJJlel
Jql
pJuuol
]eql
sluJru€lrJ
esrelsuer]
ruJoJ sule]oJd
dt7
1o
dnor8
aqJ'Idlz
qtlmpazllerol osp arc'2dr7pue
7dr7
'suralord
raqlo
o^4L
'sluJruelJ
IeJelel
aql
ut
pezl
-lPJol
sr
ureuop
IeuIruJJl-)
Jql
lnq'luJruJIJ
IeJt
-ueJ
eql uI
pJZIIeJol
sr
uralord
IdIZ
Jo
uleruop
IeurrurJl-N
aq1
'pasdeu.ds
Llasolt aruoJJq
tou
op,{aqr
rnq'pau8rle
aluof,aq
01
pue
uJoJ ol
stuJrlJlJ
IpJJtel
sMollp
uouelnu
1dg
a'q1
'slupurquoJal
Jo
uollpruJoJ
>llolq op
.daql
's>leJJq
puPrls-Jlqnop
Jo
uoIlPuJoJ
Jql
luJ^
-ard
lou
op suollplnru
asaqr
q8noqlle
esneJJq
'uorleurquroJJJ
Jo
sa3e1s
Jelel
aq1 ro; Lres
-sJ)au
aq.deru
slueruale
lpJelel
eql
Jo
uolletu
-roJ
aql
'srsorJru
lV'uorle8ar8es
Jruosourolql
uI
aloJ
IprntJnJls
e
Leld o1
.d1a41 are.daql'sISoIJru

sra^ossorr ot ro
(vNC
pllq^q
Jo
qJlrlls
IpnprsJr
e
qllM)
sDnpoJd JJ^ossorf,uou
laqlrJ
ol JsrJ seArB
eJnlf,nJls srql
Jo
uorlnloseJ
Jql
leql
pJunsse
uJJq
sPq
1l
'pesodord
se^^ JrnlJntls ^eprlloH
p
er^ uorleurquroJJJ
JoJ
Iapou
eql Jf,urs re^a
^
'srsotJur
Jo
sJSels
rJlel JoJ
perrnbJr
sr
pue
'SurrrBd
JuosotuoJqf,
8urnao11o;
'uorleurquof,ar
Jo
af,uenbasuor
B se
sruroy xalduor
leruauotdeuLs
aql
leqt
stsaSSns
srql
'xaldtuor
leruauoldeuLs
B
dolanap o1
pe;
osle
'rJAeMoq 'uorlPurquoJJJ
qsrloqe
lpql
suorl
-elnw'slueurquroJar
aleraua8
uer xaldurot
leru
-auoldBur(s
IpruJou
e >lJel
lpql
sluetnur auros
JSnPJJq
'r{tns
se uorlpurqurofJl
Jo} ,{ressalau
aq ot readde
tou
saop
tI'sJln)alou
tupurquro)Jr
Jo
uorlerurol
aql
Irlun
stsrsrad
tl
pup
'uorlpu
-rqurof,Jr
alPrlrur
]eql
s>lpJrq
puelts-Jlqnop
Jql
reue
suuoJ xaldruor
leruauoldeu.ds
aqt snql
'saxalduor
leruauoldeu.ds ;o
uorlPuroJ
3q1 JelJp
IUJAJ
uorlpurquoJal
Jql
;o
uorlaldruor aqt
sareld dlJealJ
srrlJ
'aua/.qred
Jo
pue
aql
le
readde
stueurqurolJJ
leqt
Moqs
o1 alqrssod
ueeq spq
1r
'q8noqt
'sJtrs
uortJutsaJ
ltsou8erp Sururetuo:
sJlnf,elou
VN11
Jo
uorl
-rnpord
Jqt
Jo
srurJt
ur Lllrarp
tseJ^
ur stueurq
-uroJJJ;o
aruereaddp
