Caballero B. (ed.) Encyclopaedia of Food Science, Food Technology and Nutrition. Ten-Volume Set
Подождите немного. Документ загружается.


B
BACILLUS
Contents
Occurrence
Detection
Food Poisoning
Occurrence
R Leuschner, Central Science Laboratory, York, UK
Copyright 2003, Elsevier Science Ltd. All Rights Reserved.
Systematics and Ecology of
Bacillus
Species
0001 Aerobic endospore-forming bacteria are currently
assigned to four genera in the family Bacillaceae.
Within this family, the genus Bacillus was established
to include the rod-shaped bacteria that grew in the
presence of air, thus distinguishing them from the
strictly anaerobic Clostridium spp. It is possible to
allocate many Bacillus species to one of six taxa that
have distinguishable physiologies and this is generally
consistent with the devision of the genus based on
spore morphologies. The six groups are: B. polymyxa
group (I), B. subtilis group (II), B. brevis group (III),
B. sphaericus group (IV), and thermophiles (V and
VI). In group I all species are facultative anaerobes
and grow strongly in the absence of oxygen. Acid is
produced from a variety of sugars. Endospores are
ellipsoidal. The group II species (B. subtilis group) are
phylogenetically and phenetically consistent. B. sub-
tilis is an appropriate representative of the taxon that
includes many common names. All these bacteria
produce acids from a range of sugars and some,
B. cereus and B. licheniformis, are facultative anaer-
obes. B. licheniformis grows poorly anaerobically
and can use glucose only under anaerobic conditions.
Although B. subtilis is generally regarded as an
aerobe, it can grow and sporulate slowly under strict
anaerobic conditions. Given glucose, with nitrite as a
terminal electron acceptor, it grows strongly anaero-
bically. These bacteria are therefore an intermediate
stage between the true facultative anaerobes of the
group I strains and the strict aerobes in groups III and
IV. This is reflected in their production of acid from
several sugars. All these bacteria produce oval endo-
spores that do not swell the mother cell and are
generally located centrally or subterminally. Group
III represents strict aerobes that generally do not pro-
duce acid from sugars, with the exception of two
species. They produce ellipsoidal spores that swell
the mother cell. In group IV all species produce spher-
ical spores that may swell the mother cell and contain
l-lysine or ornithine in the cell wall. All species
are strictly aerobic, but some have limited ability
to produce acid from sugars. Group V represents
thermophilic species that grow optimally at > 50
C.
Physiologically and morphologically, they are hetero-
geneous, but most produce oval spores that swell the
mother cell. In group VI are thermophilic, acidophilic
species with membraneous o-alicyclic fatty acid.
Occurrence in the Environment and Food
0002Members of the genus Bacillus have a ubiquitous
environmental distribution. The endospore is import-
ant for the dispersion of Bacillus spp. Spores are
readily blown about in dust and air currents and
are prevalent in animal feces. Bacteria related to
B. subtilis are commonly encountered and easily iden-
tified. The soil is the reservoir of these bacteria. From
the soil, they are transferred to various associated
environments, including plants and plant materials,
foods, animals, and marine and fresh-water habitats.
B. cereus, B. licheniformis, B. pumilus,andB. subtilis
are prevalent in soils, particularly low-nutrient soils.
They are also common on straw and cereals, includ-
ing rice and pulses, which they presumably colonize

from wind-blown soil particles and dust. Endospores
of bacilli represent a metabolically inactive survival
form. They are characterized by high resistance
against heat, dryness, irradiation, and other unfavor-
able environmental conditions. The success of the
endospore for the survival of the species depends on
an effective mechanism to resuscitate and enter the
vegetative cell cycle to multiply. It is usually the case
that spores resuscitate under good environmental
conditions that allow germination and cell growth.
Food of various origin and compositions can offer
these conditions and this can lead to spoilage or
even food poisoning due to growth of bacilli in food.
0003 Spores and vegetative cells of Bacillus cereus and
other food-poisoning Bacillus spp. occur widely in
soils (10
5
–10
6
CFU g
1
) and may be found in raw
materials and processed foods which were not steril-
ized by heat or irradiation. They constitute a major
portion of the microbial flora of raw milk and easily
contaminate various dairy products, causing spoilage
with their proteolytic, lipolytic, and saccharolytic ac-
tivities. B. cereus was detected and enumerated in
milk, vegetable, and meat-based products (Table 1).
B. cereus is a problem to the dairy industry because it
contaminates the udders of cows while they graze in
the fields and can be introduced into milk. B. licheni-
formis, B. pumilus and B. subtilis were the most
commonly isolated Bacillus species in bakeries and
milk (Tables 2 and 3). The incidence of B. cereus and
B. subtilis in various food products was investigated
in the Netherlands. The total number of products was
229 samples and B. cereus and B. subtilis were isol-
ated at contamination levels between 10
2
and 10
6
CFU g
1
or ml in a wide range of food products such
as milk, yeast, flour, pasta products, cocoa, chocolate,
bakery products, meat products, and herbs and
spices. B. cereus was present in 48% and B. subtilis
tbl0001 Table 1 Prevalence of Bacillus cereus in raw and processed
food products
Foodproduct No. of
samples
examined
No. positive
for B. cereus
(%)
