Хрисанфова Е.Н., Перевозчиков И.В. Антропология
Подождите немного. Документ загружается.

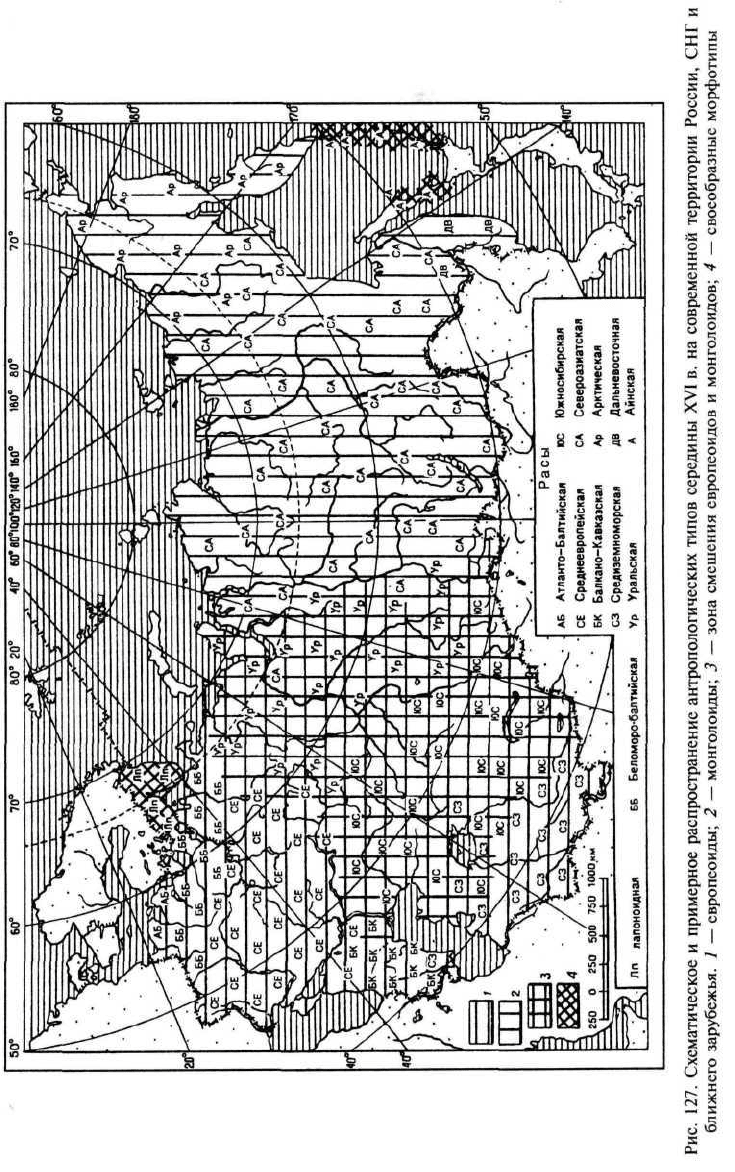
смешения. К раннему средневековью практически все население данной
зоны стало смешанным, но при значительной вариабельности в проявле-
ниях особенностей исходных вариантов у отдельных индивидов в чистом
виде они не встречаются (по крайней мере, в пределах ошибки методов).
Такая же ситуация сохранилась и в современном населении. Весь процесс
занял около 1000 лет, или 50 поколений из расчета 20 лет на поколение.
Монголоидные особенности в этом населении преобладают, так как на
протяжении последних полутора тысяч лет направление генного потока
шло из монголоидных групп более восточных районов, но наличие зна-
чительного процента смешанных оттенков глаз, ослабленность монголо-
идных особенностей в области глаз, некоторое усиление третичного во-
лосяного покрова и другие особенности, в том числе и наличие Rh-отри-
цательных индивидов, свидетельствуют о былом смешении.