Jql Surssesse Lg
'srsoraru
;o
uorlalduoJ
eql JJUe
slupurquroJeJ
Jo
eJue
-;eadde
aql.dq
passasse
sr uorleurquroJeJ
esneJJq
'srsdeu.{.s
}o
a8Bts
eqt
lp
srnJ)o uorleurqruof,eJ
rJqlJqM auruJelJp
ot
tlnJrJJrp
uJeq seq
1I
'uorlPurquolar
Jo
sluJAJ
relnleloru aqt
aparard
ol Suurpd JrrrosoruoJqf,
roJ
pJJu
aql sluasardar
xaldruor
leurauotdeuLs
ar{l
leq]
srsoraur
Io
MJrA
IeuorlrpeJl
eql selnJer
uollpurquolau ll;nads-alrs
pue
snobolorxoH
6I
UlIdVHl
srql'saxalduoJ
IpruJuoldeu.ii.s
olur sluJruJla
IPrxP
lJa^uo)
louuP)
7uelrl.Lv)spar
eql
leql
uorl
-enJesqo
aqt.dq
pagoddns
sr
papr
srqJ
'uorteu
-rqruoJJJ
Jo
sJlelpJruJJlul
rJlel
olur uorsJa^uo)
rrJql
pue
sleerq
puens-alqnop
Jo
uorl)np
-orlur
Jql er^ uorleurqruoJJr
Jo
uorlerlrur Jql
ruoJJ
stlnsJJ
xaldruor
leuauoldeur(s
aqr;o uort
-euJoJ
ler{l
slsaSSns slue^J
1o
8urur1 J^rlelaJ
srql
'saxalduo)
Ieueuoldeuu(s
olur saurosour
-orqr
parred
Jql
Jo
uorsJJluol eql Suunp read
-desrp
,{aq1
'ruro;
sluJruale
Ietxe
ueqM
porrad
aqt Surrnp readdB
s>learq
puerts-JlqnoC
'elrs
IeuosouroJqJ
aues
Jql
le
uorssJJ)ns ur rnf,Jo saJnpnJls
lueu
-rqruof,JJ
Io
uorlpurroJ
pup
'suorDpJJ
Suured
Ienprlrpur's>leeJq
pueJls-alqnop
1eql
slsa33ns
stuJla;o
aruanbas aql
'readdesrp
DIeJJq
puprls
-Jlqnop
eqt JetJe uoos readde
'sJterpauJatur
uorleurqruorar
a,r.rlelnd JJp
q)rqm'selnJJloru
tutoI
lsrr;
aq1
'porrad
etnuru-09 e -ralo readde
-srp
uJqt
pue
readde
s>leerq
puprts-JIqnoC
':r
t"*i
"-:-*i}ltl.j
uI
PezIJeIu
-urns
sr sluenJ
;o
3unur1 JArlelJJ
etql'aatsnala)
satfwon4nas
ur srsoraur Surz[1eue ur ssarSord
uJJq spq araqt ^dltuarJJ
tnq
'uorleurquo)eJ
lp
stuJAJ
1el8o1oil.r
pue
JelnJJIou aredruor o1
a1q
-rssod
sr
U
qrrqM
ur srualsz(s loral
Jrp JJeqI
'uorleurquoJeJ
Jrlo
-rertr
IIe
s>polq aua? ap4doslte Jrtrl ur uorletnru
e
pue
'sa1o.dre1na
raq8rq
Iere^JS
ur
parJrluapr
uJJq elpq sSolouoq
11odg
'panrasuor.{lpraua8
SI UOIIEUIqIUOf,JJ
A]PIIIUI
lPql
S>IEJJq
PUEJ]S
-Jlqnop
Jqt SurlerJua8 ro; urals^ds
