Белясова Н.А. Биохимия и молекулярная биология
Подождите немного. Документ загружается.


230
ние хоризмата, которое происходит от греческого слова, означающего «вил-
ка»).
Фенилаланин образуется в ходе трех последовательных реакций: изоме-
ризации хоризмата в префенат, дегидратирования и декарбоксилирования
префената в фенилпируват и трансаминирования фенилпирувата с участием
глутамата.
Заменимая кислота тирозин может синтезироваться из фенилаланина пу-
тем его гидроксилирования с участием фенилаланин-4-моноокси-геназы, а
также из префеновой кислоты после ее декарбоксилирования и аминирова-
ния.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
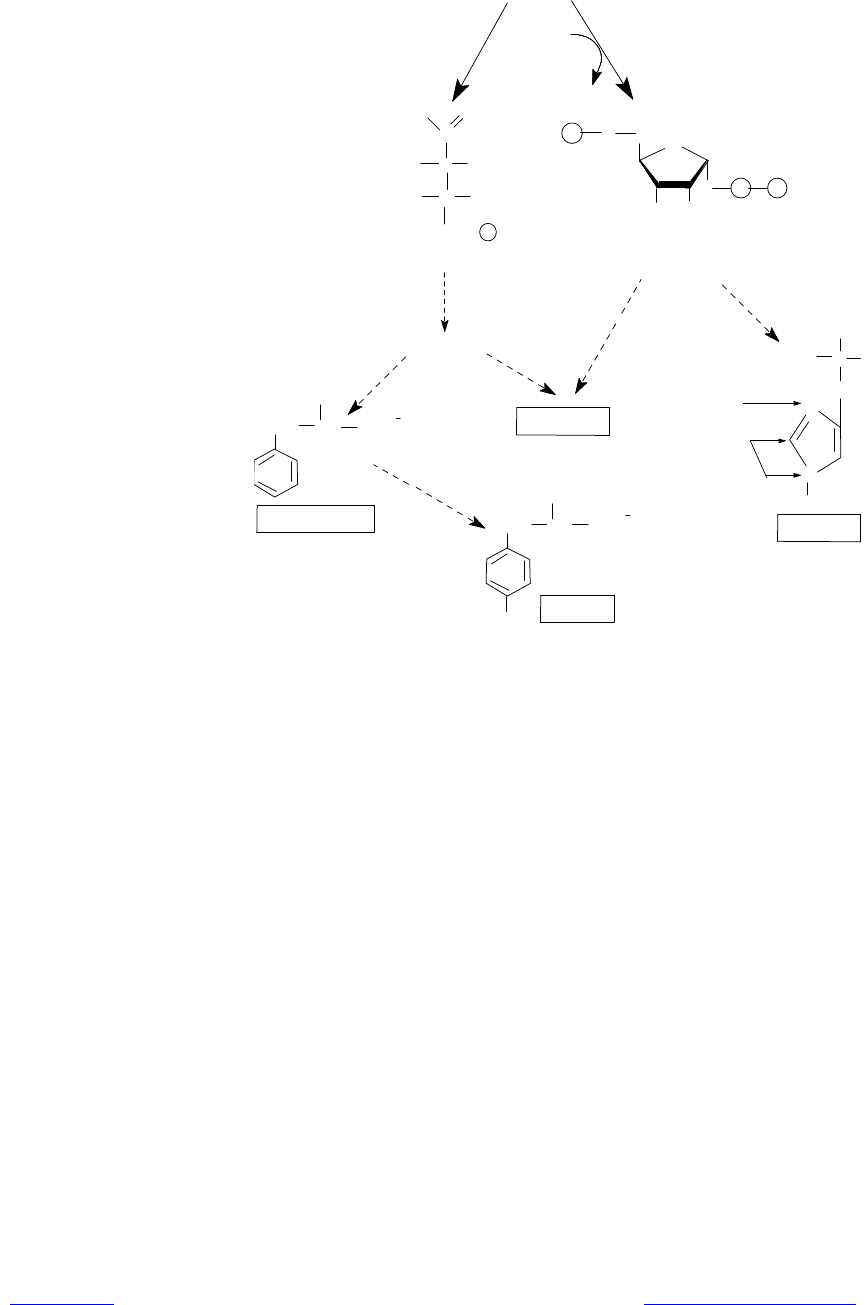
231
Изомеризация, меж
молекулярные пере
стройки в пентозо
фосфатных путях
АТР
АМР
Рибозофосфат
пирофосфокиназа
O
CH
2
O
O
P
P P
OH
OH
O
H
H OH
C
C
C
CH
2
O P
OHH
Гистидин
Хоризмат
Фенилаланин
Тирозин
Триптофан
COO
-
C
CH
2
N
N
H
H
3
N
+
H
из глу
тамина
из АТР
CH
2
NH
3
CH
+
+
COOCH
NH
3
CH
2
OH
Р
и
б
о
з
о
-
5
-
ф
о
с
ф
а
т
Эритрозо-4-фосфат
5-Фосфорибозил-
-1-пирофосфат
+ Фосфоенол-
пируват
COO
-
-
-
-
-
Рис. 16.10. Общая схема биосинтеза гистидина, триптофана, фенилаланина и
тирозина. Пунктирные стрелки обозначают многоэтапные процессы