Теперь предположим, что мы не обладаем антропологическими дан-
ными о предыстории современного населения южносибирского антропо-
логического типа. В этом случае возникли бы альтернативные смешению
гипотезы. Первая заключалась бы в том, что европеоидные черты в дан-
ном населении возникли за счет распространения нескольких мутаций,
произошедших внутри этой группы без европеоидного влияния. Вторая
гипотеза обосновывала своеобразие морфологии сохранением в данной
группе популяций древних, «недифференцированных» особенностей пе-
риода формирования современных расовых комплексов. Сторонники тре-
тьей гипотезы утверждали бы, что «псевдоевропеоидные» особенности
возникли благодаря отбору и адаптации в условиях приледниковой зоны,
в верхнем палеолите. Подобные или близкие гипотезы существуют по
поводу значительного числа популяций и антропологических типов. Можно
назвать саамов Кольского полуострова, группы населения, принадлежа-
щие уральскому антропологическому типу, многие группы Восточной Аф-
рики, Южной Индии и др.
Слабость мутационной гипотезы в том, что мутации случайны, веро-
ятность их закрепления в популяции мала. Для селективно нейтральной
мутации среднее время фиксации в популяции (т. е. достижения частоты
100%) равно 4N
e
поколений. Для принятых в предыдущей главе числен-
ностей популяции в 400-600 человек N
e
будет равно примерно 130—200
человек, что соответствует примерно 500—800 поколениям, или периоду
10 000—16 000 лет. Правда, для достижения средних частот нужно меньше
времени, но здесь вступает другой фактор — малая вероятность закреп-
ления именно «псевдоевропеоидной» комбинации генов (везде речь идет о
популяционных характеристиках). Процесс может быть ускорен, если воз-
никшие мутации имеют селективное преимущество, но если преимуще-
ство незначительное, то это слабо влияет на скорость процесса замены в
локусе старого аллеля на мутантный. Если это преимущество велико (что
считается крайне маловероятным событием), то мы возвращаемся к ди-
лемме Холдейна об одновременной адаптивной эволюции по нескольким
локусам. При отсутствии достаточных экспериментальных данных об адап-
тивности большинства признаков внешности мы остаемся перед дилем-
мой: либо рассматривать пару «смешанный — недифференцированный»,
либо сделать предположение о наследовании расовых признаков одним
342
супергеном. Интересно, что такую возможность считал наиболее вероят-
ной Я. Чекановский. Сам ход мышления можно считать полностью типо-
логическим! Как правило, этот путь отвергается на том основании, что
большинство расовых признаков связано между собой незначительными
внутригрупповыми корреляциями. Для классифицирования подбирают
признаки некоррелированные, но это еще не означает, что они не сцеп-
лены (см.: Ли, 1978). Рекомбинационный процесс может быть полностью
подавлен наличием инверсии, и целый участок хромосомы может себя
вести как неделимая единица.
Обычно рассматривается первая дуаль — «смешанный — недифферен-
цированный». Выбор делается на основании географического положения
группы, внутригрупповых корреляций, исторических и археологических
источников, лингвистической принадлежности и т. д. Нахождение группы
в зоне контакта предполагаемых исходных для нее типов и обычное на-
правление корреляционных связей между признаками трактуются в пользу
смешанного происхождения. Обратное положение рассматривается как
возможность сохранения древних особенностей.
При отсутствии достаточных палеоантропологических сведений на
выбор объяснения оказывают влияние взгляды исследователя на общий
ход дифференциации современного человека (моно- и полицентризм,
принятие даты образования основных расовых комплексов, признание
адаптивности расовых признаков или их селективной нейтральности) и
на частоты признаков, не входящих в классический набор, но имеющих
ясные географические градиенты в своем распределении. К ним относят-
ся некоторые группы крови (в примере с южносибирской расой это была
система Rhesus) или другие признаки внутренней среды (системы HLA,
Ар, Gm и т. д. ), дерматоглифика и др. Но по мере увеличения числа при-
знаков мы незаметным образом переходим от решения одной пробле-
мы — происхождения антропологического типа, к другой — происхожде-
нию конкретной популяции или системы популяций.