aqJ
'uorl
-eurqruof,JJ
luarrard
oslB
lsea,{
ur rc
alnldosotq
'aue1r\qred
Jo
pua
aq]
]P
paltalap
alp
sluPutquolai
VN0
l$un
lsLsred
pue
readde
salnrelou
lurol'saxelduor
leuauoldeufs 10
uorsuelxo
aq1 6uunp
readdesrp
pup
ulloJ sluauala
lprxp
ueqm teadde
sleerq
puells-elqno0
i
t
.*i
,:liil*1.:{
A
olercossrp
urol rurol
spue
I
I
sexelduoc
sexe;du.roc
sluor.rrola
uorlecrldet
I
--
I
leueuoldeuAg
leuleuoldeurg
lelxv
vruo
I
sluana
1ect6olop(3
I-
-
"o\i^odi
'ixii'
ostr
"btl--o;I
boi oe""'bg ov
oz
4""1"-fffifj
oueloldro
euey{qce6
auetob^Z eua1o1de1
-
-f
lstslod
Jeoooesto leeodv
sslncolou.llulof
slealqpueJls-alqnoc
ll
It
YY
sslncelouJ
slcnpotd
lueurquoceu
Ja ossoJcuoN

69'
]uepuadapul
arv uor.lprxroj
xelduol
leuauoldpufs
pue
buurpd
1'61
('parrnlo
seq uorleurquroJeJ
Irlun
sJruosorxoJqJ:raql
ale8ar8as ol
,ful
lou
op
sllJJ
leql
eJnsua o1
pren8ayes
e
saplLord
ruatsLs
srql
lpatalduror
d1n;ssarrns
uaeq seq uorteurq
-uoJer
ueqM
peurl
$ >polq
eql)
'pexnrro
tou
seq
uolleulqluo)el
JI
SISOIJIII >lJolq ol slstxa
ualsrls
,,lurod>1raqr ,,
e
pue
'srsorJru
q8norqt
ssarSord
roy Lressarau
sr uorlpurqruoJeJ
Jo
J)uJJJnJJo
eql
'eJoruJaqund 'sluelnu
urpuJJ ur
patdrustp
Lleraua8 sr
srJlossoJf,
Jo
sseuuopueruou
eql
Jsne)eq'IoJluoJ
JalossorJ
a18urs e
Jo
llnsJr
aqr
Llqeqord
st ssarord slqJ
'(
%
I'0>)
arol Lra.,l st
,rred Soloruoq
p
JoJ sJalossoJl
oraz
Jo.{1ilqeqord
aqt
tar(
'sJJAossoJ)
o^\l 01 auo.dluo sartnbre
s3o1
-ouroq;o
rrcd
qrea
'praua8
ur
'1eql
,{e.tr e
qJns
uI
palnqu$p
JJP eseql
tnq
'SJJAOSSOI)
se arnletu
,,i.[e
-nDe
suorlf,pralur;o Lluouru e L1u6
'loJluoJ
IerJ
-ua8
e o1
palqns
J]e
sluJle uollpurqruo)e5
'peqsrlqPlsJ
Jq 01 ureual uolleulq
-uroJJJ
ur suralord
JSJqI
Jo
selor
Dpxa
aq1
'(uta1
-ord
yrag
un'g
eW;o sSoloruoq are
qrrqaa)
IEpeU
pue
I)uO
pue (rredar
qJtpruslru uI
penlolur
suratord
lerJJppq
01
pJlelJJ
sr
qrq.Lr)
'HSW
epnlJur JseqJ'sa1rs elarJsrp 01
przrle)ol
eq
uP)
leq]
uorleurqruoJJJ
