Hugo W.B., Russel A.D.(ed). Pharmaceutical Microbiology
Подождите немного. Документ загружается.

organs, e.g. lymph nodes, spleen, Peyer's patches (which are collections of lymphoid
tissue in the submucosa of the small intestine) and the tonsils.
A large number of cells are involved in the immune response and all are derived
from the multipotential stem cells of the bone marrow. The predominant cell is the
lymphocyte but monocytes-macrophages, endothelial cells, eosinophils and mast cells
are also involved with certain immune responses. The two types of immunity (humoral
and cell-mediated) are dependent on two distinct populations of lymphocytes, the B
cells and the T cells respectively. Both the humoral and the cell-mediated systems
interact to achieve an effective immune response.
4.1 Humoral immunity
Humoral immunity, known as antibody-mediated immunity, is due directly to a reaction
between circulating antibody and inducing antigen and may involve complement. The
B cells originate in the bone marrow. In chickens, a lymphoid organ embryonically
derived from gut epithelium and known as the bursa of Fabricius is responsible for the
maturation of the B cells into immunocompetent cells, which subsequently can
synthesize antibody after stimulation by antigen. The bursal equivalent in humans is
the bone marrow itself. An antigen (e.g. a bacterium) may possess multiple determinants
(epitopes) and each one of these epitopes will stimulate an antibody which will
subsequently react with that epitope and with closely related epitopes only. Each B cell
is only capable of recognizing one epitope via a specific receptor on its surface. This
receptor has been shown to be antibody itself. Activation of the B cell occurs by binding
of the antigen to the receptor and the resultant complex is endocytosed. For activation
to proceed, additional signals are now required.
These are supplied by the secretion of peptide molecules (termed cytokines or
lymphokines) from a subset of the T-cell family (the helper T cells, TH cells). These
peptide molecules (interleukins (IL) 2,4,5 and 6) stimulate the B cells to proliferate,
undergo clonal expansion and mature into plasma cells which secrete antibody and
also into the longer-living, non-dividing memory cells.
Antigens requiring the assistance of TH cells are termed T-dependent (TD) antigens.
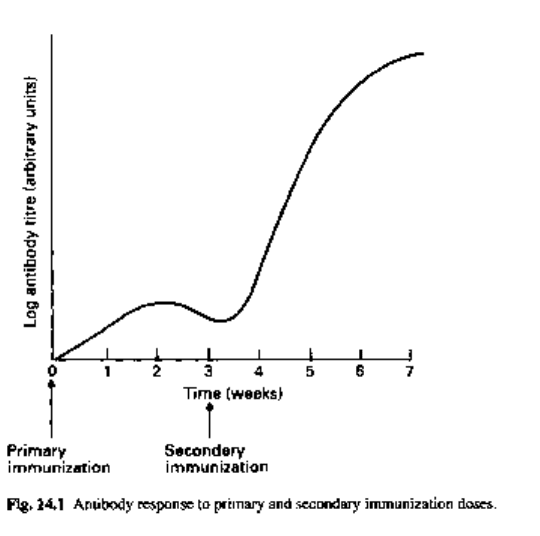
Subsequent antigenic stimulation results in high antibody titres (secondary or
memory response) as there is now an expanded clone of cells with memory of the
original antigen available to proliferate into mature plasma cells (Fig. 14.1).
Some antigens, such as type 3 pneumococcal polysaccharide, LPS and other
polymeric substances such as dextrans (poly-D-glucose) and levan (poly-D-fructose)
can induce antibody synthesis without the assistance of TH cells. These are known as
T-independent (Ti) antigens. Only one class of immunoglobulin (IgM) is synthesized
and there is a weak memory response.
Immunoglobulins are associated with the y-globulin fraction of plasma proteins
but, as stated earlier, not all immunoglobulins exhibit antibody activity.
The immunoglobulin (Ig) molecules can be subdivided into different classes on the
basis of their structure, and in humans five major structural types can be distinguished.
