Fishwick P.A. (editor) Handbook of Dynamic System Modeling
Подождите немного. Документ загружается.


28-12 Handbook of Dynamic System Modeling
TABLE 28.1
Continued
No. Rate (or Auxiliary) Equation
Units
Meaning
Used in Eq. No.
(017) J0FCS
=
K0FCS
J/y
Nonmarsh food assimilation by fishes, crabs, and shrimps
(016)
(018) J0LM
=1.42009e+017
J/y
Light influx
(027) and (031)
(019) JFCS0
=
KFCS0
∗
QFCS
J/y
Metabolic heat losses by fishes, crabs, and shrimps
(016)
(020) Jrain
=Rain
kg-H
2
O/y Rain on marsh
(002)
(021) JW2M2
=Tide
∗
KW2M2
∗
(WQ2
−KW2M2n
∗
kg-H
2
O/y Water exchange with estuary
(002)
MQ2)
(022) JW3M3
=KW3M3
∗
(WQ3 −
KW3M3n
∗
MQ3) kg-salt/y Salt exchange with estuary
(003)
(023) JW4M4
=KW4M4
∗
(WQ4 −
KW4M4n
∗
MQ4) kg-N/y
Inorganic nutrient exchange with estuary
(004)
(024) JW5M1
=KW5M1
∗
(WQ5 −
KW5M1n
∗
MQ1) J/y
Organic detritus exchange with estuary
(001)
(025) JW6M5
=KW6M5
∗
(WQ6 −
KW6M5n
∗
MQ5) kg-sediment/y Sediment exchange with estuary
(005)
(026) Jyield
=Yield
J/y
Catch of fishes, crabs, and shrimps in estuary and creeks
(016)
(027) MA07a
=J0LM /(1
+(MK07
∗
MA07b
∗
MQ7)+
J/y
Light influx available to low marsh plants
(029) and (033)
(MK20E
∗
MK20
∗
MQ2))
(028) MA07b
=(MQ2
∗
MQ4
∗
MQ6)/MQ3
Complex Environmental conditions for low marsh plant
photosynthesis
(027), (031), and (033)
(029) MA09a
=MA07a /(1
+(MK09
∗
MA09b
∗
MQ9)) J/y
Light influx available to benthic microalgae
(034)
(030) MA09b
=(MQ2
∗
MQ4
∗
MQ6)/MQ3
Complex Environmental conditions for benthic microalgal
photosynthesis (034) and (029)
(031) MA20
=J0LM /(1
+(MK07
∗
MA07b
∗
MQ7)
+
J/y
Light influx available to evaporate water from marsh
(040)
(MK20E
∗
MK20
∗
MQ2))
(032) MJ06
=Tide
J/y
Influx of tidal energy to marsh
(006)
(033) MJ07
=
MK07
∗
MA07b
∗
MQ7
∗
MA07a
J/y
Gross primary production by low marsh plants
(007), (041), (042), and (047)
(034) MJ09
=MK09
∗
MA09b
∗
MQ9
∗
MA09a
J/y
Gross primary production by benthic microalgae
(008) and (043)
(035) MJ15
=MK15
∗
MQ1
J/y
Burial of detritus in sediment (in energy units)
(001)
(036) MJ15D
=MK15D
∗
MQ1
kg-sediment/y Burial of detritus in sediment (in sediment units)
(005)
(037) MJ1a
=MK1a
∗
MQ1
∗
Mqa
J/y
Assimilation of detritus by microbes
(001) and (009)
(038) MJ1b
=MK1b
∗
MQ1
∗
Mqb
J/y
Assimilation of detritus by meiofauna
(001) and (010)
(039) MJ1c
=MK1c
∗
MQ1
∗
Mqc
J/y
Assimilation of detritus by macrofauna
(001) and (011)
(040) MJ20
=MK20
∗
MQ2
∗
MA20
kg-H
2
O/y Evaporation of water in marsh
(002)
(041) MJ207
=MK207
∗
MJ07
kg-H
2
O/y Transpiration of water by low marsh plants
(002)
(042) MJ47
=MK47
∗
MJ07
kg-N/y
Uptake of inorganic nutrients by low marsh plants
(004)
(043) MJ49
=MK49
∗
MJ09
kg-N/y
Uptake of inorganic nutrients by benthic microalgae
(004)
(044) MJ60
=MK60
∗
MQ6
J/y
Baseline dissipation of water circulation energy in marsh
(006)
(045) MJ607
=MK607
∗
MQ6
∗
MQ7
J/y
Dissipation of circulation energy by low marsh plants
(006)
(046) MJ70
=MK70
∗
MQ7
J/y
Baseline metabolic heat losses by low marsh plants
(007)
(047) MJ706
=MK706
∗
MJ07
J/y
Metabolic heat losses by low marsh plants due to production