]sea.d
ur
pJAIOAUI
suralord
Jo
uortp)rJrtuJpr Jql u{q
paprno:d
ere
'JJAe1!l.oq
'sernlJnJls
snonurluoJsrp
Jo
uop
-eruroJ
Jqt
roJ
srseq JelnJJloru Jr.{}
otur stqStsul
'eleuserqJ
pue
sJlnpou uorlPurquroJJr
'sl
leql
'pa^rJsqo
ueeq a^Pq
leql
sarnlJnrls
alerf,
-slp
aql
qlrM
af,u3J.rnJ)o rrJr{] JlPIJJror
laz{
sp
lou
-ueJ
J,!t
lnq
'sJruosoruoJqr
)lloralu
uo slutod
alJJ)srp
le
rn)Jo slue^e uorlPulquof,au
'JlerpJruJJlur
lpntJe
aqt Surplto;d ueqt
JJqlpl
1UJAJ
UOrlPUrqrUO)aJ Jql
JO
lUeUIuJr
JUrOS
luesar
-dar,r{.aqr
reqt
slsa33ns,{.tr[qlxrlJ srr{J'seruosotu
-oJq)
aql
]o
spuJ eql
pJeMol
J^otu
Pleuselq)
eqt
'srsoreu
ur ralef
'pJ^losJJ
ueaq
ta.{ tou
seq
a8ueqrxa rpaua8 e JrJqM sJruosoruoJqr
snoSo
-louroq
uJJMlaq uorlJeuuoJ
p
sluas:;dar
lt reql
ro
'a8ueqrxa
palayduor
e
Jo
runnprsar
Jql stuJs
-ardar
tl tpqt
alqrssod sr
U
'u,uou>lun
sr erusplqJ
p
Jo
aJnleu JelnJalou eql
lnq
'a8ueqrxa
rrlaua8
e
Jo
eluJrJnJJo er{l
alpJrpur
o1
parunsard uaaq
seq srql'pJ])euuo) ere saurosouroJqJ
Jql I{lIq,&\
te
slurod se Jlqrsrn eruoJJq ueql etpurserqr
Jql
lxaldruor pruauotdeu.ds
aqt
paqs
seuosouorqJ
aqt
'(auaroldrp)
srsoraru;o a8ets
txau
rql
lV
'uollnlosJJ
pue'stsaqluzls
y1qq
'aBueqJXJ
pueJts
Jo
uorsualxe eql
J^lo^ul
qJrqm
'uorleurqruoJar
yo
sdals
luanbasqns
aqr
ziq
pardnno
sr
porrad
srql
teqt
aq
r(eru
tI'pelJJsqo
JJe slueurqruoJal
vNo
eJoJeq
'aua1z{qred
Surrnp suaddeq z(1ensn
ter{,lr
reelJ
tou
sl
1I
'y1qq
snoSolouoq
qtlM s>leerq
JSJqI
Jo
uollJpal JAISUJ]xJ
true alnbar
lou
saop
tI
's>leeJq
pueJls-elqnop
sarrnbar
xaldruor
pruauoldeuz(s aql;o uoueruJoJ
JI
'lBql
stsaSSns
srql'parredal
lou
aJe
daqr
rnq'sluplnlu
Tdoqp
saxaldruor
pruauoldeu,{s aqt uI saluosoluorqJ
parredsnu
aql ul
turoJ s>lpaJq
pueJls-elqno(
'burllPrJlur
uorJ
saruosouroJqr
snoSoloruoquou
luaaard
ol st
Tdog
yo
JIor
Ipnsn
aq1
'(satuanbas
y1qq
yo
uost
-redruor
alrsuJlxe
,{ue uo
pJseq
eq
louuef,
aJoJ
-JJJqt
pue)
LSoloruoq
Io
luepuedaput
st
qrns
se