На настоящий момент в мире насчитываются многие сотни популя-
ций смешанного происхождения между значительно различающимися
антропологическими типами. Популяции, возникшие при смешении близ-
ких антропологических вариантов, мы рассматривать не будем, так как
громадное большинство ныне живущих популяций имеют смешанное
происхождение, и в этом смысле изучение любой популяции будет изу-
чением смешанной группы. Антропологов в первую очередь интересовали
популяции, образовавшиеся при смешении европеоидов, негроидов и
монголоидов или некоторых других ясно различимых морфотипов (авст-
ралийские аборигены, пигмеи и т. д. ).
С некоторой условностью эти работы можно разделить на изучение
гибридов первого поколения и последующих. С генетической точки зре-
ния фенотипические особенности гибридов первого поколения обуслов-
лены внутрилокусным взаимодействием. В последующих поколениях все
большее значение приобретает межлокусное взаимодействие.
Первое, что выяснилось в результате этих работ: плодовитость в браках
даже отдаленных друг от друга популяций не снижается. Одинаково пло-
довиты браки темнопигментированных экваториальных индивидов с
343
эскимосами и североевропейцами, французов и вьетнамцев, голландцев
и веддов, англичан и дравидийцев, русских и ительменов, немцев и буш-
менов, пигмеев и бантоидов и т. д. Есть большие группы населения, кото-
рые представляют собой трехрасовых гибридов. Их особенно много в Цент-
ральной и Южной Америке. Нормальная способность к воспроизведению
потомства у людей в межрасовых браках является основным аргументом
видового единства современного человечества. Потомство от таких браков
не имеет каких-либо дисгармонических физических черт. В двух широко-
масштабных исследованиях на Гавайях и в Бразилии было показано, что
некоторое повышение гетерозиготности в смешанных группах не оказы-
вает никакого влияния на соотношение полов, вес новорожденных, час-
тоту рождаемости монозиготных близнецов, мертворождаемость, детс-
кую смертность и частоту различных врожденных генетических аномалий.
Результаты этих и других работ косвенно показали, что частота многих
аномалий скорее всего является мутационным, а не сегрегационным гру-
зом. В связи с этими результатами неудивительно, что примерно каждый
десятый человек современности несет в своем генотипе гены из генофон-
дов по крайней мере двух больших рас.
Значительная часть данных по типу наследования основных расовых
признаков была получена при изучении смешанных групп (гл. 11). Инди-
видуальная комбинаторика признаков в смешанных группах ясно показа-
ла их независимость при расщеплении. У гибридов наблюдаются самые
«причудливые» с точки зрения обыденных стереотипов сочетания при-
знаков больших рас. Этот результат серьезно ставит под сомнение пред-
положения Я. Чекановского и всю типологическую концепцию. Здесь не-
обходимо точно оговориться, что мы не отрицаем возможности возник-
новения супергена по тому или иному комплексу признаков, применяемых
в систематике, но это событие, видимо, не частое.
Работы по смешанным группам населения не подтвердили еще двух
широко распространенных стереотипных мнений об обязательной про-
межуточности смешанных групп и о гетерозисных явлениях (так называ-
емая «гибридная сила»). Результат смешения в значительной степени за-
висит от конкретной истории рассматриваемой группы, структуры брач-
ных связей, особенностей рекомбинационного процесса в каждом
конкретном случае и т. д. Это не означает, что процесс смешения хаоти-
чен по своим результатам. Очень часто многие признаки в смешанных
группах занимают промежуточное положение, но в комплексах призна-
ков с независимым наследованием среднее положение нескольких при-
знаков не ведет с обязательностью к такому же положению очередного
изучаемого признака.
Интересны результаты изучения динамики развития детей в смешан-
ных популяциях в сравнении с контрольными. Общий ход развития не
имеет отличий от несмешанных групп. Если взрослые данной популяции
занимают промежуточное положение по какому-либо признаку то, как
правило, и дети этой популяции также занимают положение между деть-
ми исходных групп. Но бывает и так, что в ранних возрастах смешанная
группа ближе к одной из исходных, а в старших — к другой (рис. 128,
129). Сложность этих исследований заключается в разном социальном
положении сравниваемых групп. Полной ясности тут нет.