B. c ereus
counts g
1
or ml
1
Rice and oriental foods
Raw rice 13 46 10
2
–10
3
Boiled rice 32 38 10
3
–10
7
14 93 10
1
–10
3
252 10 10
2
–10
5
Fried rice 14 86 10
1
–10
3
204 24 10
2
–10
5
Egyptian rice dishes 172 40 10
1
–10
4
Japanese noodles 200 8 10
1
–10
4
Soybean curd (tofu) 257 56 10
1
–10
4
467 31 10
1
–10
6
Sashimi (raw fish) 228 14 10
2
–10
3
Milk and dairy products
Raw milk 100 9 10
1
–10
2
Pasteurized milk 100 35 10
1
–10
3
Milk powder 120 27 10
1
–10
3
Icecream 100 48 10
1
–10
3
Spices 110 53 10
1
–10
4
Meat and meat products
Raw meats 133 2 10
1
–10
2
452 6 10
1
–10
2
Modified from Kramer JM and Gilbert RJ (1989) Bacillus cereus and other
Bacillus species. In: Doyle MP (ed.) Foodborne Bacterial Pathogens, pp. 21–
70. New York: Marcel Dekker.
tbl0002Table 2 Distribution of 170 bakery Bacillus isolates identified
using API 50CHB
Isolate Bread ingredients
and uncooked
dough (%)
Swabs of
processing
line (%)
Cooked
loaves and
bread
crumbs (%)
B. subtilis 22.9 4.1 5.3
B. amyloliquefaciens 1.2 1.2 0
B. li cheniformi s 18.2 5.9 4.1
B. p umilus 11.2 2.4 2.4
B. circulan s 2.9 0.6 0.6
B. me gaterium 1.8 1.2 0.6
B. p olymyxa 2.9 0 0.6
B. macera ns 2.4 0 0.6
B. c ere u s (including
B. mycoides)
0.6 1.8 0
B. stearothermophilus 0 0 0.6
Unacceptable profile 0.6 2.4 1.2
Reproduced from Thompson JM, Dodd CER and Waites WM (1993)
Spoilage of bread by Bacillus. International Biodeterioration and
Biodegradation 32: 55–66, with permission.
tbl0003Table 3 Mesophilic Bacillus species isolated from milk
samples
Bacillus species No. of sampleswhere a specieswas
isolated (total number of samples: n ¼250)
B. li cheniformi s 131
B. p umilus 156
B. subtilis 127
B. lentus 52
B. pantothenticus 33
B. amyloliquefaciens 25
B. stearothermophilus 19
B. mycoides 22
B. c ere u s 16
B. circulan s 15
B. firmus 10
B. me gaterium 27
B. shaericus 6
B. macera ns 3
B. later o sporus 2
B. p olymyxa 5
Unknown 120
Reproduced from Sutherland AD and Murdoch R (1994) Seasonal
occurrence of psychrotrophic Bacillus species in raw-milk, and studies on
the interactions with mesophilic Bacillus sp.
356
BACILLUS
/Occurrence

in 25% of all samples examined. In spoiled bread
that developed symptoms of ropiness after 2 days’
storage at ambient summer temperatures, identified
species were B. subtilis (70%), B. licheniformis (24%),
B. pumilus (2%), and B. cereus (2%).
Food Spoilage and Poisoning
0004 The occurrence of Bacillus species in raw materials
used for food processing is generally below the infec-
tious dosis required to affect negatively the well-being
of consumers: this is normally above 10
5
cells g
1
or
ml
1
food (Table 4). This indicates that Bacillus cells
or spores must have the opportunity to multiply in the
food environment to enable them to cause food spoil-
age and poisoning.
0005 B. cereus food poisoning is principally associated
with the storage of cooked foods at temperatures and
times that allow growth and result in cell numbers
above 10
6
CFU g
1
or ml
1
. Rapid cooling of cooked
food below 10
C is generally an effective control
measure. Low-pH foods (< pH 5.0) and dry foods
will not support the growth of B. cereus, although
many dried foods will be contaminated with spores of
this microorganism. Spores are more hydrophobic
than spores of other Bacillus spp., which enables
them to adhere particularly well to many types of
surfaces. This makes them difficult to remove during
cleaning and a difficult target for disinfection.
0006 B. cereus can cause infections and intoxications. In
addition to foodborne diseases, B. cereus causes septi-
cemia, meningitis, and ocular infections. It can cause
two types of food poisoning: the emetic disease, char-
acterized by a short incubation period (1–5 h) display-
ing symptoms such as nausea, vomiting, and stomach
cramps, and the diarrheal disease, characterized by an
incubation period of 8–16 h with symptoms including
abdominal pain, watery diarrhea, and rectal tenes-
mus. Usually both types of food poisoning are rela-
tively mild and last for less than 24 h.
0007 B. subtilis was reported to be the cause of illness in
meat dishes with elements of vegetables, seafood and
rice, bread and pastry products, sandwiches, and
pizza. The infectious dose was > 10
5
CFU g
1
with
an incubation period of 10 min to 14 h and a duration
of illness of 2–8 h. Symptoms were vomiting (80%),
diarrhea (49%), abdominal pain/cramps (27%), and
nausea, headaches, flushing, and sweating. Bacilli are
capable of causing food spoilage in bread, and this is
known as ropiness. B. subtilis has been described to
be the main cause of the bread spoilage ‘ropiness.’
Ropiness is the most important spoilage of bread after
moldiness and occurs particularly during the summer
when the climatic conditions favor growth of the
bacteria. This involves mainly B. subtilis. Bacillus
counts in white and wholemeal wheat loaves pro-
duced without preservatives or sourdough were con-
sistently 10
6
CFU g
1
after 2 days of storage at
ambient summer temperatures.
0008B. licheniformis was identified in meat dishes with
elements of vegetables, bread, and pastry products,
and chicken as a cause of food poisoning. The infec-
tious dose was > 10
6
CFU g
1
, the incubation period
2–14 h and the duration of illness 6–24 h. Symptoms
were vomiting (54%), diarrhea (92%), and abdom-
inal pain/cramps (46%). B. pumilus in meat products,
sandwiches, and canned tomato juice was found to be
responsible for food poisoning. The infectious dose
was > 10
6
CFU g
1
and the incubation period 15 min
to 11 h. Symptoms were vomiting and diarrhea.
0009B. stearothermophilus and B. coagulans were
reported to be the cause of flat sour spoilage in evap-
orated milk.
Growth Characteristics and Resistance of
Bacilli
0010Knowledge about the resistance and growth charac-
teristics of Bacillus species is important to enable food
manufacturers and consumers to avoid handling
practice that enable spores to germinate and vegeta-
tive cells to multiply in a food.
