Драбкова В.Г., Прыткова М.Я., Якушко О.Ф. Восстановление экосистем малых озёр
Подождите немного. Документ загружается.


рования. Слабые системы реагируют на внешние перемены только
структурными перестройками. Озерные экосистемы во всей иерархии
своей организации также включают слабые подсистемы. К таким под-
системам следует отнести биоценозы. Чем выше уровень рассмотре-
ния экосистемы, тем сильнее системообразующие связи.
Прежде чем переходить к рассмотрению пределов устойчивости
экосистемы, следует остановиться на тех показателях, которые могут
характеризовать отклонения экосистемы от принятой нормы. А. П. JIe-
вич (1976) выделяет структурные изменения (выпадение или внедре-
ние групп особей); организационные изменения (изменения наличия
или интенсивности различных типов связей, например перераспре-
деление интенсивности трофических связей при изменении числен-
ности видов); функциональные изменения. Эти изменения могут су-
ществовать независимо друг от друга, но, как правило, они взаимо-
связаны. Структурные изменения затрагивают прежде всего системы
со слабыми взаимодействиями. В водной экосистеме это биологичес-
кое сообщество, популяция, виды. Организационные и функциональ-
ные изменения присущи всей экосистеме.
Когда мы говорим об устойчивости экосистемы, то имеем в виду
устойчивость к конкретным воздействиям, а не устойчивость вооб-
ще. В зависимости от характера воздействия одна и та же система
может быть устойчива, менее устойчива и неустойчива.
Остановимся на способах измерения устойчивости. Сразу надо
отметить, что не существует общепринятых методов таких измере-
ний. Проблема несколько упрощается, когда речь идет не обо всей
экосистеме, а лишь о ее живой компоненте. Обычно исходят из того,
что стабильность отдельных характеристик ведет соответственно
к устойчивости иерархически более организованной и сложной сис-
темы. Однако это не является бесспорным. .
В качестве меры устойчивости отдельных сообществ предлагают-
ся следующие показатели. Может быть использовано отношение ми-
нимальной за год биомассы сообществ к максимальной, которое
характеризует изменчивость биомассы в течение года (Алимов, 1989),
а также отношение дыхания сообщества к его биомассе и др. Около
30 лет назад Р. Макартур (Macarthur, 1955) и Р. Маргалеф (Margalef,
1958) предложили использовать индекс видового разнообразия как
показатель устойчивости D = -NI P
1
-InP
1
-, P
1
= JV
1
- /JV, N=Z N
1
, где N -
численность видов. Однако использование этого критерия устойчи-
вости не совсем оправдано. Дело в том, что максимально возможно-
му разнообразию соответствует структура сообщества, при которой
все виды встречаются с одинаковой частотой. Отсюда следует, что
наиболее устойчиво сообщество, в котором не существует иерархии
видов по численности, так как все виды встречаются с одинаковой
частотой. А это противоречит реально наблюдаемой картине. И толь-
ко на ранних стадиях сукцессии при движении экосистемы к зрелос-
ти действительно наблюдается увеличение разнообразия (Свирежев,
1983).
71

Чем устойчивее сообщество, тем выше соотношение стено- и эв-
рибионтных видов, т. е. выше доля в биоценозе стенобионтных (бо-
лее специализированных) видов.
Наиболее распространенный способ описания структуры систе-
мы - это способ, связанный с описанием структуры потоков энергии
и вещества через экосистему, так называемый трофический граф со-
общества. Ю. М. Свирежев (1983) описывает сложность, исходя из
взаимоотношений типа „хищник-жертва" как отношение числа ре-
ально существующих связей к числу возможных связей. Однако надо
принять во внимание, что усложнение системы не всегда приводит
к ее устойчивости и наоборот. Нередко в процессе эволюции экосис-
темы уменьшается длина трофической цепи, т. е. уменьшается ее
сложность.
Приведенные примеры оценки устойчивости системы касаются
лишь биологических сообществ. Существуют также некоторые по-
пытки рассматривать устойчивость экосистемы более широко. Так,
Б. Паттен получил на основании эмпирических расчетов числовое вы-
ражение для стабильности внешней среды и планктонных сообществ.