344
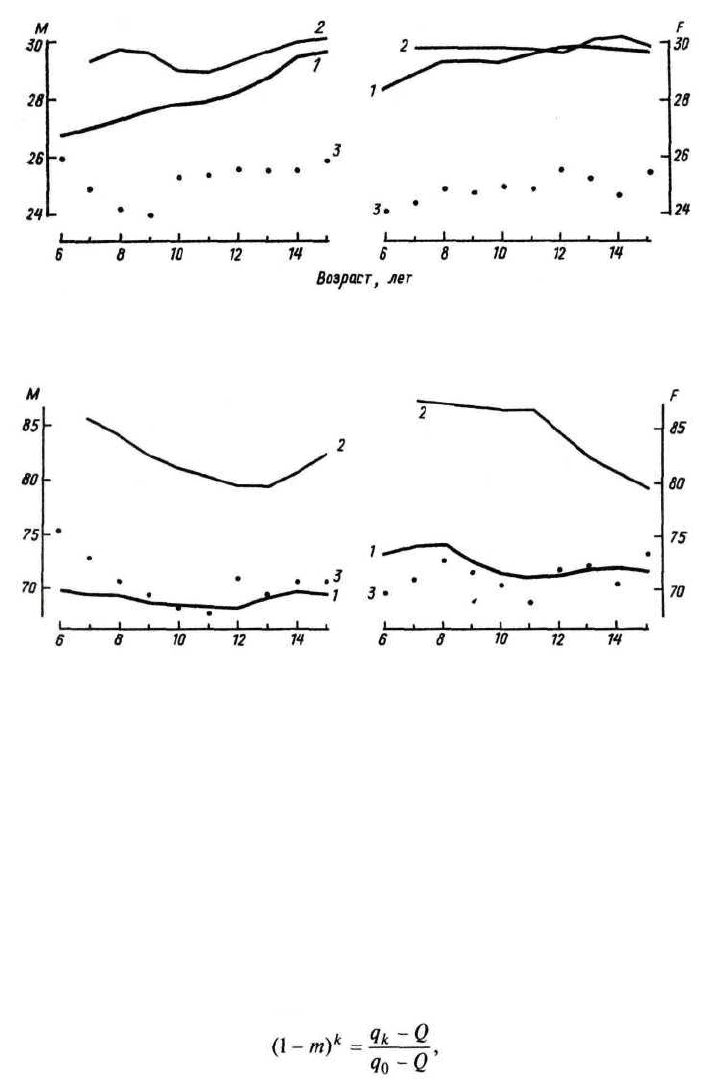
Рис. 128. Возрастные изменения отношения ширины носа к скуловой ширине у метисов
негров с японцами (1), американских негров (2) и японцев (3). Слева — мужчины, спра-
ва — женщины (Hoshi, 1973)
Нельзя не упомянуть еще об одном типе исследований. Это так назы-
ваемые расчеты доли примеси (или участия) исходных групп в смешан-
ной. Расчеты эти, как правило, делаются по дискретным признакам с
точно известной наследственностью. Обычно это различные группы кро-
ви, белки и ферменты, а также системы HLA и Gm. Есть система подобно-
го расчета и для цвета кожи. В основе данных расчетов лежит предположе-
ние, что частота гена в смешанной группе пропорциональна вкладу ис-
ходных групп. Для того чтобы избежать ошибок выборки, выбирают локусы
с сильно различающимися частотами аллелей в исходных группах и ре-
зультаты по разным локусам усредняют.
Основная формула следующая:
где q
0
и Q — частоты генов в исходных популяциях, q
k
— частота гена в
смешанной популяции, т — доля генов в смешанной популяции из по-
пуляции с частотой q
0
, k — число поколений.