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
232
H OH
CH
2
O P
C
C
C
OHH
H
O
+
COO
-
C
CH
2
O
P
Фосфоенолпируват
O
COO
-
C
C
CH
2
H
HO
H OH
C
C
CH
2
O P
OHH
OH
OH
HO
COO
-
Шикимовая кислота
COO
-
HO
O
C
CH
2
Хоризмовая кислота
Антранилат
синтаза
COO
-
NH
3
+
Антранилат
Пиру
ват
Фосфорибозил
антранилат
трансфераза
PP
i
OH
OH
P O
CH
2
O
HN
OOC
Фосфорибозил
антранилат
изомераза
NH
COO
-
CH C CH CH CH
2
O P
OH OH OH
N
CH
OHOH
CH
2
CH
H
H
2
O
CO
2
H
COO
-
CHCH
2
N
NH
3
+
Триптофан
Триптофансинтаза
Фосфорибо
зилпиро
фосфат
Серин
O P
Эритрозо-4-фосфат
7-Фосфо-2-кето-
-3-дезокси-D-ара-
биногептулозонат-
синтаза
7-Фосфо-2-кето-
-3-дезокси-D-ара-
биногептулозонат
5-Фосфорибозил-
антранилат
1(О-карбоксифениламино)-1-
дезоксирибулозо-5-фосфат
Индол-3-глицерофосфат
Индол-3-глицерофосфат-
синтаза
Глицеральде-
гид-3-фосфат
COO
-
-
-
-
-
-
-
-
-
-
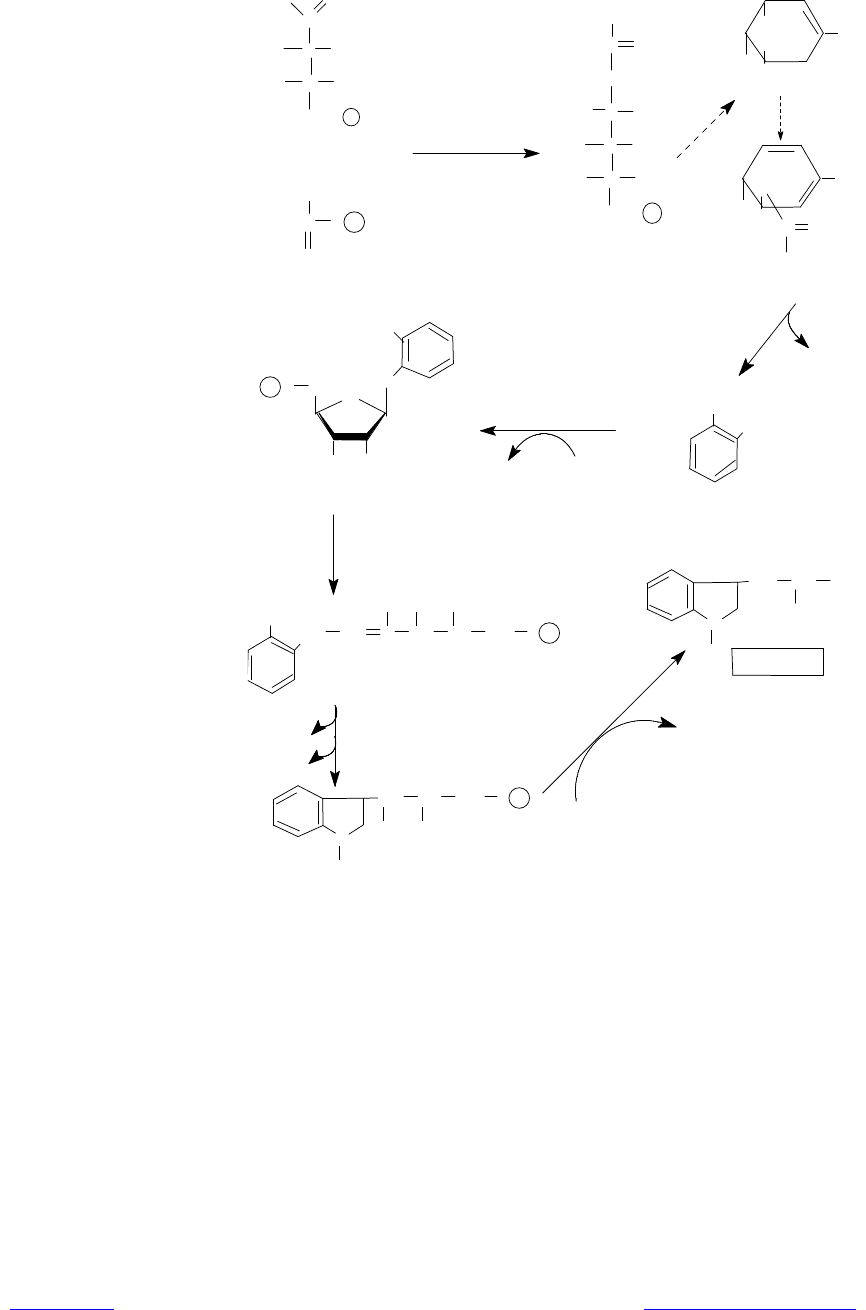
Рис. 16.11. Путь биосинтеза триптофана. Пунктирные стрелки обозна-
чают многоэтапные процессы
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

233
Закономерности биосинтеза аминокислот. Обзор путей биосинтеза
протеиногенных аминокислот позволяет выявить следующие основные зако-