Он исследовал 11 переменных, относящихся к характеристике фито-
планктона и 11 - к состоянию внешней среды. Показатель стабиль-
ности всей системы определялся как средняя величина из стабиль-
ности всех изученных переменных. Значение этой величины выше
нуля выражает стабильность системы, ниже нуля - нестабильность
(цит. по: Федоров, Соколова, 1972). В. Д. Федоров и С. А. Соколова
предлагают при оценке стабильности экосистемы в целом не противо-
поставлять оценки стабильности косной и живой природы, а опери-
ровать с оценками логически соподчиненных характеристик (напри-
мер, свет- температура - хлорофилл- фотосинтез- кислород).
По мере развития озерной экосистемы от олиготрофии к эвтрофии
до какого-то периода идет ее усложнение, и с учетом всего сказанного
ранее можно говорить, что возрастает устойчивость этих экосистем.
Некоторые авторы указывают на слабую устойчивость глубоких стра-
тифицированных озер с признаками олиготрофии и мелководных вы-
сокоэвтрофных. О. Ф. Якушко и И. А. Мысливец (1989) это установи-
ли на основании анализа содержания кислорода в озерах Белоруссии.
Учитывая многообразие типов озер, Г. С. Шилькрот (1989) выделяет
следующие механизмы устойчивости: проточность водоема, емкость
водной массы, буферная емкость отложений, буферная емкость мак-
рофитов, пространственное разнообразие экосистем водоема. Заме-
тим, что вышеперечисленные механизмы нашли отражение в появив-
шихся в семидесятых годах полуэмпирических моделях известных
лимнологов Фолленвайдера, Диллона и Риглера. Достоинство этих
моделей заключалось в том, что они позволяли по изменению нагруз-
ки озер питательными веществами в приближенной форме прогнози-
ровать изменение состояния водоемов, различающихся скоростью
водообмена и глубиной. Однако эти модели не учитывают внутрен-
ней биогенной нагрузки, поэтому они применимы не всегда. По
72

Г. С. Шилькрот (1989), „если для олиготрофно-мезотрофных водоемов
основные механизмы устойчивости - это проточность и емкость
системы по отношению к поступающим веществам, то для эвтроф-
ных - это способность самоочищения, регулируемая скоростью про-
дукционно-деструкционных процессов и гидродинамикой".
Таким образом, в настоящее время достаточно аргументированно
сформулирована проблема устойчивости экосистем. Когда мы эту
проблему пытаемся решить применительно к антропогенному воз-
действию, нужно четко понимать, что это устойчивость к конкретным
воздействиям. Хотя проблема устойчивости сформулирована, все
еще слабым местом этой проблемы является определение критериев
устойчивости. Используемые критерии имеют серьезные недостатки.
Но тем не менее уже сейчас можно перейти к оценке устойчивости
систем: если мы говорим о системах со слабыми связями (популя-
ции, биоценозы), то здесь используются структурные и организацион-
ные критерии, если же обо всей озерной системе, то здесь практи-
чески нельзя решить проблему, не использовав функциональные
критерии. На наш взгляд, для установления критерия устойчивости
необходимо взять такой показатель, который бы наиболее полно
характеризовал сущность водоема; это прежде всего интенсивность
круговорота органического вещества, на который замыкаются прак-
тически все лимнические процессы.
Интенсивность круговорота органического вещества может ха-
рактеризоваться соотношением продукционно-деструкционных про-
цессов. Скорость деструкционных процессов в значительной степени
определяется активностью бактериопланктона, в частности интенсив-
ностью бактериальной продукции. Исследования, выполненные
Институтом озероведения РАН на озерах Латгальской возвышенно-
сти, показали, что с нарастанием уровня трофии на первых этапах
возрастало отношение бактериальной продукции к первичной про-
дукции фитопланктона - от 6.5 до 22-38
%
(Драбкова и др., 1989).
Р
о6<ц>
мг
/
л
Рис. 15. Зависимость первичной продукции фитопланктона А (темные точки)
и бактериальной продукции P (светлые точки) от содержания общего фосфора
в воде озер Латгалии.