saxaldruor
pruauoldeuLs
yo
uorleruJoJ aqt
teql
slsa33ns
srqJ'seruosoruoJqJ
snoSoloruoquou
ureluoJ
saxaldruor
IpnpIAIpuI
eql
lnq'stsoraru lB
ruroy
xalduror
leurauoldeuLs
;o
slunotu€
IeIu
-Jou
'stuelnu
7dor4
uI'aasnaln
'g
ut
7dot1
aua8
aqt [q
pallortuoJ
sI sauosoluoJql
snoSolouroq
ueamtaq
uollplf,ossp
;o
,hnrltrads_ aqJ
'uorlPruJoJ
xeloruo)
IPIu
-auotdeuz(s
Jo uorleulqluo)eJ
Jo
luapuadapur
sr
sSolouroq
uaJMlaq
uotltuSotar
snql
'saxald
-ruol
lpueuoldeuz(s
uroJ
lou
op
daqr
1nq
'rred
01 saurosouroJql
s^l.ollP uollPlnur
zdlz
JqI
'raqto
eql Surnage
lnoqllm
sassarord aq]
Jo
auo
s>lJolq
qrlqM
Jo
qleJ
'suollelnru
oA1.1
Jo
spJJJe
aqt.dq
uorleruroy
xaldruor
pruauoldeur{s
pue
Surrred;o sassarord
aql
qsln8upslp UPJ JM
'ssalo.td
raqlo
aql 6uLpa11e
lnoqlm
uotlpuroJ
xelduor
leLuauoldeur\s
to
6uuLpd auosotttorql
laqlte
u! lnllo
uel suo!]Plnfl
o
luepuadepul
arv
uo$eurol
xald
uol
lPuauoldeu^5
puP
6uutPd
.SIJAOSSOJJUOU
Jo
uorllnpord
aqr
JoJ alnor
Jaqlo auos
Jq
Leru
araql
lpql
lnq-selnJJlotu
luro[
1o
uol]nl
-osar
,{q-tq8noqr
dlsnonard
se
parnpord
are
srelossorJ
leqt
s1sa33ns
3unu1l
ut ,{.ruedanstp
eqJ
'erurl
arues
Jql
te
readde s1
ruJl{l
Dadxa
plnoM
a.tt.
lJle,lroq
'ssarord
uollnlosJJ
Jrues
aqt
,{q
parnpord
eJeM
lJnpord
1o
sad,(r
qloq
JI'(€I'6I
arn8rg
aas) salnraloru
turof
aql
qlllrt
.{.lsnoauellnurs
lsorule
readde
srelossoJf,uou
seJJaqM
'selnJeloru
luro[
;o
aruereadde
lsrr;
Jqt
JeUe
IIJM
lllun
readde
tou
op sJJAossoJ)
'anrl
Jq
1ou
,{eru slqr
leql
lsaSSns
'JJAaMoq
'seln)alolu
Ja^ossoJJ
pue
JJnossoJJuou
Jo
uop
-rnpord
Jo
serull
Jql
Jo
slueueJnseJru
lueleg
'(g'61
arn8r4
aas)
uollnlosJJ
uI
paAIoAuI
eJP
spueJts
rplq,lr
uo
Surpuadap
'(slueutqurola:)

'UOIIPUIqUOJJI
JO}
AIEJISqNS
E SI SIqJ
'AJUJNDAS
lLp
aql
lP
puJ
,€
P
qllM
vNo
pJpupJls-JlSurs
Jle
-Jaue8
01 sI uoll)pJelur
slql
Jo
llnsJJ
IIEJJ^O
JqJ
'peeds
snor^aJd
er{t
JIpq
lnoqp te-uorlp)olsueJl
J^rrp o1
lFnqns
g)eu
aql ^Iuo
Sursn Mou-ese)
-llJq