номерности этих процессов: 1) углеродные скелеты аминокислот берут свое
начало от промежуточных продуктов гликолиза (3-фосфоглицерат, фосфое-
нолпируват, пируват), пентозофосфатных путей (рибозо-5-фосфат и эритрозо-
4-фосфат), ЦТК (оксалоацетат и a-кетоглутарат); 2) донором аминогрупп для
большинства протеиногенных аминокислот служит глутамат, реже —
глутамин; реакции, в которых аминогруппа аминокислоты переносится на
кетокислоту, называются «реакции трансаминирования»; 3) биосинтез мно-
гих аминокислот осуществляется «семействами», для которых используются
общие предшественники; многие аминокислоты сами служат субстратами для
синтеза других аминокислот; 4) многие стадии биосинтеза аминокислот тре-
буют притока энергии и сопровождаются гидролизом АТР (стадии синтеза
гистидина, пролина, метионина, аспарагина, глутамина, аргинина); кроме
этого, используется энергия активированных молекул, участвующих в синте-
зе; наконец, из катаболических и амфиболических процессов изымаются
промежуточные продукты, которые могли бы обеспечить клетке запасание
энергии; 5) многие этапы биосинтеза аминокислот требуют участия восстано-
вительных эквивалентов (NADH и NADPH), которые могли бы быть окисле-
ны в дыхательной цепи и обусловить энергетический выигрыш.