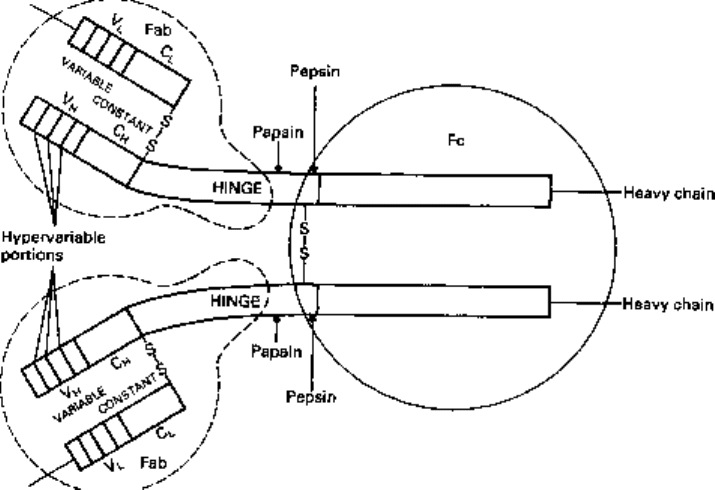
Each type has been distinguished on the basis of a polypeptide chain structure consisting
of one pair of heavy (large) chains and one pair of light (small) chains joined by
disulphide bonds. The heavy chains are given the name of the corresponding Greek
Fundamentals of immunology 285
letter (/chain in IgG, fi in IgM, a in IgA, 5 in IgD and e in IgE). All classes have
similar sets of light chains consisting of one of two types, the kappa (K) or lambda (A.)
chains. A suggested ground plan for the most abundant Ig, IgG, is illustrated in Fig. 14.2.
IgG consists of four polypeptide subunits held together by disulphide bonds. Native
immunoglobulins are rather resistant to proteolytic digestion but certain enzymes have
been useful in elucidating their structure. Papain cleaves the molecule into three
fragments of similar size:
1 two Fab fragments each carrying a single antigen-combining site and comprising
the variable regions of both chains, the constant region of the light chains and the first
constant domain of the heavy chain;
2 one Fc fragment composed of the terminal halves of the heavy chains which have
no affinity for antigen but can be crystallized.
Cleavage with pepsin yields two fragments only, one consisting of two Fab fragments
and the other an Fc fragment which is partially degraded by the enzyme. The variable
regions on both the heavy and light chains contribute towards antigen recognition,
whilst the constant regions of the heavy chain, particularly the Fc part of the heavy-
chain backbone, direct the biological activity of the molecule, e.g. complement fixation
(see later) and the interaction with a variety of tissue cells, via membrane receptors for
the Fc region.
Intrastrand bonding via disulphide links cause the molecule to fold into 'globular
domains' and it is these that direct the biological activity of the molecule.
4.2
286 Chapter 14
Monoclonal antibodies
After antigenic stimulation, the normal antibody response involves the activation of a
Light chain
Light chain
Fig. 14.2 Diagrammatic representation of IgG.
large number of clones of antibody-secreting cells (i.e. it is polyclonal). This is due to
the fact that antigens possess multiple epitopes. In 1975 Kohler and Milstein successfully
developed cell fusion techniques which enabled them to isolate clones of cells which
synthesized identical antibody molecules (Fig. 14.3).
The principles of the technique rely on the fact that an antibody-secreting cell can
become cancerous and the unchecked proliferation of such a cell is called a myeloma.
Progeny of the original transformed cell will continue to secrete a single kind of antibody
molecule only. Myeloma cells, like other malignant cells, grow indefinitely in tissue
culture. However, the specificity of the antibody is unknown. Mutant myeloma cells
have been isolated which have lost the ability to secrete antibody while still retaining
their cancerous growth properties.