0011Psychrotrophic strains of B. cereus were able to
grow and produce diarrheal toxin at temperatures
down to 4
C. Most mesophilic strains are able to
grow in low-acidic foods at temperatures between
15 and 55
C, with an optimum range of 30–40
C.
Growth can generally occur between pH 5.0 and 8.8,
tbl0004 Table 4 Infective or intoxication dosis food poisoning caused by Bacillus species
Bacillus species Intoxication dosis Infective dosis
B. c ere u s (emetic toxin) 12–32 mgkg
1
toxin (Suncus murinus: a small monkey) 10
5
–10
8
cells g
1
food
B. c ere u s (diarrheal disease) Usual 10
5
g
1
or ml
1
B. subtilis >10
5
B. li cheniformi s >10
6
B. p umilus >10
6
Modified from Granum PE and Baird-Parker TC (2000) Bacillus species. In: Lund BM, Baird-Parker TC and Gould GW (eds) The Microbiological Safety and
Quality of Food, vol. 2, pp. 1029–1039. Maryland: Aspen.
BACILLUS
/Occurrence 357

with an optimum between 6.0 and 7.0. Spores are
variable in their heat resistance, which is normally
moderate, but can be increased by fat components.
The emetic toxin is very heat-resistant and can survive
heating at 126
C for 90 min. The diarrheal toxin is
heat-sensitive and is inactivated at 56
C after 5 min.
Growth of B. cereus was found at water activities of
0.93 depending on the acidulant and humectant.
Other food-poisoning Bacillus spp. have similar
growth and resistance characteristics to B. cereus. B.
licheniformis and B. subtilis were not found to grow
at pH 4–4.2; however, growth in tomato juice at a pH
of 4.4 in the presence of oxygen was reported. Spores
of thermophilic bacilli such as B. stearothermophilus
are far more heat-resistant and are used to test steril-
ization effectiveness. Heat resistance of bacilli spores
is dependent on the temperature during spore forma-
tion and on the pH of the food. Further dependence
was observed on various acidulants (Table 5).
0012 Sterilization will inactivate all Bacillus spores. This
is however also imposing a harsh treatment towards
other valuable food ingredients. The development of
novel and milder food preservation technologies, such
as high-pressure and electromagnetic fields, have
proven to be effective in Bacillus spore inactivation.
Pressure treatments of 400 MPa for 25 min at 30
C
resulted in a 0.45 log inactivation of B. cereus spores.
Pressure treatments at lower temperatures were less
effective. Spores of B. subtilis proved to be more
resistant to ultraviolet B range (280–330 nm) than
spores of B. cereus. An exposure of 30 min reduced
the viability of spores of B. cereus by 50% and that
of B. subtilis spores by 10%. Levels of resistance to
ultraviolet B of spores from different Bacillus species
appear to be related to the quantity and quality of
small acid-soluble proteins and to activities of DNA
repair systems. The novel, milder nonthermal preser-
vation technologies need further evaluation with par-
ticular emphasis on spore inactivation mechanisms
and synergistic effects by combination of various
techniques in food systems.
See also: Bacillus: Detection; Food Poisoning; Food
Poisoning: Classification; Spoilage: Bacterial Spoilage
Further Reading
Claus D and Berkeley RCW (1986) Genus Bacillus. In:
Sneath PHA (ed.) The Bergey’s Manual of Systematic
Bacteriology, vol. 2, pp. 1105–1139. Baltimore: Wil-
liams & Wilkins.
Granum PE and Baird-Parker TC (2000) Bacillus species.
In: Lund BM, Baird-Parker TC and Gould GW (eds) The
Microbiological Safety and Quality of Food, vol. 2, pp.
1029–1039. Maryland: Aspen.
International Commission on Microbiological Specifica-
tions for Foods (1996) Bacillus cereus. In: Roberts TA,
Baird-Parker AC and Tompkin RB (eds) Microbiological
Characteristics of Food Pathogens, pp. 335–344.
London: Blackie Academic and Professional.
Kramer JM and Gilbert RJ (1989) Bacillus cereus and other
Bacillus species. In: Doyle MP (ed.) Foodborne Bacterial
Pathogens, pp. 21–70. New York: Marcel Dekker.
Priest FG (1993) Systematics and ecology of Bacillus. In:
Sonenshein AL, Hoch JA and Losick R (eds) Bacillus
Subtilis and Other Gram Positive Bacteria, pp. 3–16.
Washington, DC: American Society of Microbiology.
Detection
L A Shelef, Wayne State University, Detroit, MI, USA
Copyright 2003, Elsevier Science Ltd. All Rights Reserved.
Background
0001The genus Bacillus is large, comprising more than 60
species that are mostly saprophytes, widely distrib-
uted in nature, spreading from soil to water, plants,
and animals. The genus shows a great diversity of
strains and species. The organisms are Gram-positive
or Gram-variable spore-forming bacilli, mostly cata-
lase-positive, that may be motile by peritrichous
flagella. Most are mesophiles, but some are psychro-
throphs and thermophiles. Bacillus contains strict
aerobes (e.g., B. megaterium), as well as facultative
anaerobes (e.g., B. cereus, B. licheniformis). The
vegetative cells are rods ranging from 0.5 1.2 mm
to 2.5 10 mm, and the endospores are in the central
or paracentral, subterminal or terminal position. Sur-
vival of the organism results from the resistance of the
spores to adverse conditions.
0002Of the identification schemes proposed, that of
Gordon and co-workers divides the genus Bacillus
into three groups, according to cellular morphology
and physiological properties. Group 1 consists of
tbl0005 Table 5 Decimal reduction (D) values (min) at 95
CofBacillus
cereus spores for various pH and organic acid types
pH Citric acid Lactic acid Acetic acid Malic acid
6.5 1.03 1.09 1.21 0.96
6 0.95 0.85 0.99 0.96
5.5 0.81 0.82 0.82 0.77
5 0.74 0.66 0.67 0.69
4.5 0.64 0.54 0.55 0.62
4 0.62 0.40 ND 0.59
ND, not determined.