processes (007)

Dynamic Simulation with Energy Systems Language 28-13
(048) MJ71
=MK71
∗
MQ7
J/y
Death of low marsh plant tissue and flow to detritus
(001) and (007)
(049) MJ7e
=MK7e
∗
MQ7
∗
MQe
J/y
Assimilation of low marsh plants by insects
(007) and (013)
(050) MJ90
=MK90
∗
MQ9
J/y
Metabolic heat losses by benthic microalgae
(008)
(051) MJ9b
=MK9b
∗
MQ9
∗
MQb
J/y
Assimilation of benthic microalgae by meiofauna
(008) and (010)
(052) MJ9c
=MK9c
∗
MQ9
∗
MQc
J/y
Assimilation of benthic microalgae by macrofauna
(008) and (011)
(053) MJ9e
=MK9e
∗
MQ9
∗
MQe
J/y
Assimilation of benthic microalgae by macrofauna
(008) and (013)
(054) MJa0
=MKa0
∗
MQa
J/y
Metabolic heat losses by detritus microbes
(009) and (055)
(055) MJa4
=MKa4
∗
MJa0
kg-N/y
Nutrient regeneration by detritus microbes
(004)
(056) MJab
=MKab
∗
MQa
∗
MQb
J/y
Assimilation of microbes by meiofauna
(009) and (010)
(057) MJac
=MKac
∗
MQa
∗
MQc
J/y
Assimilation of microbes by macrofauna
(009) and (011)
(058) MJb0
=MKb0
∗
MQb
J/y
Metabolic heat losses by meiofauna
(010) and (059)
(059) MJb4
=MKb4
∗
MJb0
kg-N/y
Nutrient regeneration by meiofauna
(004)
(060) MJbc
=MKbc
∗
MQb
∗
MQc
J/y
Assimilation of meiofauna by macrofauna
(010) and (011)
(061) MJbFCS
=MKbFCS
∗
MQb
∗
QFCS
J/y
Assimilation of meiofauna by fishes, crabs, and shrimps
(010) and (016)
(062) MJc0
=MKc0
∗
MQc
J/y
Metabolic heat losses by macrofauna
(011) and (063)
(063) MJc4
=MKc4
∗
MJc0
kg-N/y
Nutrient regeneration by macrofauna
(004)
(064) MJcd
=MKcd
∗
MQc
∗
MQd
J/y
Assimilation of macrofauna by raccoons and rats
(011) and (012)
(065) MJcFCS
=MKcFCS
∗
MQc
∗
QFCS
J/y
Assimilation of macrofauna by fishes, crabs, and shrimps
(011) and (016)
(066) MJcg
=MKcg
∗
MQc
∗
MQg
J/y
Assimilation of macrofauna by birds
(011) and (015)
(067) MJd0
=MKd0
∗
MQd
J/y
Metabolic heat losses by raccoons and rats
(012)
(068) MJe0
=MKe0
∗
MQe
J/y
Metabolic heat losses by insects
(013)
(069) MJef
=MKef
∗
MQe
∗
MQf
J/y
Assimilation of insects by spiders
(013) and (014)
(070) MJeg
=MKeg
∗
MQe
∗
MQg
J/y
Assimilation of insects by birds
(013) and (015)
(071) MJf0
=MKf0
∗
MQf
J/y
Metabolic heat losses by spiders
(014)
(072) MJfg
=MKfg
∗
MQf
∗
MQg
J/y
Assimilation of spiders by birds
(014) and (015)
(073) MJg0
=MKg0
∗
MQg
J/y
Metabolic heat losses by birds
(015)
(074) MJgd
=MKgd
∗
MQd
∗
MQg
J/y
Assimilation of birds eggs by raccoons and rats
(012) and (015)
No.
Inputs
Units
Meaning
Used in Eq. No.
(075)
Rain
=4.5e+010
kg-H
2
O/y
Annual rainfall on marsh
(020)
(076)
Tide =
1.03752e+014
J/y
Tidal energy influx to marsh
(021) and (032)
(077)
WQ2
=1e+011
kg-H
2
O
Water in estuary and tidal creeks
(021)
(078)
WQ3
=2.5641e+009
kg-salt
Salt in estuary and tidal creeks
(022)
(Continued)

28-14 Handbook of Dynamic System Modeling
TABLE 28.1
Continued
(079)
WQ4
=5600
kg-N
Nutrients in estuary and tidal creeks
(023)
(080)
WQ5
=5.1e+013
J
Organic detritus in estuary and tidal creeks
(024)
(081)
WQ6
=7e+006
kg-sediment
Sediment in estuary and tidal creeks
(025)
No. Model constants
Units
Meaning
Used in Eq. No.