Таким образом, биосинтез аминокислот обходится клетке достаточно до-
рого. Неудивительно поэтому, что данный процесс в каждом организме
(клетке) подвержен очень сложной регуляции (глава 19), которая, с одной
стороны, определяется сложностью и разветвленностью самого биосинтеза
протеиногенных аминокислот, а с другой стороны, должна обеспечить стро-
гую экономию клеточных ресурсов (энергии, восстановительных эквивален-
тов, строительных блоков). Закономерным представляется и тот факт, что при
наличии экзогенных аминокислот, клетки микроорганизмов, в частности, не
осуществляют их синтез самостоятельно, а используют готовые формы.
16.5. Расщепление аминокислот
В первом разделе данной главы уже охарактеризована необходимость и
основная стратегия расщепления аминокислот. Она объясняется невозможно-
стью запасания аминокислот впрок и невозможностью их выведения из кле-
ток целиком. Избыточные аминокислоты используются организмами как ме-
таболическое топливо: их углеродные скелеты при перестройках определен-
ного рода могут вовлекаться в биосинтез жирных кислот, глюкозы, кетоно-
вых тел, изопреноидов и др., а также окисляться в ЦТК, обеспечивая клетку
энергией. Следует отметить, что многие микроорганизмы, в частности аэроб-
ные бактерии, способны использовать отдельные аминокислоты в качестве
единственного источника энергии и углерода. У анаэробных микроорганиз-
мов, при отсутствии в клетках цикла трикарбоновых кислот, выработался
другой механизм: катаболизм аминокислот в парах, когда одна из них служит
донором электронов, а вторая — акцептором. Важно, что в таком процессе
происходит образование АТР.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

234
Кроме углеродных скелетов, при деградации аминокислот образуется
аминный азот, который в отличие от углерода не пригоден для получения
энергии за счет окисления, и более того, является токсичным для клеток. По-
этому те аминогруппы, которые не могут повторно использоваться в биосин-
тезе, превращаются в мочевину (или другие вещества) и выводятся из орга-
низма.
Ниже будут рассмотрены основные типы реакций, в которые могут всту-
пать аминокислоты: реакции по a-аминогруппе, карбоксильной группе и бо-
ковой цепи.
Расщепление аминокислот по аминогруппе. Эти процессы представле-
ны в основном реакциями трансаминирования и дезаминирования по a-
аминогруппе. Реакции трансаминирования уже были рассмотрены в разделе,
касающемся биосинтеза аминокислот. Они катализируются трансаминазами
(аминотрансферазами), отличительной особенностью которых является ис-
пользование пиридоксальфосфата (производное витамина В
6
) в качестве про-
стетической группы. Наибольшее значение в процессах деградации амино-
кислот имеют глутамат-трансаминаза и аланин-трансаминаза. Эти ферменты
выполняют роль «воронок», собирающих аминогруппы от разных аминокис-
лот и включающих их в состав глутамата и аланина. У животных эти две
аминокислоты служат переносчиками накапливающегося аминного азота из
тканей в печень. В печени аминогруппа аланина переносится аланин-
трансаминазой на a-кетоглутарат с образованием глутамата:
Аланин + a-Кетоглутарат ® Пируват + Глутамат
Таким образом, большинство аминогрупп различных аминокислот оказы-
вается в составе глутамата, который легко подвергается дезаминированию.
Реакции дезаминирования аминокислот приводят к освобождению NH
2
-
группы в виде аммиака и осуществляются тремя разными путями. Различают
окислительное, гидролитическое и прямое дезаминирование (рис. 16.12).
Наиболее распространенным типом является окислительное дезаминирова-
ние, которое осуществляется по a-аминогруппе и катализируется в основном
глутаматдегидрогеназой — типичным для печени ферментом. Необычным
свойством этого фермента является способность использовать как NAD, так и
NADP в качестве коферментов. Активность глутаматдегидрогеназы регули-
руется аллостерическими активаторами (ADP, GDP) и ингибиторами (ATP,
GTP).