Modified from Leguerinel I and Mafart P (2001) Modelling the influence of
pH organic acid types on thermal inactivation of Bacillus cereus spores.
International Journal of Food Microbiology 63: 29–34.
358
BACILLUS
/Detection

Gram-positive species that form ellipsoidal or cylin-
drical spores that do not appreciably distend the spor-
angia. This group is further divided according to the
vegetative cell dimensions and presence of lipid glob-
ules in the protoplasm of the species. The ‘large-
celled’ group 1 Bacillus species have a cell width
greater than 0.9 mm and include B. megaterium,
B. cereus, B. cereus var. mycoides, B. thuringiensis,
and B. anthracis. The ‘small-celled’ species have a cell
diameter < 0.9 mm, and lipid globules are not formed.
They include B. subtilis, B. pumilus, B. licheniformis,
and B. coagulans. Organisms of groups 2 and 3 are
characterized by swollen sporangia and either ellips-
oidal (group 2) or spherical (group 3) spores. Studies
of DNA composition and other studies suggest that
the genus Bacillus consists of some bacterial families
that will necessitate further classification of the genus.
0003 Most Bacillus species are harmless saprophytes
characterized by their proteolytic and saccharolytic
properties and are rarely associated with human
or animal disease. However, several species are
proven or suspect pathogens. Of these, B. cereus is a
recognized food pathogen, and other species, particu-
larly those of the B. subtilis group (B. subtilis,
B. licheniformis, and B. pumilus), have been impli-
cated in foodborne illnesses. B. brevis, also identified
as a causative agent of foodborne disease, has
been split and transferred to the new genus Breviba-
cillus. B. anthracis is the agent of anthrax.
Five Bacillus species, B. thuringiensis, B. popilliae,
B. sphaericus, B. larvae, and B. lentimorbus, are
insect pathogens, the first three being used in com-
mercial insecticides for the control of crop insects.
0004 Of the Bacillus species associated with foods, de-
tection and identification methods have focused on
B. cereus as a recognized food pathogen. Spores and
vegetative forms of B. cereus are frequent inhabitants
of soils, sediments, dust, natural water sources, vege-
tation, and many foods such as grains, dairy prod-
ucts, dried foods, spices, meats, and vegetables.
Examination of various raw and processed foods
shows contamination levels ranging from 10
1
to
10
6
–10
7
cells per gram or milliliter. It is estimated
that B. cereus accounts for *2–3% of reported food-
borne outbreaks in the USA. In the UK, a single
outbreak was reported during the period of Octo-
ber–December, 2000. In this outbreak, 30 people
became ill after consuming contaminated soup with
cream in a canteen, and a statistical association was
established between consumption of the food and the
cause.
0005 Detection of B. cereus and its toxins and differenti-
ation between the species within the B. cereus group
are presented in this chapter. Following a brief review
of the properties of the organism and its toxins,
traditional and novel detection of the organism and
its toxins are discussed.
Properties of
B. cereus
0006Cells of B. cereus are large, Gram-positive rods that
are motile by means of peritrichous flagella. They are
typically 1.0–1.2 mm in diameter and 3.0–5.0 mmin
length. Endospores are in either the central or para-
central position without distention of the sporan-
gium. The organism sporulates freely on many
media under aerated conditions. The optimum
growth temperature is 28–35
C, with a minimum of
5
C and a maximum of 48
C, and the pH for growth
is 4.9–9.3. B. cereus strains are able to utilize glucose,
fructose, and trehalose. Sucrose, salicin, maltose,
mannose, glycerol m-inositol, and lactose are utilized
by some, but not all, members of the species. Pen-
toses, mannitol, and many of the sugar alcohols are
not metabolized. Urease is produced by few strains.
Most of the strains produce acetylmethylcarbinol
(Voges–Proskauer-positive), reduce nitrate to nitrite,
hydrolyze soluble starch, tyrosine, gelatin and casein,
and produce a neutral metalloprotease. B. cereus and
B. thuringiensis produce a broad spectrum of
b-lactamase and are, therefore, resistant to penicillin,
ampicillin, and cephalosporins. Their ability to grow
on agar containing penicillin aids in differentiating
them from B. anthracis. Certain strains have been
reported to synthesize a dehydropeptide reductase
that catalyzes cleavage of the bacteriocins nisin and
subtilin. In contrast to B. megaterium, B. cereus
strains are not sensitive to lysozyme.
0007The organism elaborates a number of toxins with
distinct diarrheal and emetic syndromes. The diar-
rheal syndrome is caused by bacterial growth and
toxin formation in the small intestine, whereas the
emetic syndrome is caused by preformed toxin
resulting from growth of the pathogen in the food.
Of the enterotoxins responsible for the diarrheal syn-
drome, hemolysin, designated BL, comprises three
heat-labile peptides, B (37.8 kDa), L
1
(38.5 kDa),
and L
2
(43.2 kDa), and the genes that encode for
these components have been cloned. The L
1
and L
2
peptides appear to be responsible for binding to
erythrocytes and the B component for lysing them.
Cereolysin (56 kDa), a cytolytic protein responsible
for hemolysis, consists of phospholipases that hydro-
lyze phosphatidylinositol, some of which are metal-
loenzymes requiring divalent cations for activity, and
sphingomyelinase, also a metalloenzyme that has
hemolysin-like activity, but shows no phospholipase
activity. The nonhemolytic enterotoxin is heat-labile,
comprising three proteins (39, 45, and 105 kDa).
Cereulide is a putative emetic toxin. It has been
BACILLUS
/Detection 359

isolated and identified as a small (1.2 kDa), heat-
stable dodecadepsipeptide, structurally related to the
ionophore valinomycin, producing a vacuole re-
sponse in HEp-2 cells. Psychrophilic strains of B.
cereus capable of producing toxins at refrigerator
temperatures have been reported.