345
Рис. 129. Возрастные изменения носового указателя в тех же группах, что и на рис. 128
(Hoshi, 1973)
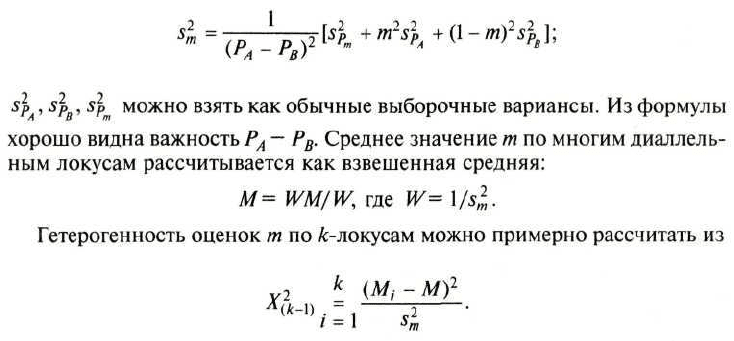
Существуют и более сложные методы расчета смешения и генного
потока. Во-первых, это оценки, разработанные для многих локусов и бо-
лее чем для двух исходных популяций, с получением обобщенного значе-
ния показателя т. Это метод наименьших квадратов (Roberts, Hiorns, 1965)
и метод максимального правдоподобия (Krieger et al., 1963). Различные
методы сравнивались между собой Р. Элстоном (Elston, 1971) и некото-
рые были им модифицированы. Как правило, различия в оценках невели-
ки. Во всяком случае, различия в обобщенных показателях по нескольким
локусам между разными методами значительно меньше, чем различия в
показателях т по отдельным локусам. В методическом отношении важной
была работа Т. Рида (Reed, 1969, 1973), который показал, что для боль-
шей достоверности оценок т необходимо подбирать локусы с максимально
различающимися частотами аллелей между родительскими популяциями.
Рид рекомендовал проверку однородности оценок т по отдельным локу-
сам, а также отбор локусов с наименьшей стандартной ошибкой. Он об-
ращает внимание и на то, что среднее значение т (т) желательно рас-
считывать через взвешивание (умножение) т отдельных локусов на их
пропорцию в общей информации и только после этого их суммировать
(так как информация есть сумма всех таких обратных значений, получен-
ных от каждой оценки генного потока). Формула вариансы следующая:
Так как т можно рассматривать как поток генов в поколении, то, если
знать дату начала смешения, можно рассчитать среднюю скорость в поко-
лении. Расчеты этого показателя для смешанных групп США, Бразилии,
Мексики, Камчатки дают очень близкие цифры: от 2 до 5%.
Скорость потока генов между соседними популяциями в группах охот-
ников-собирателей, как мы отмечали в предыдущей главе, также близка
к 5%. Видимо, эта величина характерна для человека современного вида
при обычных обстоятельствах обмена генетическим фондом.
При рассмотрении антропологических проблем удобнее различать си-
туацию, когда в результате смешения двух групп возникает третья — сме-
шанная, и ситуацию, когда одностороннее или двустороннее проникно-
вение генов (а точнее хромосом) создает промежуточные наборы геноти-
пов, но собственно самостоятельного смешанного генофонда не возникает.
346
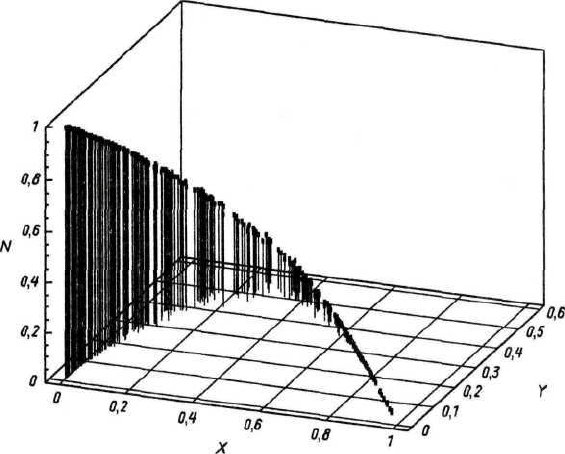
Термин «генный поток» лучше применять во втором случае (рис. 130).