Mouse myeloma cells are fused with an antibody-secreting cell from the spleen of
a mouse immunized with the required antigen. The technique is called somatic cell
hybridization and the resultant cell is termed a 'hybridoma'. The rate of successful
hybrid formation is low and a technique is necessary which can select these successful
fusions. The standard technique is to use a myeloma cell line that has lost the capacity
to synthesize hypoxanthine-guanine phosphoribosyl-transferase (HGPRT). This enzyme
enables cells to synthesize nucleotides using an extracellular source of hypoxanthine
as a precursor. The absence of HGPRT is normally no problem as cells can use an
alternative pathway. When, however, these cells are exposed to aminopterin (a folic
acid analogue; see Chapter 8) they are unable to use this other pathway and become
fully dependent on HGPRT.
Fundamentals of immunology 287
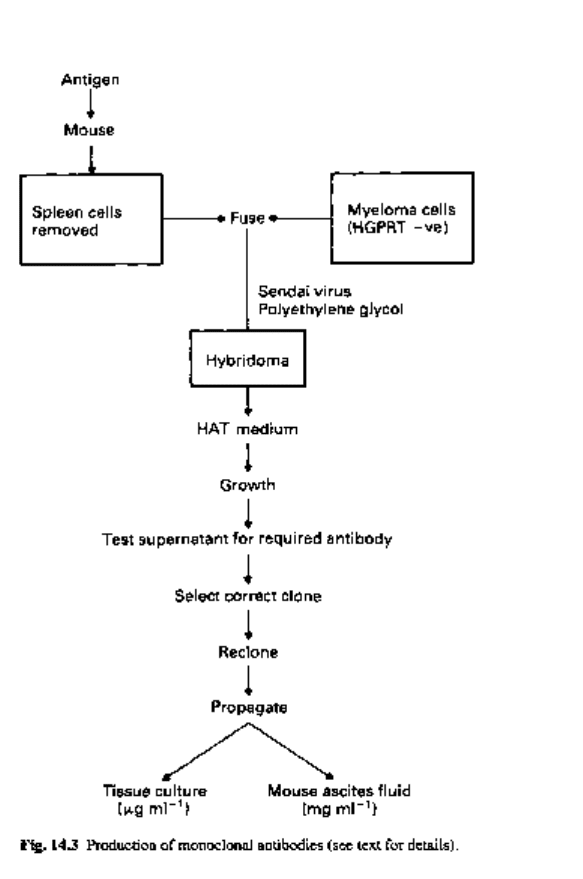
The cell fusion mixture is transferred to a culture medium containing hypoxanthine,
aminopterin and thymidine (HAT medium). Unfused myeloma cells are unable to grow
as they lack HGPRT. Unfused normal spleen cells can grow but their proliferation is
limited and they eventually die out. The hybridoma cell can proliferate in the HAT
medium as the normal spleen cell supplies the enzyme which enables the hybridoma to
utilize extracellular hypoxanthine.
The hybridoma is now screened for the production of the desired antibody by testing
the supernatant from each culture. A single culture, even though positive for antibody
production, can contain the progeny of two or more successful fusions. Therefore, it is
necessary to dilute positive cultures so that fresh cultures can be started with a single
hybridoma cell. When successful, such cultures are truly monoclonal and the antibody
is directed against a single epitope on a preselected antigen. Once established, these
cell lines are immortal.
The concentration of antibody in tissue cultures of the hybridoma is low (10-
60^gml
_1
) but the use of large culture vessels can obviate this. The hybridoma can
also be propagated in mice where the antibody concentration in the serum and other
body fluids can reach lOmgml
-1
.
4.2.1 Uses of monoclonal antibodies
Monoclonal antibodies are very sensitive, specific reagents and have applications in
many areas of the biological sciences. They revolutionized immunology within a few
years of their discovery.
The investigation and characterization of cell surfaces by probing with monoclonal
antibodies is one of the most vital areas of application. In this context they have been
used in the following ways:
1 To study the ABO and rare blood groups.
2 To detect HLA antigens and consequently to type tissues for transplantation.
3 To classify cell lines, e.g. the T-cell subsets, and thence to separate these cell
subpopulations.