Isolation and Identification of
B. cereus
Standard Agar Media
0008 The characteristics of B. cereus and of the culturally
similar B. mycoides, B. thuringiensis,andB. anthra-
cis are summarized in Table 1. Most procedures for
the isolation, identification, and enumeration of
B. cereus involve direct plating techniques. Media
developed for the identification of the organism pro-
vide conditions for the organism to exhibit fermenta-
tion properties, hemolysin production, lecithinase
activity, and morphological characteristics. Selective
agents to inhibit competitive microorganisms, such as
antibiotics (most often 10 mg of polymyxin per milli-
liter), are normally incorporated into the media.
0009 MYP agar Mannitol–egg yolk-polymyxin agar
(MYP) was the first selective medium, introduced in
1967 by Mossel and coworkers. It relies on lecithin
hydrolysis (the egg-yolk reaction), negative mannitol
reaction for differentiation of B. cereus from other
commonly occurring Bacillus species, and polymyxin
B. The agar plate is surface-inoculated, and typical
colonies of B. cereus after 20–24 h of incubation at
30–32
C have a dry, flat, ‘ground-glass’ appearance,
translucent to creamy white with a violet–red back-
ground surrounded by a readily visible zone of egg-
yolk precipitate (Figure 1). Less commonly, colonies
may be large (5 to over 10 mm in diameter), amorph-
ous, spreading, and highly irregular. MYP remains to
date a popular choice for the isolation of B. cereus
and is available commercially. Plates can be stored for
up to 7 days at 4
C.
0010KG agar In 1971, Kim and Goepfert developed the
Kim–Goepfert (KG) medium with a similar sensitivity
and selectivity to MYP. It contains no carbohydrates
and low levels of peptone to promote free spore for-
mation within 24 h of incubation at 37
C. Surface
colonies of B. cereus after overnight incubation at
37
C are flat, dry, and of a rough appearance, similar
to the appearance of the colonies on MYP agar. The
composition of KG agar allows direct confirmation
of lecithinase-producing organisms by microscopic
examination, differentiation of B. cereus from
B. thuringiensis based on production of parasporal
crystals of d-endotoxin by the latter species during
sporogenesis, and the use of fluorescent-labeled anti-
bodies to spore-coat antigens as a rapid and specific
serological test for confirmation of identity. Bacillus
spp. that may produce lecithinase, such as B. poly-
myxa and B. laterosporus, are unable to do so under
the nutritionally poor KG medium.
0011PEMBA medium Polymyxin–pyruvate–egg yolk–
mannitol–bromothymol blue agar (PEMBA), de-
veloped by Holbrook and Anderson in the early
1980s, also relies on the mannitol-negative, lecithi-
nase-positive properties of the organism. The medium
contains pyruvate to reduce the tendency of B. cereus
to form rhizoid colonies, improve the egg yolk reac-
tion, and enhance sporulation. After 18–24 h of incu-
bation at 37
C, B. cereus strains form flat, crenate to
rhizoid, turquoise–peacock blue colonies, 2–5mm in
diameter, with a ‘ground-glass’ surface appearance.
After a further 24 h of incubation at ambient tem-
perature, all colonies turn peacock blue. B. cereus is
differentiated microscopically from other mannitol-
negative species by microscopic examination of the
stained cells for the presence of lipid granules and
spores in the cytoplasm.
0012PEMPA medium Polymyxin–Pyruvate–egg yolk–
mannitol–bromocresol purple agar (PEMPA), a
modified agar, gives comparable results to PEMBA
after shorter incubation times (18–24 h).
tbl0001 Table 1 Characteristics of Bacillus cereus and other members
of the B. cereus group
Reaction B.
cereus
B.
mycoides
B.
thuringiensis
B.
anthracis
Egg-yolk reaction þ
a
þþ (þ)
b
Acid from mannitol
Catalase þþ þ þ
Gram reaction þþ þ þ
Motility +
c
d
+
Hemolysis
(sheep RBC)
þ (þ)(þ)
Rhizoid growth þ
Production of
toxin crystals
þ
Anaerobic
utilization
of glucose
þþ þ þ
Nitrate reduction þþ þ þ
VP reaction þþ þ þ
Tyrosine
utilization
þ (þ) þ (þ)
Resistance
to lysozyme
þþ þ þ
a
þ, 90–100% of strains are positive.
b
(þ), usually weakly positive.
c
+, 50–90% of strains are positive.
d
, <10% of strains are positive.
360
BACILLUS
/Detection
0013 Blood agar On 5% horse blood agar, after 24 h at
35–37
C, B. cereus colonies are 3–8 mm in diameter,
flat to low convex, gray–white to gray–green, with a
matt ‘ground glass’ surface. Growth is also associated
with a strong ‘mouse-like’ odor. Hemolytic reactions
on the medium vary from partial lysis of erythrocytes
to complete b-hemolysis. Small colonies, 2–3mm in
diameter, form during anaerobic incubation. The ir-
regular, b-hemolytic colonies resemble those of Clos-
tridium perfringens and require further identification.
0014 Of the plating media, MYP agar is the official
medium in the USA (Association of Official Analyt-
ical Chemists), Canada, France, and several other
countries. PEMBA is widely used in the UK. These
plating media do not differentiate between B. cereus
and B. thuringiensis organisms (Table 1). In general,
the numbers of B. cereus recovered from foods are
sufficiently high for direct plating, and preenrichment
is unnecessary. However, preenrichment may be re-
quired when low viable counts or damaged cells are
present. The most probable number (MPN) tech-
nique, using trypticase soy-polymyxin B broth and
incubation for 48 h at 30
C, is recommended for
estimation of B. cereus numbers in foods expected to
contain less than 10 cells per gram. Tubes showing
growth are streaked on to MYP agar. On egg-yolk-
containing agar plates, zones of egg-yolk precipita-
tion from individual colonies tend to coalesce if high
levels of the organisms are present (Figure 1), and
enumeration of colonies may become difficult. Fur-
ther dilutions of samples to three to 30 colonies are
therefore recommended in quantitative analyses.