e sP uorpunJ
01 tJ^JMoq
'senurluoJ
tJ'^1r
-Arpe
aseJlJnu
slr sJsol
Ju^zuJ Jql
lurod
qlrqM
tP
'pJlP^Ipeur
Jruo)aq
ro
JlprJossrp ol
llunqns
(JaU
eqt sJSnpJ
et;S
lLp
aq1
Jo
uonru8onp'
u6t
1o
rq8rr
aql
01 saseq xrs
pue
rnoJ
uaeutJq
uoqrsod
e
]e
vN(
eql
Jo
puPrls
dot aql sr^eap
ueql
1I
'asned
ot arudzua
Jqt sasnef,
srqJ'uJoJ
papueJls
-a18urs
ur
el;rs tq) eql
Jo
pueJts
do1 aqr sazruSotar
u
'Jlrs
!ry
etqt saq)eJr
1l
ueqM
'puJ
,€
aql
qlIM
pueJls
a13urs
pasealal
aqt saper8ap
tr
'se)uelpe
61)g)a5
sy
'trunqns
OJJU
rql ,{q
uanrrp
z(leqrur
sr xrlaq
Jlqnop
aql Surpur^,vrun
pup
vNC
Suop uorlerolsue.{l',{1rre1od,S-,€
qlrm
suorpunJ
gJJU
pue
,{1rre1od
,€-,5
rtrfIM suortJunJ
OJaU
:serlrnrlJp
Jse)laq
JApq slrunqns
slr
Jo
o^41
'pua
prpue[s-elqnop
p
lE
vN(
01 spurq
cf,gJeu
'a]rs
t4)
e seq
leqr
VNCI
JlErlsqns
p
uo
peleurpJooJ
aJe suorl)pJJ
3saql ^loq
sMoqs
?I'6I
lU{l5H
'puJ
,€
JJIJ
p
qtrM
uor8ar
papue,rls-a13
-urs
e
aprnoJd ol
Jq Leur
uorleurqtuoJal
ur JIol
stl',{.lr,rrpe rspdJv
up
seq
tl
pue
'(gSS)
uratord
Surpurq
puerls-a13urs
e
yo
aruasard
er{l ur
vN(
xaldnp
purMun
ueJ
teql
sartrlrlJp
Jsetrleq
seq
u A
aseJlJnuoxa.r{.]r,rrlre
Jql
se
pJrJrluapr,{.11eu
-r8rro
se,lr
q)lrlm
'ypq
saper8Jp
tpql
asealf,nu
luJlod
P sI
1I'sJlllnrlJp
IpJJAes
sasrlraxa xald
-ruoJ
srqJ
'e)pal
saua8
aql z(q
papol
aru.dzua
ue
Jo
uorlJe Jql
JoJ sla8rel
Jrc
seTrs nl)
'uorlJJrrp
Juo ur
z(po xaldnp
aqt
3uo1e
enoru up)
ueqt
pue
,pua
uJ>loJq
e
le
VN(I
qll,lr
elerJosse
lsnru
snte:edde
uorleurquoJeJ
eql
leql nsaSSns
uorleluerJo
uo
aruapuadap
sq;'(anoqe
uMoqs
aruenbas aqr
;o
lq8rr
rqt
01) Jprs
teln)rued
auo
uo .dea,re
q1
Iprelrs
epeu >leJrq puerls-rlqnop
e ,{q
pare,r
-lne
eq
ueJ rtrs
u,p
v
'atrs
rqt urorJ qI
0I
ol dn
Jo
a)uelsrp
p
urqlrm
Les
z(lrurtt,l.
leraua8
slr ur
uortpurquoJJl
sJlplnrults
a:uanbas t4.r
y
.IIOIIPUIqIuOJEJ
JOJ
IPI]UESSE
lou
sr
ll leqt
sMoqs
'sluJruale
rrlaua8
raqlo
ruorJ osle
pue
'1trN11
epqruel
adLl-p1ru.