73

Это наблюдалось при увеличении первичной продукции от 40 до
144 гС/м
2
за вегетационный период. Дальнейшее увеличение первич-
ной продукции не сопровождалось аналогичным увеличением бак-
териальной продукции, отношение этих показателей уменьшалось до
4-12% при максимальных величинах первичной продукции 230-
280 гС/м
2
за вегетационный период (рис. 15). Уменьшение интенсив-
ности деструкционных процессов по сравнению с продукционными
обычно отмечалось при концентрациях общего фосфора около 0.06-
0.08 мг/л. Превышение этих концентраций приводит к нарушению
равновесия рассматриваемых озерных экосистем. Такие нарушения
проявляются в отсутствии баланса между выделением и потреб-
лением кислорода, между образованием и распадом органического
вещества, в накоплении органического вещества в озерах, сокраще-
нии защитных свойств экосистем.
Очевидно, для решения практических задач возможно рекомен-
довать в качестве критерия устойчивости соотношение продукцион-
но-деструкционных процессов.
3.2. Методы оценки устойчивости озерных экосистем
Установление пределов устойчивости существования экоси-
стем - актуальная задача, решение которой необходимо не только
для создания теории эвтрофирования, но и для решения ряда прак-
тических вопросов. Пока мы располагаем очень небольшим количест-
вом данных, характеризующих поступление в озера таких концентра-
ций биогенных элементов, которые приводят к нарушению сбаланси-
рованности экосистем озер.
Пределы устойчивости экосистем в значительной степени опреде-
ляются изменчивостью внешней среды. Пределы устойчивости озер-
ных экосистем умеренной зоны выше, чем субтропиков и тропиков.
Вот почему озера тропической зоны наиболее подвержены антропо-
генному воздействию, несмотря на то что для экосистем этих озер
характерно наличие сильных и разносторонних связей. Последнее
в соответствии с гипотезой эволюционного времени подтверждает
наличие прямого соотношения между разнообразием системы и ее
возрастом (Thiery, 1982). По мере старения экосистем устойчивость ее
возрастает и достигает своего максимума в зрелых экосистемах.
Примером озерных экосистем, не достигших устойчивости, обычно
называют озера Крайнего Севера. Однако, как показали исследова-
ния Института озероведения РАН, это не совсем так. Здесь озера мож-
но четко разделить по происхождению на ледниковые и термокарсто-
вые, эти озера имеют разный возраст. Наиболее молодыми являются
современные термокарстовые мелководные озера, возникшие в ре-
зультате вытаивания льда из толщи минеральных грунтов или мерз-
лых бугристых торфяников. Эти озера действительно имеют неста-
бильную экосистему, что сказывается на видовом разнообразии
планктонных и бентосных сообществ, которые значительно ниже, чем
74

в глубоководных озерах ледникового происхождения, чей возраст
определяется несколькими тысячами лет. Эти озера могут служить
хорошим примером нестабильных и стабильных экосистем. Фито-
планктон в первых представлен 30-56 видами против 105-127; во вто-
рых зоопланктон представлен соответственно 5-8 и 19-36 видами, а
простейшие соответственно - 4-14 и 18-28 видами (Особенности...,
1992). Эти две группы озер различаются по функциональным показате-
лям гидробионтов, прежде всего - по соотношению продукционно-де-
струкционных процессов. В мелководных термокарстовых озерах,
как правило, величины первичной продукции (Ф) значительно пре-
вышают скорость деструкции органического вещества (Д), коэффи-
циент Ф/Д в абсолютном большинстве озер был больше единицы.
Следовательно, эти озера способны к быстрому накоплению в толще
воды органического вещества, создаваемого фитопланктоном и, сле-
довательно, более уязвимы к эвтрофированию при поступлении до-
полнительных биогенных элементов. В глубоководных озерах со
сбалансированными экосистемами наблюдалось преобладание
деструкционных процессов над продукционными (коэффициент Ф/Д
был меньше единицы). В более зрелых озерных экосистемах антропо-
генное воздействие (поступление буровых растворов) приводит
к структурным и организационным перестройкам. Уменьшаются чис-
ло видов и видовое разнообразие. Особенно четко это наблюдается
в зоопланктонном сообществе, число видов которого уменьшается
с 19-36 в природночистых озерах до 10-13 в загрязняемых. Число
видов простейших также уменьшается с 18-28 до 5-11 соответствен-
но, эта тенденция проявляется и у фитопланктона. Функциональные
же показатели, и прежде всего соотношение продукционно-деструк-
ционных процессов в этой группе озер, практически не меняются.