(082) K0FCS
=2.898e+012
J/y
Nonmarsh food assimilation by fishes, crabs, and shrimps
(017)
(083) KFCS0
=0.38375
1/y
Fraction of biomass energy used per year in metabolic proc
esses by fishes, crabs, and shrimps (019)
(084) KW2M2
=5.86735e−014 1/y
Rate constant for water exchange with estuary
(021)
(085) KW2M2n
=18934.6
dmls
Equilibration constant for water exchange with estuary
(021)
(086) KW3M3
=3.04375
1/y
Rate constant for salt exchange with estuary
(022)
(087) KW3M3n
=10007.5
dmls
Equilibration constant for salt exchange with estuary
(022)
(088) KW4M4
=8.11667
1/y
Rate constant for inorganic nutrient exchange with estuar
y
(023)
(089) KW4M4n
=−1.09205 dmls
Equilibration constant for inorganic nutrient exchange with
estuary
(023)
(090) KW5M1
=1
1/y
Rate constant for organic detritus exchange with estuary
(024)
(091) KW5M1n
=0.0555556 dmls
Equilibration constant for organic detritus exchange with
estuary
(024)
(092) KW6M5
=1.33333
1/y
Rate constant for sediment exchange with estuary
(025)
(093) KW6M5n
=0.000492821 dmls
Equilibration constant for sediment exchange with estuary
(025)
(094) MK07
=1.1316e−033
complex Rate constant for primary production by low
marsh plants
(027), (031), and (033)
(095) MK09
=1.03014e−031 complex Rate constant for pr
imary production by benthic algae
(029) and (034)
(096) MK15
=0.0206749
1/y
Rate constant for detritus burial in sediment (detritus outflo
w)
(035)
(097) MK15D
=1.21617e−009 1/y
Rate constant for detritus burial in sediment (sediment inflo
w)
(036)
(098) MK1a
=2.37925e−012 1/(J-y) Rate constant for assimilation
of detritus by microbes
(037)
(099) MK1b
=2.18782e−014 1/(J-y) Rate constant for assimilation
of detritus by meiofauna
(038)
(100) MK1c
=5.46954e−015 1/(J-y) Rate constant for assimilation
of detritus by macrofauna
(039)
(101) MK20
=9.51857e−014 1/J
Fraction of water evaporated per unit of available light energ
y
(027), (031), and (040)
(102) MK207
=1.26752e−005 kg-H
2
O/J Water transpired per unit of energy assimilated
by low marsh plants
(041)
(103) MK20E
=
1.36889e+007 J/kg-H
2
O Energy required to evaporate water
(027) and (031)
(104) MK47
=2.25e−008
kg-N/J Inorganic nutrients assimilated per
unit energy assimilated by low marsh plants
(042)
(105) MK49
=2.5e−008
kg-N/J Inorganic nutrients assimilated per
unit energy assimilated by benthic microalgae
(043)
(106) MK60
=635.217
1/y
Fraction of water circulation energy dissipated per unit time
(044)
(107) MK607
=
7.68842e−014 1/(J-y) Fraction of circulation
energy dissipated year per unit of low marsh plants
(045)
(108) MK70
=0.773471
1/y
Fraction of biomass energy used in metabolic processes by low
marsh plants
(046)
(109) MK706
=
0.005
dmls
Fraction of low marsh plant production lost as heat
(047)
(110) MK71
=0.689162
1/y
Fraction of low marsh plant biomass that dies per year
(048)
(111) MK7e
=3.03863e−014 1/(J-y) Fraction of
low marsh plants consumed per year per unit insect
(049)

Dynamic Simulation with Energy Systems Language 28-15
(112) MK90
=69.0035
1/y
Fraction of biomass energy used per y
ear in metabolic processes by benthic microalgae
(050)
(113) MK9b
=1.36912e−011 1/(J-y) Fraction of benthic
microalgae consumed per year per unit meiofauna
(051)
(114) MK9c
=1.09529e−011 1/(J-y) Fraction of benthic
microalgae consumed per year per unit microfauna
(052)
(115) MK9e
=2.73823e−012 1/(J-y) Fraction of benthic
microalgae consumed per year per unit insect
(053)
(116) MKa0
=1747.32
1/y
Fraction of biomass energy used per year in metabolic proc
esses by detritus microbes
(054)
(117) MKa4
=5e−008
kg-N/J Regenerated nutrients per unit energy metaboliz
ed by detritus microbes
(055)
(118) MKab
=1.04007e−010 1/(J-y) Fraction of microbes
consumed per year per unit meiofauna
(056)
(119) MKac
=6.93382e−011 1/(J-y) Fraction of detritus
microbes consumed per year per unit macrofauna
(057)
(120) MKb0
=72.7162
1/y
Fraction of biomass energy used per year in metabolic proc
esses by meiofauna
(058)
(121) MKb4
=6e−008
kg-N/J Regenerated nutrients per unit energy metaboliz
ed by meiofauna
(059)
(122) MKbc
=1.60309e−012 1/(J-y) Fraction of meiofauna
consumed per year per unit macrofauna
(060)
(123) MKbFCS
=8.36395e−014 1/(J-y) Fraction of meiofauna
consumed per year per unit fishes, crabs, and shrimps
(061)
(124) MKc0
=48.7219
1/y
Fraction of biomass energy used per year in metabolic proc
esses by macrofauna
(062)
(125) MKc4
=6e−008
kg-N/J Regenerated nutrients per unit energy metaboliz
ed by macrofauna
(063)
(126) MKcd
=1.71895e−012 1/(J-y) Fraction of macrofauna
consumed per year per unit raccoons and rats
(064)
(127) MKcFCS
=4.48327e−014 1/(J-y) Fraction of macrofauna
consumed per year per unit fishes, crabs, and shrimps
(065)
(128) MKcg
=4.29646e−013 1/(J-y) Fraction of macrofauna
consumed per year per unit birds
(066)
(129) MKd0
=4.93012
1/y
Fraction of biomass energy used per year in metabolic proc
esses by raccoons and rats
(067)
(130) MKe0
=31.3155
1/y
Fraction of biomass energy used per year in metabolic proc
esses by insects
(068)
(131) MKef
=6.90377e−012 1/(J-y) Fraction of insects
consumed per year per unit spiders
(069)
(132) MKeg
=6.90377e−013 1/(J-y) Fraction of insects
consumed per year per unit birds
(070)
(133) MKf0
=15.6578
1/y
Fraction of biomass energy used per year in metabolic proc
esses by spiders
(071)
(134) MKfg
=6.90377e−
013 1/(J-y) Fraction of spiders consumed
per year per unit birds
(072)
(135) MKg0
=
2.69679
1/y
Fraction of biomass energy used per year in metabolic processes
by birds
(073)
(136) MKgd
=
2.37812e−013 1/(J-y) Fraction of birds eggs
consumed per year per unit raccoons and rats
(074)
(137) Yield
=0
J/y
Capture of fishes, crabs, and shrimps in estuary and creeks
(026)
No.