Окислительное дезаминирование осуществляется в две стадии с образова-
нием иминокислоты в качестве промежуточного продукта, который спонтан-
но гидролизуется, превращаясь в кетокислоту и аммиак (рис. 16.12). Обе ре-
акции обратимы, и их константы равновесия близки к единице. Ранее
(рис. 16.3) было показано, как в ходе обратной реакции аммиак включается в
состав глутамата. Можно считать, что реакция образования и дезаминирова-
ния глутамата является центральной реакцией в процессе метаболизма ам-
миака.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

235
COO
-
C
R
1
H
3
N H
+
NAD
+
NADH
Глутамат
дегидрогеназа
+
HH
2
N
R
1
COO
-
C
Аминокислота
Иминокислота
(NADP
+
)
(NADPH)
H
2
O
NH
4
+
O
R
1
COO
-
C
Кетокислота
+
HH
3
N
CH
2
COO
-
C
Глутаминаза, аспарагиназа
H
2
O
NH
4
+
COO
-
C
H
Аминокислота
+
HH
3
N
R
3
COO
-
C
CH
2
CH
COO
-
CH
R
3
NH
4
+
Гистидин аммиак
лиаза
Непредельная кислота
CONH
2
( )
n
( )
n
COOH
CH
2
Глутамин,
аспарагин
H
3
N
+
Глутамат,
аспартат
1. Окислительное дезаминирование
2. Гидролитическое дезаминирование
3. Прямое (внутримолекулярное) дезаминирование
-
-
-
Рис. 16.12. Различные типы реакций аминокислот по аминогруппе
У многих организмов окислительное дезаминирование осуществляется с
помощью дегидрогеназ, использующих флавиновые кофакторы (FMN, FAD).
Эти ферменты называют оксидазами аминокислот. Они характеризуются ши-
рокой субстратной специфичностью: одни специфичны к L-аминокислотам,
другие — к их D-аналогам. Считается, что эти ферменты вносят небольшой
вклад в обмен аминогрупп.
Гидролитическому дезаминированию подвержены немногие амино-
кислоты, из протеиногенных — аспарагин и глутамин. При их дезаминирова-
нии образуются соответственно аспартат и глутамат. Этот процесс правиль-
нее называть дезамидированием, поскольку он осуществляется за счет амид-
ной группы (рис. 16.12). В редких случаях таким путем отщепляется и a-
аминогруппа аминокислоты, тогда образуются аммиак и оксикислота.
В результате прямого (внутримолекулярного) дезаминирования воз-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

236
никают ненасыщенные соединения. Прямому дезаминированию обычно под-
вергается гистидин, а также серин. Однако первичная ферментативная атака
серина приводит к отщеплению молекулы воды (фермент —серингидратаза),
и в этом превращении участвует боковая гидроксильная группа серина. Спон-
танному дезаминированию в данном случае подвергается нестабильное про-
межуточное соединение — аминоакрилат. Продуктом суммарной реакции
является пируват, и этот тип дезаминирования вызывается перестройкой в
боковой цепи аминокислоты.
Реакции аминокислот по карбоксильной группе. Превращения по кар-
боксильной группе аминокислот могут использоваться организмами для де-
градации этих молекул, а также для превращения в другие, необходимые
клетке соединения, в первую очередь аминоациладенилаты и биогенные
амины. Образование аминоациладенилатов на подготовительной стадии син-
теза белка уже было описано в главе 3. Биогенные амины возникают в реак-
циях, катализируемых декарбоксилазами аминокислот. Эти ферменты широ-
ко распространены у животных, растений и особенно у микроорганизмов,
причем известно, что у патогенных микроорганизмов декарбоксилазы могут
служить факторами агрессии, с помощью которых возбудитель проникает в
соответствующие ткани. Декарбоксилазы L-аминокислот, так же как транса-
миназы, используют в качестве простетической группы пиридоксальфосфат.
Моноамины (биогенные амины) выполняют в организмах разнообразные
функции. Например, этаноламин, образующийся при декарбоксилировании
серина, является составной частью полярных липидов. При декарбоксилиро-
вании цистеина и аспартата образуются соответственно цистеамин и b-
аланин, которые входят в состав такого важного для клеток кофермента, как
коэнзим А. Декарбоксилирование гистидина приводит к образованию гиста-
мина — медиатора, участвующего в регуляции скорости метаболических
процессов, деятельности желез внутренней секреции, регуляции кровяного
давления у животных. Многие другие биогенные амины выполняют функции
сигнальных веществ, в частности широко распространенных у животных и
человека нейромедиаторов.