Обратная картина наблюдалась в неустойчивых молодых экосисте-
мах - практически отсутствовали структурные и организационные
перестройки, но четко проявлялись функциональные изменения -
повышение интенсивности продукционных процессов не сопровожда-
лось повышением деструкционных. Намечалось еще более четкое
отставание деструкционных процессов от продукционных. Все это
указывает на то, что мелководные озера Болыыеземельской тундры,
а их большинство в этом районе, в наибольшей степени реагируют на
антропогенное воздействие, поскольку оно затрагивает функцио-
нальные особенности озер, что отрицательно сказывается на их жиз-
неспособности.
Как отмечалось в предыдущем разделе, хорошим критерием
устойчивости может быть соотношение продукционно-деструкцион-
ных процессов. Было показано, что в довольно глубоких озерах, где
основным источником органического вещества является фитопланк-
тон, уменьшение показателей гетеротрофной активности микроорга-
низмов начинается с концентрации 0.06-0.08 мг/л общего фосфора (см.
рис. 15). Указанные концентрации фосфора можно принять за тот пре-
дел, превышение которого может привести к нарушению устойчивости
75
Озера.
Олиготрофные Мезатросрные
ЗВтро/рные
CanpompaipHbie
штаВое,
I
Ii
сельскохозяист
—Ценное
Органическое Вещество
I £
Общее количест8о бактерий мл -10
-inn-pnn
ZM
r
^oT
П ?.Ч-П R4
Гете
атибность
органичеся
бак
ютротная —
глюкоза/мкг/л/ч
р -| ~~1Ch20
Деструкция —
ого Вещества,гС/м/г^ч
1.9-3.1 —
0.01-0.03
Гете
атибность
органичеся
бак
•
— 'нроОукция
териальнпй.
биомассы
Самоочищение
_ О.ОБ-0.25
ч
0.03-0.18
ч
0.03-0.18
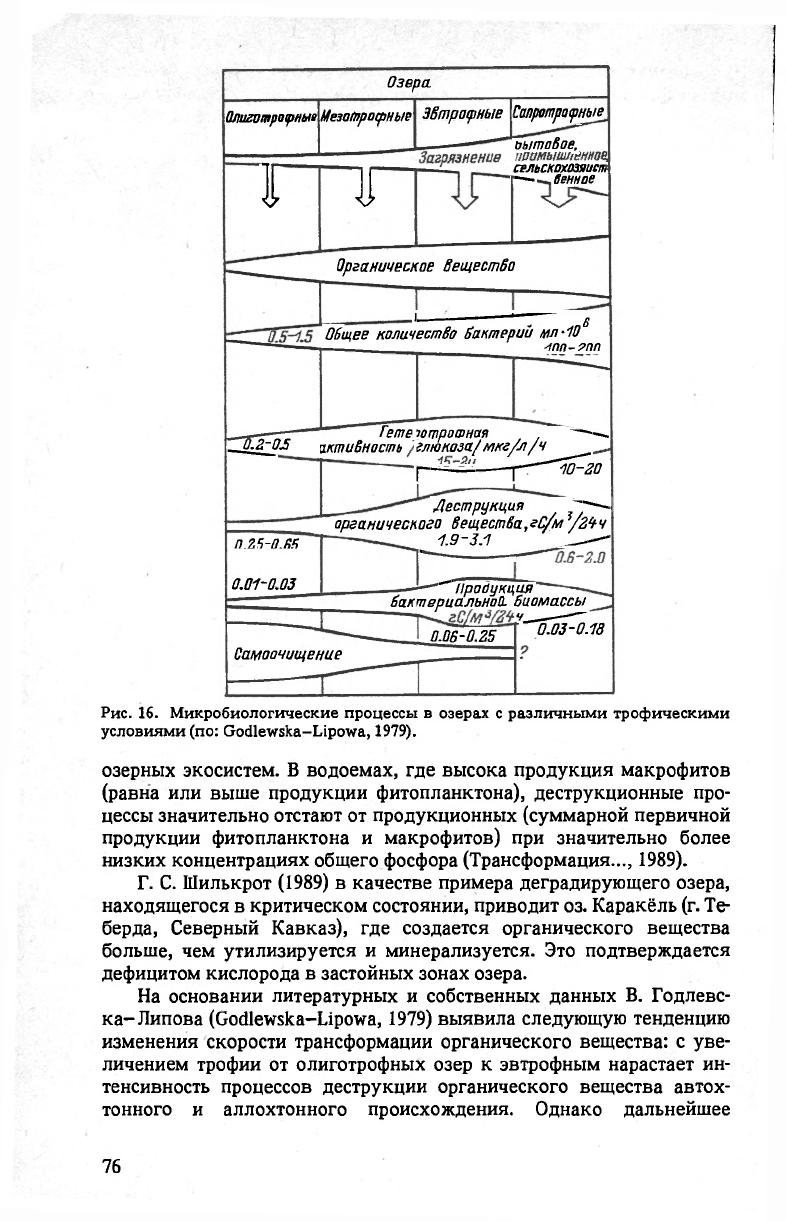
Рис. 16. Микробиологические процессы в озерах с различными трофическими
условиями (по: Godlewska-Lipowa, 1979).