Chromogenic Agar Media
0015 Chromogenic substrates that utilize specific enzym-
atic activities are increasingly used for rapid detection
and identification of microorganisms. Their incorpor-
ation into selective agar media facilitates identifica-
tion of colonies, improves the accuracy of the test and
reduces the need for isolation of pure cultures and
confirmation. The enzyme inositol-specific phospho-
lipase C (PI-PLC) was first reported in culture filtrates
of B. cereus in 1965, and a similar enzyme was later
identified in B. thuringiensis. On a chromogenic, se-
lective, and differential plating medium that detects
the enzyme phosphatidylinositol-specific phospholi-
pase C (PI-PLC) in B. cereus and B. thuringiensis
(BCM
1
Bacillus cereus and Bacillus thuringiensis,
Biosynth, Staad, Switzerland), these organisms form
large, 2–7-mm, turquoise, flat colonies with or with-
out turquoise halos on the medium. Other Bacillus
spp. form white, 1–2-mm colonies or are inhibited;
some Listeria strains form colonies less than 1 mm in
diameter, white or turquoise on the medium. Com-
parison with MYP agar has shown a significantly
higher inhibition of Gram-positive non-B. cereus/B.
thuringiensis organisms that interfere with enumer-
ation of small numbers of B. cereus in heavily con-
taminated samples. Typical colonies of B. cereus on
the chromogenic agar plate are shown in Figure 1.
The shelf stability of the chromogenic agar plates (3
months at 2–5
C) is superior to that of MYP plates.
As with other procedures, the chromogenic medium
does not differentiate between B. cereus and B. thur-
ingiensis organisms.
Confirmatory Procedures
0016The specific confirmation tests that distinguish
B. cereus from the other members of the B. cereus
group include motility, toxin crystal formation, rhi-
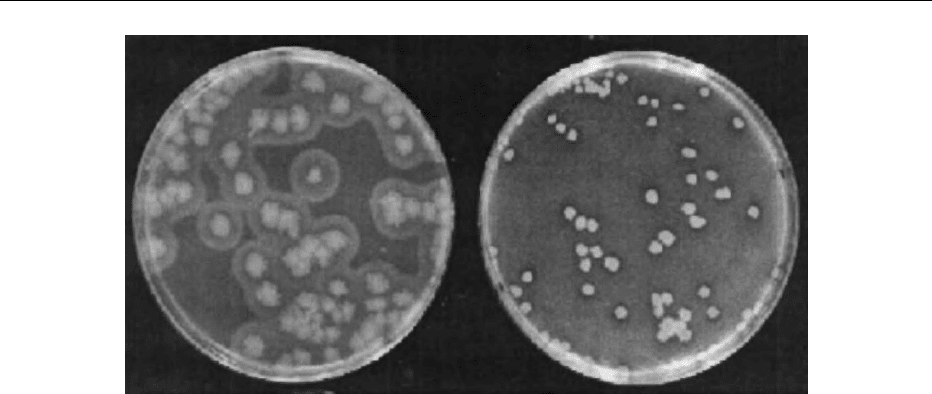
zoid growth, and hemolysin activity (Table 1).
fig0001 Figure 1 Typical colonies of B. ce r e u s on plates of MYP agar (left) and BCM
Õ
agar (right).
BACILLUS
/Detection 361

Motility is determined by stabbing the center of a
tube of semisolid medium and incubating for 18 h at
30
C. The motile organisms (most B. cereus and
B. thuringiensis) diffuse out from the stab, forming
an opaque growth pattern. The only established dif-
ference between B. cereus and B. thuringiensis strains
is the presence of genes encoding for the insecticidal
toxins, usually present on plasmids. The ability of
B. thuringiensis to form toxin crystals is determined
by inoculating nutrient agar slants with the
overnight culture and incubation for 2–3 days at
room temperature. Toxin crystals appear on a smear
of a microscope slide stained with 0.5% basic fuchsin
as dark-staining diamond-shaped objects after their
release from the sporangium upon lysis. The test is
inconclusive if the release is not observed. Rhizoid
growth, characteristic of B. mycoides, is observed by
the production of hair- or root-like structures project-
ing from an agar plate inoculated with the organism
and incubated for 48–72 h. Rhizoid growth may be
lost by the organisms. All three species show strong
hemolytic activity on trypticase soy–sheep blood agar
plates inoculated with an overnight culture and incu-
bated at 35
C for 24 h.
0017 A rapid confirmatory staining technique for the
identification of B. cereus developed by Holbrook
and Anderson uses colonies grown on PEMBA
medium and a combined spore and intracellular
lipid stain. Only the B. cereus group bacilli produce
the intracellular lipid globules.
0018 The US Food and Drug Administration confirms
presumptive B. cereus colonies from MYP agar plates
by anaerobic production of acid from glucose, reduc-
tion of nitrate to nitrite, production of acetylmethyl-
carbinol, decomposition of l-tyrosine, and growth in
the presence of 0.001% (wt/vol) lysozyme. Add-
itional tests for rhizoid growth, hemolysis of sheep
erythrocytes, motility, and production of parasporal
d-endotoxin crystals enable differentiation of
B. mycoides, B. thuringiensis,andB. anthracis. The
Central Public Health Laboratory, London, UK con-
firms presumptive B. cereus by testing motility, hem-
olysis, rhizoid growth, susceptibility to g-phage
(B. cereus-negative; B. anthracis-positive), and fer-
mentation of ammonium salt-based glucose, but not
of mannitol, arabinose, or xylose, after 5 days of
incubation at 36
C.
Serological Tests
0019 The serological relationships within the genus Bacil-
lus are not as well defined as for other genera, and the
development of serological procedures for use as
diagnostic aids has met with limited success. Studies
with spore antigens have shown cross-reactions with
other Bacillus species. The multiplicity of spore sur-
face antigens in a single strain of B. cereus has also
been demonstrated. Some of the problems in the use
of spore antigens stem from contamination of their
preparations with vegetative cell debris and forma-
tion of antivegetative cell immunoglobulins, germin-
ation of spores during serum production, which also
results in the formation of antivegetative cell im-
munoglobulins, and in-vitro spore germination
during immunoassay.