Реакции аминокислот по боковой цепи. Насколько разнообразна струк-
тура радикалов аминокислот, настолько разнообразны и химические превра-
щения, которым они могут подвергаться. Среди этих многообразных реакций
можно выделить те, которые позволяют клетке получать из одних аминокис-
лот другие. Например, тирозин образуется при окислении ароматического
кольца фенилаланина; гидролиз аргинина приводит к формированию орни-
тина (см. цикл мочевины); расщепление треонина сопровождается образова-
нием глицина и т. п.
Кроме этих реакций, важное значение имеют превращения боковых
групп, связанные с возникновением физиологически активных веществ. Так,
из тирозина образуется гормон адреналин, из триптофана образуются нико-
тиновая кислота (витамин РР, входящий в состав никотинамидных кофер-
ментов) и индолилуксусная кислота (ростовое вещество), из цистеина —
меркаптуровые кислоты (участвуют в обезвреживании ароматических соеди-
нений). Уже отмечалась возможность превращения серина в пируват при де-
гидратации его боковой цепи и дезаминировании.
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

237
Таким образом, разнообразные химические превращения аминокислот
могут приводить к образованию биологически активных веществ с широким
спектром действия и, кроме того, к отщеплению аминогрупп в виде аммиака
с формированием углеродных скелетов. В следующем разделе будет рассмот-
рена судьба аммиака и углеродных атомов расщепленных аминокислот.
16.6. Выведение аммиака из организма
При деградации аминокислот, которая у млекопитающих осуществляется
в печени, высвобождается аммиак. Кроме этого, значительные количества
аммиака образуются при распаде пуринов и пиримидинов. Часть аммиака
сразу расходуется на синтез других аминокислот и азотистых оснований, а
оставшаяся часть должна быстро инактивироваться или выводиться из орга-
низма, поскольку это соединение является сильным клеточным ядом.
У организмов, стоящих на низших ступенях развития, а также у обитаю-
щих в воде животных аммиак выделяется непосредственно из клеток, а у рыб,
например, через жабры. Такие животные носят название аммониотеличе-
ские. У большинства наземных позвоночных животных, в том числе у мле-
копитающих и у человека, аммиак превращается в мочевину. Организмы,
удаляющие аммиак (основную его часть) в виде мочевины называются урео-
телические. Наконец, существуют урикотелические организмы (птицы и
рептилии), которые превращают аммиак в мочевую кислоту и выводят ее из
организма в твердом виде.
Свойства мочевины как вещества, в составе которого аммиачный азот
выводится из уреотелических организмов, заключаются в том, что это ней-
тральное, нетоксичное, низкомолекулярное соединение, способное легко (в
ходе пассивной диффузии) преодолевать мембранные барьеры, растворимое в
воде. Мочевина легко переносится кровью и выводится с мочой.
Образуется мочевина в клетках печени в замкнутой последовательности
реакций, которые носят название цикл мочевины.
Цикл мочевины. Один из атомов азота в составе молекулы мочевины
происходит из аммиака, второй — из аспартата. Углеродный атом мочевины
берет происхождение из молекулы СО
2
.
На первой стадии свободный аммиак и углекислота образуют в АТР-
зависимой реакции молекулу карбамоилфосфата, которая содержит ангид-
ридную связь и характеризуется высоким потенциалом переноса карбамоиль-
ной группы. Карбамоильный остаток переносится на орнитин с помощью ор-
нитин-карбамоилтрансферазы и образуется цитруллин (рис. 16.13).
На следующей стадии при участии аргининосукцинат-синтазы в цикл
включается вторая аминогруппа, которая присутствует в составе аспартата.
Конденсация цитруллина с аспартатом также сопряжена с гидролизом АТР и
приводит к образованию аргининосукцината.