озерных экосистем. В водоемах, где высока продукция макрофитов
(равна или выше продукции фитопланктона), деструкционные про-
цессы значительно отстают от продукционных (суммарной первичной
продукции фитопланктона и макрофитов) при значительно более
низких концентрациях общего фосфора (Трансформация..., 1989).
Г. С. Шилькрот (1989) в качестве примера деградирующего озера,
находящегося в критическом состоянии, приводит оз. Каракёль (г. Te-
берда, Северный Кавказ), где создается органического вещества
больше, чем утилизируется и минерализуется. Это подтверждается
дефицитом кислорода в застойных зонах озера.
На основании литературных и собственных данных В. Годлевс-
ка-Липова (Godlewska-Lipowa, 1979) выявила следующую тенденцию
изменения скорости трансформации органического вещества: с уве-
личением трофии от олиготрофных озер к эвтрофным нарастает ин-
тенсивность процессов деструкции органического вещества автох-
тонного и аллохтонного происхождения. Однако дальнейшее
76

увеличение уровня трофии под влиянием индустриальных, комму-
нальных и сельскохозяйственных стоков влечет за собой ингибиро-
вание этих процессов, резкое замедление самоочищения вод
(рис. 16).
Нарушение соотношения продукционно-деструкционных про-
цессов естественно отражается на поведении кислорода в воде озер
в годичном цикле, поэтому возможно выразить устойчивость лимно-
систем через изменение запасов кислорода в столбе воды пло-
щадью 1 м
2
(г/м
2
) в точке максимальной глубины (Якушко, Мысли-
вец, 1989). Содержание кислорода в столбе воды определялось по
формуле
О = Oi Vi + O
2
Vj
8
+ Оз Уз+ - + On V
n
^
maxj
где О], О2, ..., O
n
- содержание кислорода в определенном слое во-
ды, г/м
3
; V
1
, V
2
, ..., V
n
- объемы слоев воды с сечением изобат в
1 м, млн. м
3
; V- общий объем воды в озере, млн. м
3
; Jj
max
- глубина
максимальная, м.
Такой расчет позволяет проследить связь между количеством
кислорода и глубиной озера, стратифицированностью его водной
массы и сравнивать озера с разными морфометрическими параметра-
ми. Расчеты проводились для периодов летней и зимней стагнации
в разнотипных озерах с разной степенью антропогенного воздейст-
вия (АВ). Отношение между запасами кислорода (в г/м
2
) за опреде-
ленный промежуток времени принято за показатель изменения эко-
системы по кислороду (КУ).
Расчеты показали, что запасы кислорода за промежуток времени
в несколько лет могут колебаться в значительных пределах: от
нескольких грамм на квадратный метр до сотен. При условии высо-
кого запаса кислорода во все сезоны года устойчивость экосистемы
сохраняется. Резкое падение кислородных запасов указывает на
неспособность системы сопротивляться АВ.
При условии интенсивного проявления AB наименее устойчи-
выми оказались мезотрофные с признаками олиготрофии глубокие
и высокоэвтрофные мелководные озера. Это было выявлено на при-
мере озер Белоруссии. Если в естественном состоянии экосистемы
запасы кислорода в первом типе достигают 600-400 г/м
2
(оз. Долгое),
а величина КУ около единицы, то в оз. Лесковичи при интенсивном
поступлении трофогенных веществ содержание кислорода в летнюю
стагнацию опускается до 100-90 г/м
2
, а в гиполимнионе достигает
нуля. При этом показатель КУ > 1. Чувствительность этого типа
к антропогенным воздействиям соответствует воронкообразной кот-
ловине, слабому ветровому перемешиванию и мощному гиполим-
ниону, на долю которого приходится до 0.5 объема водной массы -
явление, получившее название „эффекта морфометрии" (табл. 8).