0020Forty-two flagellar (H) and 13 somatic (O) anti-
gens have been identified. A serological typing
scheme for B. cereus strains based on flagellar anti-
gens was developed by Gilbert and coworkers and is
used routinely by the Food Hygiene Laboratory, Cen-
tral Public Health Laboratory, Colindale, London,
UK for investigations of food-poisoning outbreaks
and for serotyping strains from nongastrointestinal
infections. It is composed of 42 H antigens raised
against prototype strains of diverse origin. Twenty-
three of the 42 H serotypes are linked to illness, and
serotype H1 is dominant and contains both emetic
and diarrheal strains. A system similar to that used in
the UK was also developed by the Tokyo Metropol-
itan Research laboratory of Public Health in Japan.
0021B. thuringiensis strains have been classified on the
basis of their flagellar antigens. A serological typing
scheme for this species has been developed, and 14
serotypes have been recognized. There is evidence
that certain strains of B. cereus and B. thuringiensis
share common antigens.
Rapid Identification Methods
0022The term rapid methods refers to a variety of tests,
which can shorten the analysis time and reduce labor
(i.e., by automation). Some eliminate the need for
confirmation, whereas others require enrichment
prior to testing, which limits the assay speed but
provides more reliable identification of the target
organism. All require pure culture isolates of the bac-
teria. Rapid methods applicable to B. cereus are dis-
cussed below.
0023Miniaturized biochemical kits to identify Bacillus
species have been developed to produce standardized
test media that improve reproducibility, have a long
shelf-life, are rapid and convenient to use, economize
the use of media, and are inexpensive. The API 20E
and 50 CHB strips (bioMe
´
rieux, Marcy l’Etoile,
France; Hazelwood, MO, USA), are ready-to-use
microtube systems that contain dehydrated media
for standard biochemical tests. The API 50 CHB for
the identification of the genus Bacillus tests the fer-
mentation of 49 carbohydrates, and the phenotypic
profile obtained from these tests is used to identify the
362
BACILLUS
/Detection

strain. The API 20E strips are used for the identifica-
tion in the Vitek automated identification system,
which eliminates the need for manual identification.
Incubation for 24–48hat30or37
C (mesophiles) is
required before reading the results. Other manual and
automated systems for identification of Bacillus
based on biochemical properties are available. Micro-
Log
TM
microbial carbon-source utilization (Biolog
Inc., Hayward, CA) generates an identity profile
based on the ability of the organism to oxidize
different carbon sources based on changes in the
redox potential following respiration of the viable
bacterial cells. Oxidation is determined colorimetri-
cally with a tetrazolium-based compound, and the
color change data measured automatically by a spec-
trophotometer are fed into a computer. Bacteria are
identified by comparison with a reference data base.
The BBL Crystal
TM
Gram-positive Identification
System (Beckton Dickinson, Sparks, MD) is another
example of miniaturized identification method for
Gram-positive aerobes employing modified conven-
tional, fluorogenic, and chromogenic substrates.
0024 Another approach for the identification of the or-
ganism is based on its fatty acid profile. Analysis of
the cellular fatty acid composition is carried out by
gas chromatography following extraction of pure cul-
tures and injection into a gas chromatograph (Micro-
bial Identification System, MIS, Microbial-ID,
Newark, DE). The cellular fatty acid profile is ana-
lyzed by computer and compared with database
values to provide the best-match identification of
the organism.
0025 Other diagnostic schemes include polyacrylamide
gel electrophoresis analysis, pyrolysis mass spectrom-
etry, and Fourier-transform infrared spectroscopy.
Classification and identification based on differences
in rRNA are available commercially for food patho-
gens including B. cereus. Ribotyping (RiboPrinter
1
Microbial Characterization System, DuPont Quali-
con, Wilmington, DE) generates precise fingerprints
that can be used to classify isolates at the species level
and trace them to the contaminated source. Steps of
the identification, from cell lysis to data capture and
database comparisons, are automated. All of the
above identification methods require large updated
databases. DNA sequence data for the design of spe-
cific primers for B. cereus are being investigated. The
design of primers within the coupled sequences for
phospholipase C and sphingomyelinase of B. cereus
has been reported, and PCR tests with these primers
are positive only with isolates of the B. cereus group.
Moreover, positive results have also been observed
with an egg-yolk-negative B. cereus isolate, suggest-
ing that the primers could be specific for atypical
B. cereus.
0026Phage typing of B. cereus strains is another epi-
demiological tool in outbreaks of B. cereus food
poisoning. Phages are used in routine test dilution,
and drops are deposited on bacterial lawns on nutri-
ent agar plates that are incubated overnight at 32
C
and examined for lysis. A typing scheme using a set
of 12 phages selected on the basis of host range,
stability, and reproducibility of reaction showed that
most isolates of the pathogen were typeable. A good
correlation (80–100%) was observed between phage
types of strains isolated from specimens of suspected
foods and from symptomatic patients. Most B. thur-
ingiensis strains were also typeable by the same set of
phages.
Detection of
B. cereus
Toxins
0027In-vivo and in-vitro methods for the detection of
B. cereus diarrheal and emetic toxins are summarized
in Table 2. In-vivo methods for the detection of
B. cereus diarrheal enterotoxins include the rabbit
or guinea-pig ileal loop test, dermonecrotic test in
guinea-pig, mouse lethality test, vascular permeabil-
ity test, and Rhesus monkey feeding trials. In the
ileal loop assay, the lower portion of the intestine of
the animal is surgically exposed to allow ligation,
usually of the ileal regions, thereby testing different
samples in a single animal. The sample is injected
into a ligated section, and fluid accumulation is
monitored with time and compared with control
sections. Animal feeding trials (Rhesus monkey and
Suncus marinus) and lethality tests (mouse and
Suncus marinus) are used to detect the presence of
the emetic toxin. Animals (a set of six) are observed
for about 5 h after samples have been introduced
tbl0002Table 2 Detection methods of B. c ereus toxins
a
In-vivo methods In-vitro methods
Diarrheal syndrome
Rabbit or guinea-pig
ligated ileal loop
Gel-diffusion assays
Guinea-pig
dermonecrotic test
Cell cytotoxicity
(hemolysin BL)
Mouse lethality test
BCET-RPLA kit (L
2
of BL)
Vascular permeability test
BDE-VIA kit
(nonhemolytic toxin)
Rhesus monkey feeding trials
Emetic syndrome
Feeding trials
(Rhesus monkey and
Suncus marinus)
Cytotoxicity (proliferation of
HEp-2 cell line)
Lethality tests (mouse and
Suncus marinus)
a
See text for details.