Simulation time parameters
b
Units
Meaning
(138)
FINAL TIME
=4
y
The final time for the simulation
(139)
INITIAL TIME
=
0
y
The initial time for the simulation
(140)
TIME STEP
=2e−007
y
The time step for the simulation
(141)
SAVEPER
=0.001
y
The frequency with which output is stored
a
INTEG(inflows
−outflows) refers to the integration of the net flow rate
to determine the storage value. The initial values given for each stor
age are the assumed steady-
state values used for
model calibration. These values yield a flat-line output. To obtain
dynamic output, halve the initial value for MQd.
b
Vensim simulation software was used with the fourth-order
Runge–Kutta routine se
lected. The model was run with identical results both
on a MacIntosh
PowerBook G4 (1 Ghz PowerPC
G4 (3.3) processor, OS: Apple Classic 9.2.2 under OS10.4.6) and
on an Abit KT7-RAID PC motherboard with an AMD Duron 700
processor (OS: Windows 2000 Pro
fessional).
28-16 Handbook of Dynamic System Modeling
28.3.2 Basic Equation Forms Indicated in Energy Systems
Language Diagrams
Each storage in an EnergySystems Language diagram has lines indicating one or more inflows and outflows.
The equation for each storage is simply the integral of its net flow rate over time, that is to say, the integral
of the sum of all inflows into it minus the sum of outflows from it from all time past to the present
moment. Because the concept of “all time past” is indefinite, an initial starting time is specified (Time
Zero), at which point each storage is represented by an initial value. Eq. (001)–(016) in Table 28.1 show
the rates contained in an integration function and the initial value supplied for each of the model storages.
The form of each rate equation, however, is tied to specific details in the diagram. Each rate arises from a
symbol combination. The translation rules can be illustrated best with simpler diagrams of processes that
repeat many times in Figure 28.3. By applying a few simple rules, the entire equation list can be constructed.
The composite symbols for primary producers (bullet shapes) and consumers (hexagons) have the
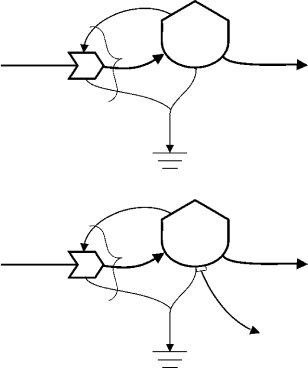
same standard symbol combination to represent a reproduction process called “autocatalytic growth.”
This standard structure is given in Figure 28.4. The only difference between primary producers and
consumers is the type of energy assimilated: usually sunlight for primary producers and some form of
organic matter for consumers.
In either case, the designated energy source R passes at rate J
1
through a multiplicative interaction to be
assimilated and stored in biomass Q
1
. As shown in Figure 28.4, energy from the recipient storage (Q
1
) feeds
back and intersects the top of the interaction symbol, and then dissipates as heat to the heat sink symbol.
TherateJ
1
is a net assimilation rate. Assimilation measurements for most living organisms are usually of
net assimilation. As the curly bracket in Figure 28.4 indicates the energy of Q
1
fed back and used specifically
in the production process is rarely separated explicitly from the amount assimilated. In ecological literature,
the rate J
1
is known simply as “assimilation” for consumers. For primary producers, it is the gross primary
production. The net production rate is J
1
−J
2
, which is net primary production in the case of plants and
net secondary production for consumers. Old age death, or other passive loss of biomass is indicated by
rate J
3
.
The equations for the relevant energy flows and their combined effect on the storage are indicated in
the top panel of Figure 28.4. Consider first the net assimilation rate J
1
, the form of which arises from the
multiplicative interaction symbol involved. According to the translation rules of Energy Systems Language,
allflows(J
i
) into and out of an interaction term have the same equation form. Equations differ only by
the numerical value of an associated constant (k
i
). In the case of multiplicative interaction, the resource
(R) and the biomass (Q
1
) multiply together to produce the indicated equation for net assimilation rate
J
1
=k
1
RQ
1
. In other words, the rate of production is proportional to the multiple of the amount of energy
availablefor transformation in resource R and the amount already available in the autocatalyticbiomass Q
1
.