Слабая устойчивость высокоэвтрофных озер связана с резкими
77
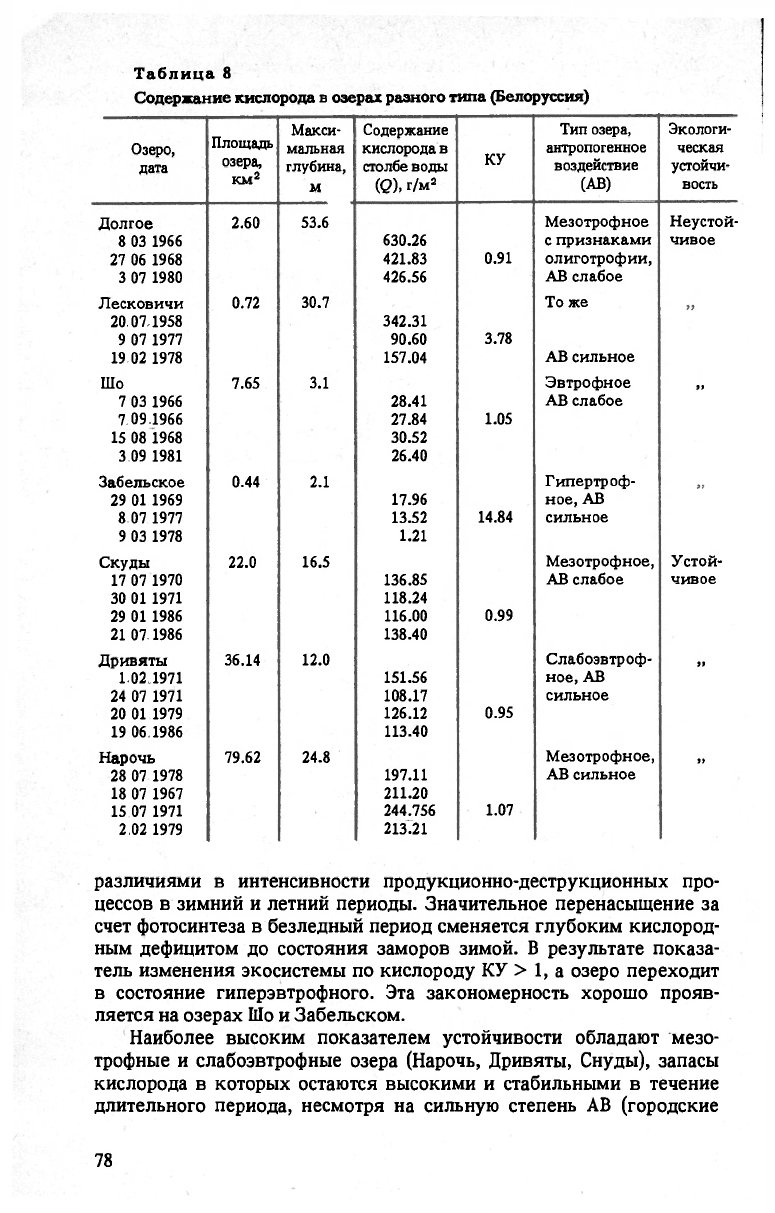
Таблица 8
Содержание кислорода в озерах разного типа (Белоруссия)
Озеро,
дата
Площадь
озера,
KM
2
Макси-
мальная
глубина,
M
Содержание
кислорода в
столбе воды
(Q), г/м
2
КУ
Тип озера,
антропогенное
воздействие
(AB)
Экологи-
ческая
устойчи-
вость
Долгое
8 03 1966
27 06 1968
3 07 1980
2.60
53.6
630.26
421.83
426.56
0.91
Мезотрофное
с признаками
олиготрофии,
AB слабое
Неустой-
чивое
Лесковичи
20.07.1958
9 07 1977
19 02 1978
0.72
30.7
342.31
90.60
157.04
3.78
То же
AB сильное
Шо
7 03 1966
7 09 .1966
15 08 1968
3 09 1981
7.65
3.1
28.41
27.84
30.52
26.40
1.05
Эвтрофное
AB слабое
»
Забельское
29 01 1969
8.07 1977
9 03 1978
0.44
2.1
17.96
13.52
1.21
14.84
Гипертроф-
ное, AB
сильное
Скуды
17 07 1970
30 01 1971
29 01 1986
21 07 1986
22.0 16.5
136.85
118.24
116.00
138.40
0.99
Мезотрофное,
AB слабое
Устой-
чивое
Дривяты
1.02 1971
24 07 1971
20 01 1979
19 06.1986
36.14
12.0
151.56
108.17
126.12
113.40
0.95
Слабоэвтроф-
ное, AB
сильное
»
Нарочь
28 07 1978
18 07 1967
15 07 1971
2.02 1979
79.62 24.8
197.11
211.20
244.756
213.21
1.07
Мезотрофное,
AB сильное
>1
различиями в интенсивности продукционно-деструкционных про-
цессов в зимний и летний периоды. Значительное перенасыщение за
счет фотосинтеза в безледный период сменяется глубоким кислород-
ным дефицитом до состояния заморов зимой. В результате показа-
тель изменения экосистемы по кислороду КУ > 1, а озеро переходит
в состояние гиперэвтрофного. Эта закономерность хорошо прояв-
ляется на озерах
Шо
и Забельском.
Наиболее высоким показателем устойчивости обладают мезо-
трофные и слабоэвтрофные озера (Нарочь, Дривяты, Снуды), запасы
кислорода в которых остаются высокими и стабильными в течение
длительного периода, несмотря на сильную степень AB (городские
78

и промышленные сбросы, рекреация). Постоянство режима обеспе-
чивается значительными объемами воды, слабой стратифицирован-
ностью и интенсивностью ветрового перемешивания, обеспечиваю-
щих окислительную среду в течение года. Величина КУ в таких озе-
рах близка к единице.
Предлагаемый метод оценки устойчивости озерных экосистем
следует рассматривать с точки зрения восстановления озер и носит