4 To study cell-cell interactions and differentiation, e.g. embryology.
5 In oncology, to study the relationship between the normal and the tumour cell, to
detect tumour-associated antigens (CEA, carcino-embryonic antigen, and AFP, a-
fetoprotein) and subsequently to enable cancer therapy to be monitored, to locate tumour
metastases, and to deliver cytotoxic drugs, toxins, radionuclides, or liposomes to tumour
cells.
6 To identify and characterize bacterial and viral antigens which can then be purified
and used to prepare subunit vaccines.
Monoclonal antibodies have further been employed for studying drug and hormone
receptors, enzymes and proteins. A whole range of immunoassay techniques using
monoclonals have been developed to detect low levels of materials in body fluids, e.g.
oxytocin can be detected in human blood using a radioimmunoassay down to 1 pmol I"
1
.
Similar assays are used to monitor antibiotic therapy using potentially toxic drugs, e.g.
gentamicin. The future of monoclonal antibodies continues to be one of enormous
potential and excitement.
4.3 Immunoglobulin classes
The synthesis of antibodies belonging to the various classes of immunoglobulin proceeds
at different rates after the initial and subsequent antigenic stimuli.
4.3.1 Immunoglobulin M (IgM)
Synthesis of this class occurs after the primary antigenic stimulus. IgMs are polymers
of five four-peptide subunits and have a theoretical valency of 10, although against
large antigens such as bacteria their effective valency is five. They are extremely ef-
fective agglutinating agents and, as they are largely confined to the bloodstream and
they appear early in the response to infection, they are of particular importance in
bacteraemia.
Fundamentals of immunology 289
Serum concentrations lie between 0.5 and 2.5mgml
_1
. IgM can fix complement
and a single molecule can initiate the complement cascade. IgM (with IgD) is the
major immunoglobulin expressed on the surface of B cells where it acts as an antigen
receptor.
4.3.2 Immunoglobulin G (IgG)
This is the major immunoglobulin synthesized during the secondary response and in
normal human adults is present at serum concentrations between 10 and 15mgmH.
Within this class there are four subclasses, designated IgGj, IgG
2
, IgG
3
and IgG
4
.
It has the ability to cross the placenta and therefore provides a major line of defence
against infection for the newborn. This can be reinforced by transfer of colostral IgG
across the gut mucosa of the neonate. It diffuses readily into the extravascular spaces
where it can act in the neutralization of bacterial toxins and can bind to microorganisms
enhancing the process of phagocytosis (opsonization). This is due to the presence on
the phagocytic cell surface of a receptor for Fc.
Complexes of IgG with the bacterial cell activate complement with the resultant
advantages to the host.
4.3.3 Immunoglobulin A (IgA)
This occurs in the seromucous secretions such as saliva, tears, nasal secretions, sweat,
colostrum and secretions of the lung, urinogenital and gastrointestinal tracts. Its purpose
appears to be to protect the external surfaces of the body from microbial attack. It
occurs as a dimer in these secretions but as a monomer in human plasma, where its
function is not known. The function of IgA appears to be to prevent the adherence of
microorganisms to the surface of mucosal cells thus preventing them entering the body
tissues. It is protected from proteolysis by combination with another protein—the
secretory component.
It is present at serum levels between 0.5 and 3mgmH but higher concentrations
are found in secretions. There are two subclasses of this immunoglobulin.
4.3.4 Immunoglobulin D (IgD)
This occurs in normal serum at very low levels (30-50 fjg ml
-1
) but is the predominant
surface component of B cells. Immature B cells express surface IgM without IgD but
as these cells mature IgD is also expressed. After activation of the B cells, surface IgD
can no longer be detected and it would appear that IgD may be involved with the
differentiation of B cells.