The biomass acquires resources to form additional biomass. That is to say, in the presence of its resources,
growth is autocatalytic. The rate constant k
1
is analogous to the second-order rate constant in chemical
reaction kinetics. The dimensional units for k
1
are fractions of R assimilated per unit Q
1
per unit time.
The outflows J
2
and J
3
in Figure 28.4 do not pass through an interaction symbol as drawn. If they did,
the form of the equation would follow interaction rules like those for J
1
. When the rate of outflow is simply
a constant proportion of the donor storage, the outflow line does not pass through an explicit interaction
symbol downstream. Hence, the equations for J
2
and J
3
in Figure 28.4 are constant proportions of Q
1
.The
dimensional units of the rate constants involved are simply the fraction of Q
1
that is degraded per unit
time for k
2
, or in the case of k
3
, the fraction of upgraded material (Q
1
) that dies from old age, leaks out,
or is otherwise passively lost (per unit time).
In the complete marsh sector diagram (Figure28.3), all of the metabolic heat losses from living organisms
are constant proportions of the energy in biomass. These flows are identified in the equation list (Table 28.1)
as MJx0, where x is the subscript associated with the standing stock Q of a living component x. Donor
proportional outflows are used elsewhere in Figure 28.3 as well. The flow from plants to detritus (MJ
71
,
Eq. [048]) is proportional to plant biomass (MQ
7
) and the burial of detritus in sediments as peat (MJ
15
,
Eq. [035]) is proportional to the detritus accumulation (MQ
1
).
Dynamic Simulation with Energy Systems Language 28-17
R
(a)
(b)
x
J
2
J
3
J
1
k
1
R
.
Q
1
J
2
k
2
Q
1
J
3
k
3
Q
1
dQ
1
/dt J
1
J
2
J
3
Q
1
J
2N
k
2N
Q
1
J
1
R
x
J
2
J
3
Q
1
J
1
FIGURE 28.4 Basic symbolic representation and equations for the autocatalytic process usually used in the com-
posite symbols for primary producers and consumers: (a) without nutrient regeneration indicated; (b) with nutrient
regeneration indicated (J
2N
). The curly bracket pointing to J
1
indicates combined flows (see text). R is a variable that
represents the energy resource to be transformed, J’s are for the various flows, k’s are rate constants, Q
1
represents the
storage of biomass, and dQ
1
/dt is the rate of change of biomass with time.
Although not illustrated in Figure 28.4, some of the biomass of one living component may be captured
and consumed by predators or other consumers. The amount is called the yield of biomass, and it becomes
the resource for the autocatalytic growth of the recipient consumers. The equation for such an outflow
from Q
1
to a downstream consumer would have the same multiplicative form as used for J
1
. If, for example,
Q
1
was the food of consumer Q
2
, the flow between them—call it J
4
—would be J
4
=k
4
Q
1
Q
2
. The constant
k
4
would represent the fraction of Q
1
assimilated per unit time by each unit of consumer Q
2
.
A food Web can be represented by linking primary producer symbols to a network of consumer symbols.
Consumers may represent trophic levels, feeding guilds, or other functionally similar combinations of
species. Figure 28.3 includes a food Web based on marsh plant detritus, live marsh plants, and benthic
microalgae that after passing through a number of intermediate consumers, culminates in the birds,
raccoons,and rats of the salt marsh and the fishes, crabs, and shrimps of the tidal creeks and adjacent estuary
(Q
FCS
). Each equation for an energy transfer from resource to consumer uses the two-way multiplicative
interaction described above.
The composite symbol for consumers sometimes has a flow of regenerated nutrients indicated in
conjunction with the metabolic energy loss. One way to handle this is illustrated by the flow labeled J
2N
in the bottom panel of Figure 28.4. The flow line emanates from a small rectangle attached close to the
bottom of the storage tank. The small rectangle, called a “sensor,” denotes that no loss of the storage of
energy occurs, in this case because the flow is in units of something else besides the energy that is stored in
biomass (nutrients). Nutrient regeneration is a product of metabolism. Nutrients accompany the energy
losses denoted by the flow to the sink, but they must be tracked in a separate flow stream. Regenerated
nutrients flow from the consumer to a nutrient storage elsewhere. The lower panel of Figure 28.4 shows
the equation for nutrient regeneration (J
2N
). It has the same form as the metabolic energy loss equation
(J
2
), but the dimensional units for the constant (k
2N
) are nutrient units released per unit time per unit of
energy stored, rather than fraction of energy per time.