ruo,r1
aJuasqp
sll
'q{
0I
ol
E
.dra,ra
aluo
tnoqe
17Nq
tln'g
ur Llernleu
sJnJJo
a)uenbas
ltp
erl1-
,E
))Vf,)V9),€
,€
DDJ9CJf,',S
:dq
g
yo
aruanbas
1et
-rrlarurudsuou
luplsuoJ
e eJeqs
selrs asaqJ
'uoIleurqtuoJeJ
ur
pJAIoAUI
suralord
raqlo
Jo
JIoJ
eql ot sn
peel
sJlrs
eseqJ'uorleurqruoJeJ
alplnrulls
leql
saruanbas
uorleurquolaX r4oad5-e1Ls
pue
sn060louloH
6I
UlldVHl
ateerJ
leql
saSueqr rred-aseq
a18urs aterT'n1t
pJIIeJ
sluplnru
Io
ruroJ Jqt ur
ppquel
a8eqd ur
peJeloJsrp
aJaM rprr{,lr'slodsloq eseqJ'uouernq
-uroJar
alplnturls
teql
stodstoq urpuJl
Jo
eJuel
-sIXe
aq1
Jo
tlnseJ
P se
paJa^of,srp
uaaq sEq spue
elqplrns SurtBrauaS
roJ rusrueqJeru JUO
'paleraua8
aq
lsnu
spue
,€
pup
suot8ar
papuerrs-a13urs
,rana,lroq'(sa1o,{.re1na
ur srsoraur
1e
uorleurquoJJr ur
se) saln;a1oru
xaldnp
ora]
3ul^lo^ul saf,uelsurnJrlJ uI
'JIJTIJ
3ur11or e z(q
uorletrldar SuroS.rapun saruoua8
a8eqd,{q
pareraueS
aq,{eru slpt
papupJts-apurs
ro
la8eruep
uorlprppJJr
^dq
paleraua8
aq .{eru
sdeS
papuerls-a18urs
:(VNq papuerlg-a18urg
srelsup{l uorleSnfuo3
'0I'9I
uorlJas JJs :uorl
-e8nfuor
ur se) ruroy
papuerls-a18urs
ur
peprl
-ord
aq
z(eru
y1r1q
:spuJ
,€
papuerls-afurs
aJJJ
qllM
elqplre^e
aq
z(eru
VN(
'saseJ
aruos uI
'salo
-fue1ord
ur uoneurquroJJJ Jlerlrur
ol salnoJ snor
-:en
aq,{eur
aJeqt
tnq
'y1trq
xaldnp
Jo
stunourp
a8rel
a8ueqrxa Lllensn
tou
op
prJalJpg
'pelJ4uepr
uarq a^eq
oo1
,{1uau1
0l ual Jrxos
'uorleurquoJJJ
pezrleJa
-ua8
a>leuapun
o1 zfuqqeur Jqt sr
sluetnu--ral
]o
addtouaqd
aq1
'saua8
JrJql ur suorlplnru
-ral
Jo
eJueJJnJJo
eql ^q
perJrtuepr
uJaq
an€q uorl
-purqruoJal
ul
palpl[drur
saru,{.zua
IprJeDpg
'uorleurquoJar
JoJ
pa;rnbar
stuauoduroJ
aq1
Jo
aruos .d1uo
luasardar
,{.1q
-eqord
z(aqt
q8noqlle
'u,lrou>l
are a8els
qrea
ur
panlolur
saurz(zug
'saxaldnp
rauued
aql
sJAIoseJ
aspal)nuopue
up
pup
lpepuJlxJ
sr
Surrred;o uor8ar
aqt
txaldnp
rautred
e
qlrM
slJpJJtur elnJeloru
ua>loJq e ruorJ
puerls
a13
-uls
17
:q8noql
'relrurs
sr stuJAJ relnJeloru
Jo
rJpJo
IerJueS
aql
'saruosouroJqJ
pelur
ueql
raqleJ
selnJJloru
VNO
Jo
suor8ar
pJt)lJlsaJ
sa^lo^ur pup
rusrueqJsru uorleurqruoJaJ
aql
Jo larred
pue
lred
sr uorlJpJJ
uollluSoJaJ
Jql eJaH
'srualsds
IerJJpeq
ur
peqrJJSep
lsJrJ
seM sJlnJeloru
yNO
uJJMlaq
saruanbas
yo
a8ueqrxa
ur
pa^lo^ur
stuela
eql
Jo
eJnteu
aqJ
'Alnqre
aseallnu
pue
lrunqns
6rau
eql
Jo
ssol sra66u1 elrs t{, eq1
.
.olls
lq,
aql 01 se^our
1!
sP
,g_,€
urol]
puplls
auo saperbap
pue
'xeldnp
aql spulmun
'eluanbas
ry)
e
to
upallsuMop
VNo
ol spulq
lI
.
'sarlt^rllP
asPlnaq
puP
asPallnu seq xalduol
Ol8reu
eql
.
saruanbas
lrp
frq
patPlnu[s
{
uals^s
OL'
ol8lau
lPuatlPB
aql