4.3.5 Immunoglobulin E (IgE)
This is a very minor serum component (0.1-0.3 fig ml
-1
) but is a major class of
immunoglobulins. It binds with very high affinity to mast cells and basophils via
a site in the Fc region of the molecule. Crosslinking of the cell-bound IgE
antibodies by antigen triggers the degranulation of these cells with the release of
290 Chapter 14
histamine, leukotrienes and other vasoactive compounds. This class may play a role
in immunity to helminthic parasites but in the western world it is more commonly
associated with immediate hypersensitivity reactions such as hay fever and extrinsic
asthma.
Humoral antigen-antibody reactions
Antibody molecules are bivalent whilst antigens can be multivalent. The resultant
combination may result in either small, soluble complexes, or large insoluble aggregates,
depending on the nature of the two molecules in the system. The following are examples
of the reactions that can occur.
1 Neutralization. Small soluble complexes neutralize microbial toxins.
2 Precipitation. The formation of insoluble precipitates which enable the phagocytes
to eliminate soluble antigen from the body.
3 Agglutination. The aggregation of bacterial cells into agglutinates enabling
phagocytes to eliminate these cells rapidly from the body.
4 Cytotoxic reactions. The antibody and cell react, with resultant lysis of the cell. It
was found that the presence of a third component, called complement, was necessary
for this reaction to take place.
Complement
Complement activity was first recognized by Bordet, who showed that the lytic activity
of rabbit anti-sheep erythrocyte serum was lost on heating to 56°C but was restored by
the addition of fresh serum from an unimmunized rabbit. Thus, two factors were
necessary, a heat-stable factor, antibody, plus a heat-labile factor, complement, which
is present in all sera.
Complement is not a single protein but comprises a group of functionally linked
proteins that interact with each other to provide many of the effector functions of humoral
immunity and inflammation. Most of the components of the system are present in the
serum as proenzymes, i.e. enzyme precursors. Activation of a complement molecule
occurs as a result of proteolytic cleavage of the molecule, which in itself confers
proteolytic activity on the molecule. Thus, many components of the system serve as
the substrate of a prior component and, in turn, activate a subsequent component. This
pattern of sequential activation results in the system being called the 'complement
cascade'.
Complement can be activated by two pathways, the classical pathway and the
alternative pathway (Fig. 14.4).
The classical pathway
The first component of complement is CI. This is a complex of three molecules
designated Clq, Clr and Cls. The classical pathway is only initiated by an immune
complex (antibody bound to antigen) when Clq binds to the Fc portion of the complexed
antibody (IgM or IgG). The binding of Clq activates the Clr and Cls molecules
associated with it to yield activated CI which now cleaves C4 and then C2 (subunits of
Fundamentals of immunology 291
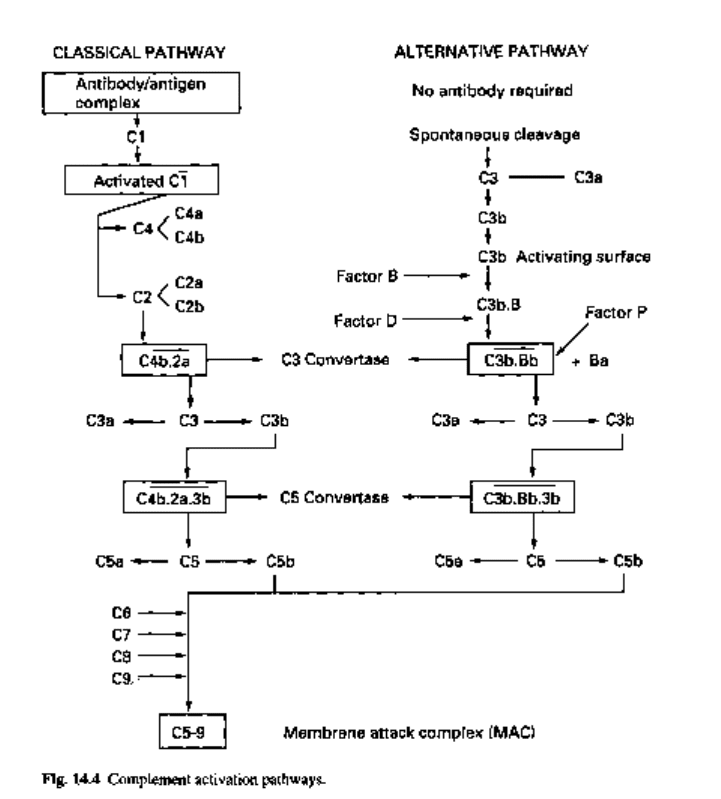
complement that possess enzymatic activity have a bar over the subunits name).