Это, казалось бы, чисто количественное различие, тем не менее, имеет
важные последствия. При низкой интенсивности генного обмена (1—2%)
возрастает вероятность случайной утери привнесенного нового аллеля в
последовательном ряду поколений. Мы уже писали о некоторой аналогии
генного потока и мутационного процесса. То же самое можно сказать и о
передаче определенного гена по географической цепочке популяций. Ре-
комбинационный процесс может привести к ситуации, когда какой-то
определенный ген может проникнуть по цепочке популяций очень дале-
ко от своей исходной популяции, но в комбинации с совершенно други-
ми аллелями из других локусов данной хромосомы. В этом случае такой
«одинокий путешественник» может быть принят за параллельную мута-
цию в данном локусе. Для выбора между гипотезами необходимо провес-
ти поиск других признаков исходной популяции — донора (или проме-
жуточных популяций), так как вероятность мутационного происхожде-
ния нескольких сходных аллелей в разных популяциях будет достаточно
мала. Безусловно, что в пользу миграционного происхождения будет го-
ворить и наличие клинальной изменчивости.
Рис. 130. Генный поток в Северной Евразии.
Представлены результаты дискриминантного анализа примерно 400 черепов V—II тыс. лет
до н. э. по признакам горизонтальной уплощенности лица. Все множество было предвари-
тельно разделено на три географические зоны: 1 — Восточная Европа и Средняя Азия,
2 — Урал, Западная Сибирь и Алтае-Саянское нагорье, 3 — Байкальский регион. По
результатам дискриминантного анализа каждый череп получил вероятностные оценки
своей принадлежности к одной из зон (так называемые апостериорные вероятности). Пра-
вый нижний угол — максимальные вероятности — принадлежат к 1-й зоне, левый даль-
ний нижний угол — ко 2-й зоне, левый ближний верхний угол — к 3-й зоне. Хорошо
видно, что для территории Северной Евразии уже в древности был характерен достаточно
плавный и постепенный географический градиент (клина) по изменению признаков упло-
щенности лица в направлении восток-запад
347
Вышеизложенное приводит к выводу, что должны существовать некие
пороговые значения коэффициента миграции, ниже которых последствия
генного потока не фиксируются. Судя по приведенным выше экспери-
ментальным значениям т, этот порог, скорее всего, должен иметь значе-
ния ниже 1%.
§ 4. ИЗОЛЯЦИЯ И ПОЛИТИПИЯ
Неоднократно в истории человечества популяции и группы популя-
ций оказывались в полной изоляции от остального мира. Мы рассмот-
рим только биологический аспект изоляции — отсутствие генного по-
тока и миграций извне. С этой точки зрения безразлична причина изоля-
ции. Горный хребет и религиозная нетерпимость могут быть одинаково
эффективны. Например, парсы Бомбея (по происхождению иранцы-
персы), исповедующие зоорастризм, не менее строго изолированы в
течение многих сотен лет от своих соседей — индуистов и мусульман,
чем коренное население Австралии в течение тысячелетий от остально-
го человечества.
Изоляция часто рассматривается как важный, а иногда и единствен-
ный фактор политипии популяций. Интенсивность воздействия изоляции
на межгрупповую изменчивость прямо пропорциональна времени и об-
ратно пропорциональна численности популяции. Первое обусловлено
постоянством частоты мутаций, их случайностью и малой вероятностью
закрепления в популяции мутантной формы аллеля, если его не поддер-
живает отбор. В силу этого изоляты большой численности дивергируют
медленно. В малых изолятах генетический дрейф может способствовать
быстрому изменению генофонда за счет случайного закрепления одних
аллелей и потери других.
Одним из последствий малочисленности популяций является инбри-
динг, т. е. увеличение вероятности браков между близкими родственника-
ми. Обычно он приводит к росту гомозиготности. Так как многие наслед-
ственные дефекты рецессивны, то увеличивается вероятность фенотипи-
ческих отклонений от нормы. Все это может приводить к увеличению
числа людей с контрастными фенотипами и к уменьшению генетическо-
го разнообразия (за счет потери аллелей), снижению гетерозиготности и
общего фенотипического разнообразия. В известной степени изоляцию
можно уподобить центрифугированию, в которой скорость фракциони-
рования пропорциональна инбридингу.