The marsh sector diagram (Figure 28.3) shows nutrient regeneration by microbes (MQ
a
), meiofauna
(MQ
b
), and macrofauna (MQ
c
). The nutrient regeneration flow lines arise from the vicinity of the
28-18 Handbook of Dynamic System Modeling
metabolic heat losses for these three consumers and cycle up and to the left terminating in the nutrients
storage (MQ
4
). The tiny sensor symbols are left out of the diagram in Figure 28.3, and in the equation list
(Table 28.1) an alternate equation form is used for MJ
a4
,MJ
b4
, and MJ
c4
(Eq. [055], Eq. [059], and Eq.
[063], respectively). In this form, nutrient regeneration is a constant proportion of the respective metabolic
heat losses rather than to the amount of energy stored in each consumer’s biomass. The constants therefore
define the amount of nutrients regenerated per unit of energy metabolized, which may be conceptually
more familiar to some. Either way is consistent with the diagram.
In either case, the flow of regenerated nutrients is not subtracted from the accumulations of energy
(MQ
a
,MQ
b
, and MQ
c
) because the associated energy subtraction takes place with the respective flows
to the heat sink (see Eqs. [009]–[011]). The nutrient flows are, however, added to the accumulation of
nutrients (Eq. [004]).
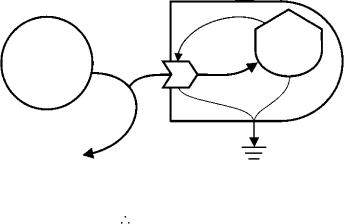
28.3.2.1 The Flow-Limited Source
A curved line entering the system boundary from a source and then passing back out, while being tapped
along the way inside the boundary, means that the equation for a “flow-limited source”is to be used. In the
coastal system and marsh sector diagrams (Figure 28.1 and Figure 28.3), sunlight is tapped by evaporation
of water and by the various primary producers in the system. Unused light is reflected by the system (the
system’s albedo), as represented by the line that curves back out of the system boundary.
Some resources, such as fossil fuels, are limited by the amount available in storage. For these resources,
use rate exceeds the regeneration rate. Once the storage is depleted, further use is limited by the rate of
regeneration. The resource then becomes flow-limited. Water flowing in a hillside stream is limited by the
flow from upstream. From its first diversion for irrigation or drinking water, a stream is recognized as
flow-limited. After diversion only the remaining flow is available for additional withdrawals. Likewise, the
flow of sunlight ultimately limits photosynthesis. Sunlight used in evaporating water is not available for
photosynthesis, and only the sunlight remaining in the shade and sunflecks beneath the tallest plants can
be used by the shorter ones.
When storage is the resource transformed for the production of an autocatalytic component, the
associated flow rate equationsarebased on the multiple of the donor and recipient storages. The production
rates of all consumers of the food Web illustrated in the marsh sector diagram (Figure 28.3) are of this type.
For primary producers, however, the main energy resource is flow-limited. Hence, the equation for gross
primary production is based on the remaining unused flow of the resource, rather than a donor storage.
The remaining flow is known generally as J
R
. A simple diagram of this process is shown in Figure 28.5.
In this case, the only tap is for gross primary production (J
1
), so the remaining light (J
R
) is simply the
solar input (J
0
) less that already incorporated into production. Substituting and rearranging this set of
implicit equations yields the explicit relationship suitable for models. The equation for J
R
is hyperbolic
and declines to zero as the storage grows. The first equation for J
1
is simply the multiple of J
R
and Q
1
.
The dimensional units of the constant k
1
are the inverse of the storage units. The constant represents the
fraction of the remaining light assimilated by each unit of biomass.
The equation derived for J
1
from the definitions of J
1
and J
R
(shown in Figure 28.5) is the Monod
function (or the Michaelis–Menten equation), often used in ecological models to represent flow saturation
at high levels of a resource and limitation at low levels. In the case illustrated, the maximum gross primary
production rate is equal to J
0
and the level of biomass energy (Q
1
) at which the rate is half the maximum
is the inverse of k
1
. The practical maximum is actually considerably less than J
0
for two reasons: (a) other
factors also limit photosynthesis; and (b) other processes compete for sunlight, such as water evaporation,
conversion to heat on dark surfaces, and reflection.
In the marsh sector diagram (Figure 28.3), the taps of sunlight for evaporation and for marsh plant
production originate from the same point, and the tap for benthic microalgae occurs afterwards. Usually,
tapping from the same point means the processes are all competing for the same remaining light, however,
this is not the case when the point refers to different areas. The vertically overlapping primary producer
symbols for low and high marsh plants indicate this distinction. The various grasses, rushes, and sedges that
comprise each of the two elevation regions exist literally side by side in the marsh, so they receive the same
Dynamic Simulation with Energy Systems Language 28-19
x
J
2
J
1
J
1
k
1
J
R
Q
1
J
R
J
0
J
1
Q
1
J
0
J
R
J
R
J
0
/(1 k
1
Q
1
)
J
1
J
0
Q
1
/[(1/k
1
) Q
1
)
]
Sun
Plants
FIGURE 28.5 Flow-limited source coupled with an autocatalytic primary production process. Equations for J
1
and
J
R
are given two ways.
amount of light from overhead. A complete model of the marsh sector would have two subsectors: one for
the low elevation marsh and the other for the high marsh. The equations of Table 28.1 show only the low
marsh subsector. The high marsh would be represented by an identical set of equations calibrated for high
marsh conditions. The total marsh sector would consist of the sum of each component in each subsector.