Cleavage of C4 yields a small fragment (C4a) and a large fragment, designated C4b,
which binds to the cell surface near the CI molecule.
Cleavage of C2 yields two fragments: a larger one, C2a, and a smaller one, C2b.
C2a binds to a site on C4b to yield C4b2a, which is a C3 convertase as it can now
cleave C3 into C3a and C3b. C3b is bound to the C4b2a complex to yield C4b2a3b.
This complex is enzymatically active against C5 and for this reason is described as a
C5 convertase. C5 is cleaved into C5a and C5b; it is the latter molecule that serves as
a locus for the assembly of a single molecule each of C6, CI and C8. The resulting
C5b.6.7.8 complex allows the polymerization of C9 into a tubular hydrophobic structure
that is inserted into the lipid bilayer of the cell membrane which forms a transmembrane
channel through which ions and small molecules are able to diffuse freely. This structure
is termed the membrane attack complex (MAC).
C3a and C5a are released into the fluid surroundings where they serve as potent
anaphylotoxins in that they cause vasoactive substances such as histamines to be released
from mast cells and basophils. C5a is also strongly chemotactic for neutrophils.
292 Chapter 14
Free C3b fragments bind to the surface of the target cell. There are specific receptors
for membrane-bound C3b on polymorphs and macrophages. This allows immune
adherence of the complexes to these cells, thus facilitating subsequent phagocytosis.
While antibodies alone bring about phagocytosis of antibody-coated particles through
Fc receptors that are also found on phagocytes, the presence of C3b markedly enhances
the phagocytic process.
4.5.2 The alternative path way
The cleavage of C3 and the activation of the remainder of the complement cascade can
be triggered, in the absence of complement-fixing antibody, by agents such as bacterial
polysaccharide. C3 in the serum cleaves spontaneously and the C3b generated is rapidly
inactivated (factors I and H). However, C3b bound to the surface of many microbes is
able to bind to a serum protein designated factor B, which is now, in turn, cleaved by
another serum protease, factor D. The resulting complex, C3b.Bb, is stabilized by another
protein called/?roperdein (P). The resultant stable complex, C3b.Bb, is a C3 convertase,
analogous to C4b.2a. It cleaves C3 to form a multimolecular complex, C3b.Bb.3b,
which is a C5 convertase and can generate C5b which is the focal point for the assembly
of the MAC.
Proteins B, D and P also amplify the effects of the classical pathway in that some of
the 3b generated by this pathway interacts with these proteins to form additional C3
convertase that supplements that provided by C4b.2a. Likewise, enhanced cleavage of
C5 occurs due to the dual activity of C4.2a.3b and C3b.Bb.C3b complexes.
4.5.3 Regulation of complement activity
The spontaneous generation of C3b creates the potential for the triggering of the entire
complement cascade. Two regulatory proteins prevent this. Factor I inactivates C3b
unless it is bound to a surface. This action is enhanced by factor H, which also removes
Bb from the C3b.Bb complex, thus inactivating the C3 convertase. The classical pathway
is also under regulatory control as activated CI could theoretically continue to cleave
C4 and C2 molecules until they were entirely consumed. The presence in the serum of
a CI inhibitor (CI INH) prevents this by binding to activated CI, allowing only a brief
interval during which it can cleave C4 and C2 before it is deactivated by CI INH.