Следует подчеркнуть, что в отсутствие «вредных» аллелей сам факт
увеличения гомозиготности, видимо, не наносит заметного ущерба по-
пуляции. Существует также мнение, что при некоторых обстоятельствах
(например, при пониженной жизнеспособности гетерозигот и отсутствии
воспроизводства у одной из гомозигот) инбридинг будет снижать частоту
неблагоприятного аллеля.
Ранее мы отмечали, что Homo sapiens в течение длительного периода
состоял преимущественно из небольших по численности популяций. Не-
сомненно, некоторые из них могли выпадать из системы генного обмена,
т. е. становились изолятами. Время изоляции могло быть большим. Например,
348
для населения Австралии и обеих Америк практическое отсутствие ген-
ного обмена с Евразией длилось тысячелетиями. В таком же положении
оказывались и некоторые окраинные группы Старого Света. Климатогео-
графические предпосылки для этого были.
Косвенным подтверждением роли изоляции и сопряженных с нею
случайных процессов в создании политипии является значительное сход-
ство филогенетических «деревьев» традиционных классификаций с со-
временными нумерическими классификациями, в большинстве которых
делается предположение о ведущей роли генетического дрейфа.
Есть еще одна особенность изоляции как фактора эволюции. В изолятах
могут длительное время сохраняться реликтовые формы. Правда, дли-
тельное существование малочисленного изолята — событие маловероят-
ное. Случайная гибель нескольких особей может фатально нарушить де-
мографическую структуру, и популяция вымрет. Можно полагать, что это
неоднократно происходило в истории человечества. К такому же резуль-
тату приводит уменьшение генетической изменчивости, приводящее к
сужению нормы реакции, а соответственно, и адаптационных способнос-
тей индивидов и популяций.
Глава 15
ЭТНИЧЕСКАЯ АНТРОПОЛОГИЯ
Под этнической антропологией обычно понимается описание
антропологических особенностей этносов. Во многих случаях антрополо-
гические данные содержат важную (а иногда и решающую) информацию
о путях формирования того или иного этноса— об этногенезе. В данной
области знаний о человеке антропология выступает совместно с. истори-
ей, этнографией и лингвистикой. Как уже говорилось выше, этнос опре-
деляется самосознанием, языком, самоназванием и некоторыми поведен-
ческими характеристиками. Ввиду этого объект исследования для антро-
полога определяют историки, этнографы и лингвисты.
Широко распространено мнение о существовании причинной связи
особенностей внешности с принадлежностью к тому или иному народу,
т. е. каждый народ имеет свой особый антропологический тип как неотъем-
лемую часть своей характеристики. Это мнение ошибочно вот по каким
общим соображениям и фактам.
Даже самые подробные антропологические классификации населения
Земли не превышают 200 морфологических вариантов. Количество же
этносов значительно больше: около 2000. Отсюда вывод: многие этносы
имеют очень сходный антропологический состав.
Если отвлечься от типологической системы описания разнообразия
человечества, то клины постепенных изменений частот самых разнооб-
разных признаков будут пересекать этнические границы, практически не
«замечая» их.
С другой стороны, большинство современных этносов, как правило,
политипично по антропологическому составу. Более того, в результате
постоянно идущих процессов смешения физический тип многих этносов
с трудом поддается описанию в рамках действующих типологических схем.
Правомерно интересоваться принадлежностью индивида к тому или
иному антропологическому типу (индивидуальная типология) либо
антропологической характеристикой того или иного этноса. Индиви-
дуальная типология на уровне антропологических типов, которые по
своей различимости соответствуют пандиакритическому и макродиак-
ритическому уровню, может быть вполне успешной. На более низких
уровнях различимости индивидуальная типология ненадежна и во мно-
гом связана с индивидуальным искусством классификатора. Антропо-
логическую характеристику этноса в силу ненадежности индивидуаль-
ной типологии лучше представить через средние величины измери-
тельных признаков и частоты качественных признаков или другие
статистические параметры, характеризующие изменчивость. Эти пара-
метры более надежны и допускают возможность сравнения антрополо-
гических характеристик этносов с помощью стандартных математи-
ческих методов.
350