The independent process of water evaporation shown at the same tap point as plant production,
however, is a competing process, separately competing in both the low marsh and the higher elevation
marsh. Processes competing for the same flow-limited resource use the same calculation of remaining light
for its functions. Water evaporates through an interaction with sunlight, as illustrated in the diagram. The
equation for the evaporation rate (MJ
20
, Eq. [040]) is based on the multiple of the storage of water (MQ
2
),
and an auxiliary that provides the remaining light available (MA
20
, Eq. [031]). The right-hand side of the
corresponding auxiliary equation for plants is identical (MA
07a
, Eq. [027]). The form of each equation
for light remaining at the combined tap is similar to that given for a single tap in the simple diagram of
Figure 28.5, except that the denominator is based on the sum of the competing uses: evaporation of water
and production by low marsh plants. Note also the conversion of evaporation from water to energy using
the constant MK
20E
(specified in Eq. [103]) for dimensional consistency.
Benthic microalgae exist in water-saturated soil often covered by water, and they grow in the shade of the
emergent vascular plants of the marsh. Accordingly, the diagram shows the sunlight for benthic microalgae
drawn downstream of the simultaneous tap for the marsh plant production and water evaporation. The
source light available to benthic microalgae (analogous to J
0
in Figure 28.5) is the light remaining after
evaporation of water and assimilation by the plants that shade them. This was already determined by
MA
07a
in Eq. (027) (or equivalently by MA
20
in Eq. [031]). Otherwise the calculation of J
R
for benthic
microalgae (MA
09a
) follows the form given in Figure 28.5, as shown in Eq. (029).
28.3.2.2 Multiple Simultaneous Interactions in Energy Systems Language
According to Figure 28.1 five other factors influence the transformation of sunlight by plants. These are
illustrated more precisely in the marsh sector diagram (Figure 28.3). Water, nutrients, sediment, and
circulation energy (caused by winds and tide) all positively influence gross primary production. In Energy
Systems Language, these combine in a series of multiplicative interactions to determine the rate of gross
primary production. Salt negatively influences gross production, however, so it is represented by a divisor.
The resulting autocatalytic production equation for plants consists of a constant multiplied by the four
positive influence variables, divided by salt, and all multiplied as before by the remaining light and the
energy stored in plant biomass. A total of seven state variables are combined with one constant creating a
seventh-degree equation for gross photosynthesis.
The multiplication of all these values to achieve a given rate of gross photosynthesis means that the
single constant involved will have very complex dimensional units. Because production units and light
28-20 Handbook of Dynamic System Modeling
flux units are both in energy units per time, a production function based only on the multiple of remaining
light and biomass involves a constant with dimensions of inverse biomass units (as shown in Figure 28.5).
If, however, two additional variables (say A and B) multiply and one divides (C), then for dimensional
consistency, the implied units for the constant are C units per A unit per B unit per biomass unit. In the case
given by the marsh sector diagram in Figure 28.3, the constant must cancel units from a total of six variables.
An additional practical detail concerning the constant is the small numerical value that must be assigned
to it. The multiple of all the variables involved in a complex interaction produces a very large number that
is proportional to the much smaller number that represents the resulting production rate. A very small
constant does the conversion. In fact, it can be too small to represent on a computer!
The need for a tiny constant became a significant practical issue in the marsh sector simulation, so the
multiplier auxiliaries for the gross photosynthesis equations, MA
07b
and MA
09b
(Eq. [028] and Eq. [030]),
do not include the influence of sediment (MQ
5
). To include the sediment influence, the constant needed
was on the order of 10
−44
. The number was too small to be represented precisely enough to run a required
model check on the available computer. Sediment was potentially the least dynamic influence on gross
production. The timescale of sediment dynamics in the model was on the order of 10,000 y, whereas the
next longest time scale (that of salt) was around 3 y. Sediment was unlikely to influence model output
spanning less than a few decades. By leaving out the sediment influence on production, the order of
magnitude for the gross production constant was increased to 10
−33
, large enough to allow the simulation
check (the check involved reproducing the steady-state condition used to calibrate the model).
28.3.2.3 Feedback Effect of Production Processes on the Environmental
Variables Involved
Some of the environmental influences on production are significantly consumed in the process. Nutrients,
for example, are used at a rate proportional to gross production of low marsh plants (MJ
47
, Eq. [042]) and
benthic microalgae (MJ
49
, Eq. [043]). Likewise, water is transpired by low marsh plants (MJ
207
, Eq. [041]),
and circulation energy is dissipated by marsh plant biomass (MJ
607
, Eq. [045]). The accumulations of salt
and sediment, however, are not changed directly by gross primary production. This lack of effect is denoted
in Figure 28.3 by the use of the sensor symbol (small rectangle attached to the storage symbol where the
line of influence originates).