Complement plays a significant part in the defence of the body. It can cause lysis of
Gram-negative organisms by allowing lysozyme to reach the peptidoglycan layer of
the organism. The generation of the C3b complex on the surface of the cell facilitates
phagocytosis as the phagocytes possess a receptor for C3b, whilst C3a and C5a cause
the release of histamine with the resultant increase in vascular permeability increasing
the flow of serum antibody into the infected area. C3a and C5a also attract phagocytic
cells to the focus of the infection.
4.6 Cell-mediated immunity (CMI)
The term cell-mediated immunity is used to describe the localized reactions that
occur to those microorganisms that have the ability to live and multiply within the
Fundamentals of immunology 293
cells of the host, e.g. the tubercle bacillus, viruses and protozoal parasites. These
reactions are mediated by lymphocytes and phagocytes and antibody plays a subordinate
role.
When immunologists recognized that there were different classes of lymphocytes
that were functionally and developmentally different, attempts were made to develop
methods to distinguish them. This was initially done by raising antibodies to the cell
surface proteins using animals of a different strain or type, i.e. 'alloantibodies'. The
advent of hybridoma technology allowed the production of monoclonal antibodies
that reacted specifically with defined populations of lymphocytes via cell surface
molecules which acted as antigens (markers). Some of these markers are specific
for cells of a particular lineage, whereas others indicate the state of activation or
differentiation of the same cells. Thus, a marker that is recognized by a group ('cluster')
of monoclonal antibodies is called a member of a cluster of differentiation and given
a 'CD' designation.
The lymphocytes involved in CMI originate from the multipotential stem cell and
are processed by the thymus gland; hence the name 'T cells. The role of the thymus is
to rearrange the genes associated within the T cell receptor (TCR) so that the mature T
cells recognize foreign but not self antigens. This receptor has been isolated using
monoclonal antibody probes and has been shown to consist of two disulphide-linked
polypeptide chains termed the a and (3 chain. This receptor is associated with a
characteristic cell surface marker, CD3. Antigen recognition occurs via this membrane
structure CD3/TCR. Mature T cells also express other antigenic markers, notably CD4
or CD8. Thymectomized neonate mice do not exhibit the CMI response indicating the
importance of the thymus gland.
Infection with a human immunodeficiency virus (HIV-1 and HIV-2; see Chapter 3)
can cause the destruction of the TH cell, which is the critical cell of the immune system.
This leads to the condition known as acquired immune deficiency syndrome (AIDS).
At present, it is still not known why, in some cases, infection with HIV leaves the
immune system intact whereas in others it is irreversibly destroyed, giving rise to AIDS.
The immune system must be able to distinguish between antigens against which an
immune response would be beneficial and those where such a response would be harmful
to the host, i.e. it must be able to distinguish between 'self and 'non-self. This is
achieved via molecules of the major histocompatibility complex (MHC). The human
MHC is located on chromosome 6 and is known as the HLA (human leucocyte antigen).
It is divided into four main regions, designated A, B, C and D. Products of this region
are expressed on the surface of cells and these enable cells of the immune system to
recognize and signal to each other. Three main groups of these molecules have been
identified.
1 Class 1 MHC molecules are integral membrane proteins found on the surface of all
nucleated cells and platelets. They are the classical antigens involved in graft rejection.
2 Class 2 MHC molecules are expressed on the surface of B cells, macrophages,
monocytes, various antigen-presenting cells (APCs) and certain cells of the T-cell family.
3 Class 3 MHC molecules consist of several complement components.
T cells only respond to protein antigens when the antigen has been processed by
the APCs. The resultant small peptide molecules are then bound to the Class 2 molecules
on the surface of the APCs. Monocytes, macrophages, B cells, dendritic cells and some