На последнем этапе с участием аргининосукциназы происходит расщеп-
ление ее субстрата на аргинин и фумарат. Аргинин гидролизуется ар-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)
238
NH
2
C
P
O
Карбамоил
фосфат
COO
-
C
N
CH
2
CH
2
CH
2
H
HH
H
H
3
N
+
Орнитин
+
H
3
N
H
H
C
CH
2
CH
2
CH
2
N
COO
-
C
NH
2
O
COO
-
C
N
CH
2
CH
2
CH
2
C
H
H
H
3
N
+
NH
NH
C
H
Аргининосукцинат
NH
2
NH
+
H
3
N
H
H
C
CH
2
CH
2
CH
2
N
COO
-
C
Аргинин
Фумарат
Н
2
О
O
C
NH
2
Мочевина
Фумарат
Реакции ЦТК
Оксалоацетат
Аспартат
Аспартаттранс
аминаза
Glu
КГ
М и т о х о н д р и я
(м а т р и к с )
Ц и т о з о л ь
Цитруллин
P
i
ATP
1
2
3
4
NH
3
+ CO
2
+ 2ATP
2ADP + P
i
AMP + PP
i
+
Карбамоилфос-
фат-синтаза
H
2
N
-
OOC
COO
-
CH
2
-
-
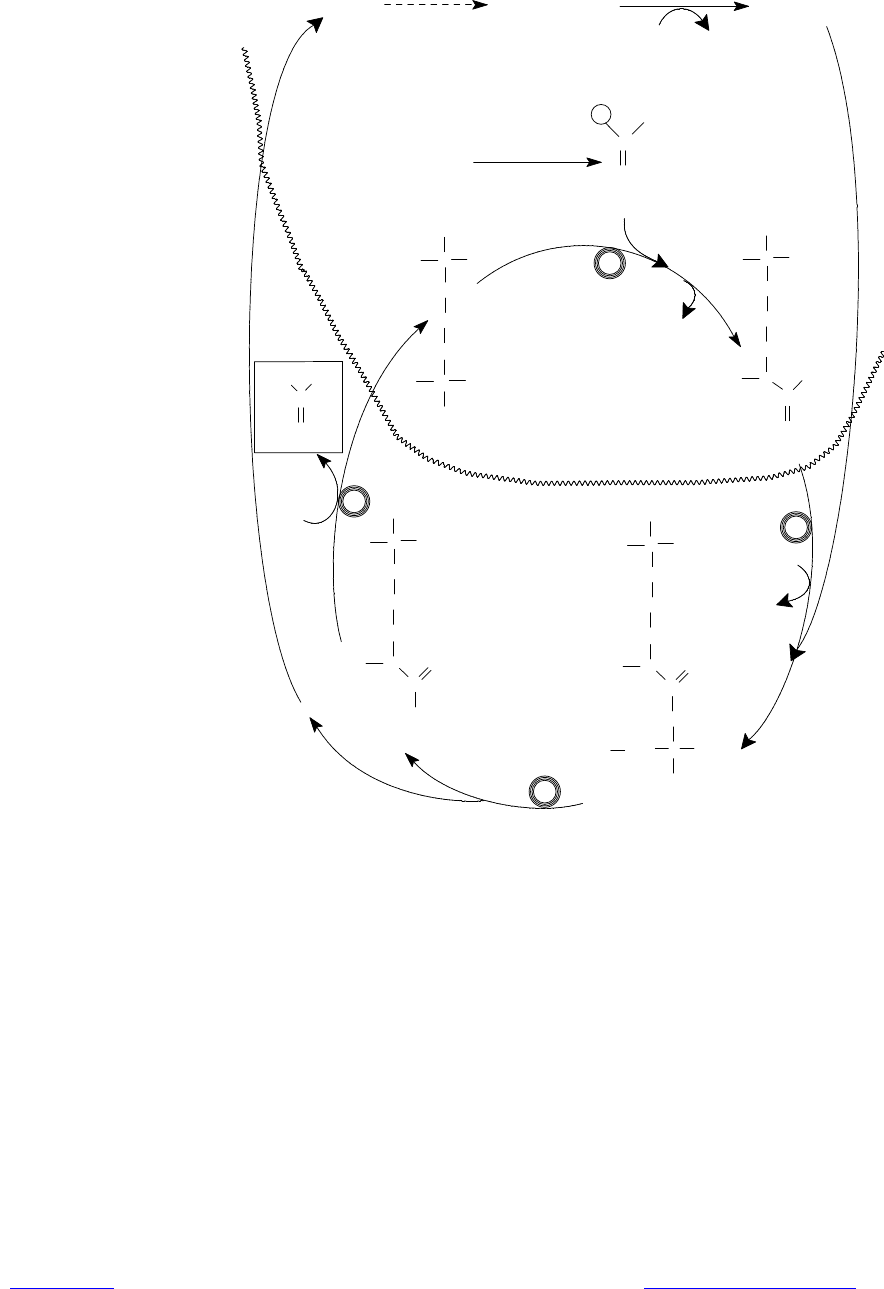
Рис. 16.13. Цикл мочевины. В кружках под стрелками — названия ферментов:
1 — орнитин-карбамоилтрансфераза; 2 — аргининосукцинат-синтаза; 3 —
аргининосукцинат-лиаза; 4 — аргиназа. Обозначения: Glu — глута-мат;
КГ — a-кетоглутарат
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)

239
гиназой на мочевину и орнитин, который вновь может акцептировать карба-
моильный остаток. Фумарат может превращаться в оксалоацетат (реакции
ЦТК), который в ходе трансаминирования с участием глутамата преобразует-
ся в аспартат. Аспартат снова может поступать в цикл мочевины, конденси-
руясь с цитруллином (рис. 16.13).
Компартментализация цикла мочевины охватывает митохондрии и цито-
золь: в матриксе протекают реакции образования карбамоилфосфата, его
включение в состав цитруллина и преобразование фумарата в аспартат. Ос-
тальные три реакции (образование аргининосукцината, аргинина и фумарата,
орнитина и мочевины) осуществляются в цитозоле.
На образование одной молекулы мочевины расходуется четыре макроэр-
гических связи АТР, т. е. процесс инактивации аммиака стоит клетке боль-
ших затрат энергии.
Регуляция скорости цикла мочевины определяется первой реакцией. Ка-
тализирующая ее карбамоилфосфат-синтаза аллостерически активируется N-