Because gross production by benthic microalgae is not explicitly drawn on the marsh sector diagram,
the influencing variables and any influences on them are unspecified. The same variables involved in
low marsh plant production are involved with benthic microalgae. An examination of the equation list
(Table 28.1) shows that with the exception of nutrient uptake (MJ
49
, Eq. [043]), gross primary production
by benthic microalgae does not consume any of the variables that influence it, at least for this model.
28.3.2.4 Two-way Interactions
In coastal systems, water circulating with the tides creates an exchange of materials between marshes
and adjacent estuarine waters. Five exchanges are illustrated at the top left of the marsh sector diagram
using two-way interaction symbols acted upon as a group by the tides. These are water, salt, nutrients,
detritus, and sediment. All of these exchanges are driven by tidal energy. A two-way interaction indicates
a balance of forces arising from storages of the same material on the two sides of the interaction. In the
case of Figure 28.3, the storages are in the marsh and in the adjacent waters. A tendency toward dynamic
equilibrium of materials is expected. The rate equation involves the difference between the two storages.
Its general form is
J = k
r
(Q
S
−k
e
Q
R
)
where Q
S
is the quantity stored in the designated source location, and Q
R
is that stored in the receiving
location. A positive difference in parentheses sends the flow toward the recipient, and a negative difference
establishes flow in the opposite direction. For the marsh sector model, a positive flow represents an import
into the marsh from the estuary. The constant k
e
is the dimensionless equilibrium ratio of Q
S
:Q
R
expected
once a dynamic equilibrium is established. The other constant k
r
is a rate constant: the fraction of the

Dynamic Simulation with Energy Systems Language 28-21
difference within the parentheses that can be closed per unit time. It is inversely proportional to the half-life
of the difference (half-life =ln 2/k
r
).
The above rate equation was applied to the two-way flow for water, salt, inorganic nutrients, organic
detritus, and sediment in Eqs. (021)–(025) in Table 28.1. Note that Eq. (021) for exchange of water (J
W2M2
)
includes the multiplication by tide. This is not the case for the other four variables. In those cases, the
effect of tide is implicit in the selection of a value for the rate constant. As long as tidal influx is constant,
this distinction is unimportant to the model output; however, before tidal energy influx can be tested as
a variable, all the two-way exchanges must be converted to the form of Eq. (021). The conversion can be
done simply by dividing each rate constant by the constant tidal influx used now (Eq. [086]). The values
in question are given in Eq. (086), Eq. (088), Eq. (090), and Eq. (092), respectively.
28.3.2.5 Passive Inputs and Environmental Conditions
A basic input from the environment passively received by storage within the system is perhaps the simplest
concept used to create a rate equation. Unlike with flow- and storage-limited sources from outside the
system boundary, nothing within the system feeds back to limit the availability of such inputs. Examples
in the marsh sector model are the input of rain to marsh water (Rain, Eq. [075]) and the input of tidal
power to circulation (Tide, Eq. [076]).
When modeling a sector of a larger system, inputs from other sectors become basic environmental
conditions similar to passive inputs. For example, inputs to the marsh sector include the storages of water,
salt, nutrients, organic detritus, and sediment in the adjacent estuarine water (Eqs. [077]–[081]). These
storages would become dynamic model variables of the water sector in a complete model of the coastal
system diagrammed in Figure 28.1. Instead, they are set as passive inputs that do not change in response
to the flows in and out of the marsh.
A passive input may be an average represented as a constant, a test pattern (such as a sine wave), a statis-
tical distribution, or a complex time variable based on real data. In the early stages of model analysis, such
variables are often set to average conditions represented by constants so as to not confound dynamics gener-
ated by the model’s own feedback structure with those that are simply responding to variable driving forces.
Holding the environmental inputs constant allows the internally created dynamics to be revealed first.
28.3.2.6 An Unspecified Process (Rectangle)
The interaction of tide with the exchange of water circulation is represented by a small rectangle, indicating
a general process that must be made explicit in the documentation. In Figure 28.3, the rectangle merely
signifies that water circulation energy is affected by the tide and by other forces provided through the water
sector. Wind effects, for example, are shown in the coastal system diagram of Figure 28.1. The two kinds
of forces are additive, but the marsh sector model only uses tidal power to drive circulation.
28.4 Calibration of Model Constants
All of the types of rate equation indicated or implied in the marsh sector diagram and given in the equation
list have now been defined. The next step is to provide numerical values for all constants. In Energy
Systems Language, this is nearly always accomplished by back calculation from quantitative estimates of
all storages, rates, and system inputs. To do this, the rate equations are algebraically rearranged to solve for
the constants in terms of the rates, storages, and inputs involved. The process of back-calculating constants
in this manner is called model calibration.
To illustrate, consider the detailed diagram for a living composite symbol given in Figure 28.4(a). With
estimates of the amount storage (Q
1
), the amount of resources (R), and flow J
1
, an estimate for the constant
k
1
can be calculated. Likewise, with estimates of the storage Q
1
and each of the other flows (J
2
, and J
3
),
the constants k
2
and k
3
can be made.
Obtaining a complete set of applicable estimates for model calibration can be a time-consuming task;
however, it is easier than obtaining independent estimates for the constants themselves. Many of the