практический характер. Он учитывает индивидуальные особенности
каждого озера, его принадлежность к генетическому типу.

4. ПУТИ ВОССТАНОВЛЕНИЯ ЭКОСИСТЕМ
МАЛЫХ ОЗЕР
4.1. Принципы и методология
Основу научной концепции восстановления озерной экосистемы
составляют такие понятия,как природный потенциал экосистемы,
трофический статус водоема и устойчивость его экосистемы.
Природный потенциал озера оценивается величиной биогенной
нагрузки на озеро, соответствующей естественному состоянию водо-
сбора. В лесной зоне это будет сплошь облесенный водосбор, а в степ-
ной зоне - сплошь покрытый травой. Знание природного потенциала
озера необходимо для оценки минимального значения биогенной
нагрузки, к которому можно стремиться при восстановлении эко-
системы озера и которого при современных климатических условиях
при данной морфометрии озера и хозяйственном использовании его
водосбора практически нельзя достичь. Поэтому при разработке
мероприятий по восстановлению экосистемы озера нельзя ставить
задачу, связанную со снижением биогенной нагрузки до ее природ-
ных значений.
Концепция повышения трофического статуса озера с увеличени-
ем биогенной (фосфорной) нагрузки широко используется в совре-
менный период для оценки критических значений нагрузки, соот-
ветствующих переходу озера из одного трофического состояния
в другое. Это известная модель Р. Фолленвайдера (Vollenweider, 1975,
1976), который при использовании данных наблюдений на 20 озерах
установил связь между ежегодным поступлением биогенных ве-
ществ и средней глубиной озера (рис. 17) при его соответствующем
трофическом состоянии. Предполагалось, что поступающий фосфор
немедленно перемешивается в озере (для периода летней стратифи-
кации это неприемлемо, но в годовом разрезе это допустимо), и
поэтому его концентрация в истоке из озера равна преобладающему
ее значению в озере; скорость осаждения фосфора пропорциональна
его концентрации в озере, а сезонные колебания биогенной нагрузки
пренебрежимо малы. Эта модель наиболее применима для расчета
стационарного режима или межгодовых изменений. При таком допу-
щении определяется для стационарного режима осредненная кон-
центрация фосфора в озерной воде в виде выражения
р =
L(I-R
y
) (4.1)
80