BACILLUS
/Detection 363

using a stomach tube. An emetic response in two
of the six animals is considered positive for the
toxin.
0028 The disadvantages of bioassays are the time to
complete the assay, the need for special animal facil-
ities, the cost, as well as the concern about the use
of experimental animals in research. Alternatives to
the in-vivo tests are in-vitro assay methods that
include diffusion techniques, enzyme-linked immu-
nosorbent assays (ELISA), and cell cytotoxicity
tests. The gel diffusion assay detects hemolysin BL,
producing a discontinuous hemolysis pattern on
blood agar plates. The microslide immunodiffusion
assay and the fluorescent immunodot assay detect
enterotoxin by line of identity and fluorescence,
respectively.
0029 Two immunoassay kits are available commercially
for the detection of the B. cereus diarrheal toxins.
These include the B. cereus enterotoxin-reversed pas-
sive latex agglutination (BCET-RPLA) assay (Oxoid,
Unipath, Basingstoke, UK), and the Bacillus diarrheal
enterotoxin visual immunoassay (BDE-VIA) (TECRA
Diagnostics, Batley, UK). The RPLA procedure uses
latex particles to amplify the antibody: antigen reac-
tion. The antibody detects the L
2
component of
hemolysin BL, and can provide a semiquantitative
measure of the enterotoxin in foods. In the TECRA
immunoassay, a sandwich ELISA is used, in which
the antibody is absorbed on the solid phase, and the
enterotoxin is added. The colorimetric reaction
detects the 45-kDa and 105-kDa proteins of the non-
hemolytic enterotoxin and other proteins. The pres-
ence of preformed toxin is detected in 4 h, and
production of the toxin in samples containing
enterotoxigenic Bacillus spp. is detected after over-
night enrichment. Since these two kits detect compon-
ents in the different enterotoxins, isolates should
be tested by both methods. Many B. cereus strains
have been shown to react with both the Oxoid and
the TECRA detection kits, suggesting that they are
able to produce both enterotoxin complexes.
0030 B. thuringiensis, B. subtilis, B. licheniformis,and
B. pumilus strains have been involved in foodborne
illnesses. Although the nature of their toxins is not
well defined, reaction with antibodies to B. cereus
enterotoxins in both the Oxoid and the TECRA de-
tection kits has been demonstrated for some of them.
0031 Cell cytotoxicity techniques have been used for
both the diarrheal toxin and the emetic toxin. Cell
lines used include HeLa, HEp-2, Vero, and others,
and cellular responses in the presence of the toxin
range from morphological changes (subjective), to
more specific, e.g., metabolic status of the cells and
detection of lactate dehydrogenase release from dam-
aged cells.
Taxonomic Relationship of
B. cereus
,
B. thuringiensis
, and
B. anthracis
0032The lower end of the GC range of the genus Bacillus
(32–38%) is occupied by B. cereus and the closely
related species B. anthracis, B. mycoides, and B. thur-
ingiensis. It was suggested in 1952 that there are no
consistent phenotypic properties that differentiate
these species and that these three species be desig-
nated varieties of B. cereus. However, this has not
been accepted for B. anthracis and B. thuringiensis
because of their pathogenic qualities. The high hom-
ology between DNA from B. anthracis, B. thuringien-
sis, and B. cereus suggests that organisms in these
taxa should have the same name. Recently, research-
ers used multilocus enzyme electrophoresis and se-
quence analysis of nine chromosomal genes to
provide further evidence that B. anthracis should
be considered a lineage of B. cereus. Evidence of
the close taxonomic relationship of B. cereus, B. thur-
ingiensis,andB. anthracis was indirectly obtained
from serological studies that showed extensive
cross-agglutination between the spores of the three
species.
0033The only established difference between B. cereus
and B. thuringiensis strains is the presence of genes
encoding for the insecticidal toxins, usually present
on plasmids. None of the detection methods discussed
above distinguishes between the two species.
See also: Bacillus: Occurrence; Food Poisoning
Further Reading
Ahmed R, Sankar-Mistry P, Jackson S, Ackerman H-W and
Kasatiya SS (1995) Bacillus cereus phage typing as an
epidemiological tool in outbreaks of food poisoning.
Journal of Clinical Microbiology 33: 636–640.
Berkeley RCW and Goodfellow M (1981) The Aerobic
Endospore-Forming Bacteria: Classification and Identi-
fication. London: Academic Press, Society for General
Microbiology.
Claus D and Berkeley RCW (1986) Genus Bacillus Cohn
1872, 174. In: Sneath PHA, Mair NS, Sharpe ME and
Holt JG (eds) Bergey’s Manual of Systemic Bacteriology,
vol. 2, pp. 1105–1139. Baltimore, MD: Williams &
Wilkins.
Granum PE and Lund T (1997) Mini review: Bacillus cereus
and its food poisoning toxins. FEMS Microbiology
Letters 157: 223–228.
International Commission on Microbiological Specifica-
tions for Foods (1996) Bacillus cereus. In: Microorgan-
isms in Foods 5. Characteristics of Microbial pathogens,
pp. 20–35. London: Blackie Academic & Professional.
Kramer JM and Gilbert RJ (1989) Bacillus cereus and other
Bacillus species. In: Doyle MP (ed.) Foodborne Bacterial
Pathogens, pp. 21–70. New York: Marcel Dekker.
364
BACILLUS
/Detection