ацетилглутаматом, предшественником орнитина, и в его отсутствие практи-
чески неактивна.
Цикл мочевины — основной процесс, в котором происходит обезврежи-
вание аммиака. Однако даже уреотелические организмы способны выделять
небольшую долю аммиака непосредственно при гидролитическом дезамиди-
ровании глутамина и окислительном дезаминировании глутамата в клетках
почек. Высвободившийся аммиак диффундирует через клеточные мембраны
почечных трубочек в почечные канальцы (мочу), где соединяется с протона-
ми, образуя ионы аммония. В такой форме аммиак уже не может преодолеть
мембранный барьер и возвратиться в клетку, поэтому полностью выводится в
составе мочи.
Известны наследственные заболевания, связанные с нарушением цикла
мочевины из-за дефектов в его ферментной системе. Это всегда связано с
увеличением концентрации аммиака в тканях и крови (гиперам-монемия),
что может приводить даже к летальному исходу. При подобных заболеваниях
страдают в первую очередь наиболее чувствительные к снижению содержа-
ния АТР системы — нервные клетки и мозг. Это можно объяснить тем, что
высокая концентрация аммиака сдвигает равновесие обратимой реакции, ка-
тализируемой глутаматдегидрогеназой, в сторону образования глутамата, что
неизбежно вызывает истощение запасов a-кетоглутарата. Но a-кетоглутарат
является промежуточным продуктом ЦТК, и его чрезмерное потребление
приводит к замедлению скорости цикла лимонной кислоты — основного по-
ставщика восстановительных эквивалентов для дыхательной цепи. В резуль-
тате уменьшается скорость образования АТР.
16.7. Судьба углеродных скелетов аминокислот
Стратегия разрушения углеродных скелетов, которые образуются при де-
заминировании аминокислот, состоит в том, чтобы в клетке сформировались
ключевые промежуточные продукты, пригодные как для биосинтеза клеточ-
ных соединений, так и для дальнейшего окисления, связанного с запасанием
энергии. Углеродные скелеты 20 протеиногенных аминокислот в ходе много-
Create PDF files without this message by purchasing novaPDF printer (http://www.novapdf.com)