Титоренко Т.Н. Лекции по курсу стратиграфии
Подождите немного. Документ загружается.

в котором эти виды встречаются совместно. Конечно, эмпирическое определение
комплексов является достаточно субъективным. Поэтому для однозначного понимания
комплексов и, следовательно, для однозначной корреляции разрезов всеми стратиграфами
необходимо, чтобы выделение каждого комплекса сопровождалось его четким диагнозом.
Так, для комплекса, показанного на рис.8., а, характерно: а) совместное существование
видов 3—11, 15—18; б) существование только в рассматриваемом комплексе видов 6—8;
в) исчезновение на нижней границе комплекса видов /—2 и появление видов 5—6, 8—10;
г) исчезновение на верхней границе комплекса видов 4—6, 8 и появление видов 12—14.
Несколько иным будет диагноз комплекса, показанного на рис. 5.64, б, для которого
характерным окажется: а) совместное существование видов 2—8 и 11—14; б)
исчезновение на нижней границе вида / и появление видов 6—8; в) исчезновение на верх-
ней границе видов 3—4 и появление видов 9—10.
Первое преимущество этого метода состоит в том, что каждый органический комплекс
представляет ассоциацию видов, различные группы которых могут иметь
контролирующее значение по отношению друг к другу при определении геологического
возраста. По одной руководящей форме можно сказать о возрасте только ориентировочно.
Определение одной какой-либо формы, выхваченной из комплекса, недостаточно для
стратиграфических выводов. В них нельзя быть уверенными, так как неизвестен
конкретный диапазон вертикального распространения этой формы именно в данном
районе.
Конечно, разные комплексы тоже могут иметь разное значение для выводов о возрасте
вмещающих отложений. И чем больше в комплексе форм, близко подходящих к понятию
руководящих ископаемых, тем ценнее этот комплекс для стратиграфических целей
(рисунок 8).
Наконец, необходимо отметить, что характер изменчивости органического комплекса в
целом будет иметь несравненно большее значение, чем изменчивость отдельных форм.
Допустим, какой-то исследуемый комплекс включает три группы видов, относящихся
даже к разным типам (например, фораминиферы, мшанки, моллюски). При более или
менее резкой смене внешних условий все эти три группы прореагируют на них, но каждая
по-своему: одна, предположим, мало заметно, вторая довольно заметно, а третья резко.
Если строить выводы не то что по отдельным видам, а и по отдельным группам, то в
первом случае характер изменений и наличие какого-то рубежа в развитии данного района
могут быть почти не замечены. Во втором случае этот рубеж бросится в глаза. В третьем
— он будет казаться очень глубоким и значительным.
При исследовании комплекса в целом, зная характер реагирования каждой группы на
конкретные внешние изменения и корректируя общий вывод по каждой из них, можно не
только достоверно зафиксировать на определенном уровне границу между двумя этапами
развития изучаемого района, но и представить себе масштаб происходивших явлений, а
отсюда и ранг наметившегося рубежа. Конечно, важные выводы могут дать комплексы, не
обязательно включающие столь резко различные группы, как в приведенном примере.
В основе метода расчленения и корреляции разрезов по руководящим комплексам
лежит неповторимость каждого органического комплекса, поскольку органический мир
находится в постоянном развитии. В силу этого каждый комплекс не должен быть Похож
на предыдущий, хотя, при относительно небольшом временном интервале между ними,
накопившиеся различия не всегда могут быть выявлены имеющимися в настоящее время
методами.
Неповторимость органических комплексов может быть обусловлена многими
факторами, в первую очередь спецификой филогенетического развития отдельных групп,
путями их распространения на фоне изменяющейся среды в связи с общим ходом
геологического развития района и т. д.
При сопоставлении по руководящим комплексам нужно все время принимать во
внимание фациальную обстановку. В некоторых случаях в разных фациях можно
встретить представителей тех же семейств и родов, но принадлежащих иным видам. Иног-
да разновременные комплексы, но принадлежащие сходным фациальным обстановкам,
могут иметь больше общих черт, чем комплексы из одновозрастных, но разнофациальных
отложений. При наличии в пределах определенной стратиграфической единицы более или
менее заветного числа эврипичных форм хорошо выделить для нее комплекс основных
характерных видов, прослеживая смену органических форм по простиранию в разных
фациях и сравнивая представителей однофациальных отложений, принадлежащих разным
вертикальным уровням.
Т. А. Мордвилко (1953) предлагала выделять в общей палеонтологической
характеристике стратиграфических подразделений возрастные комплексы, или
биокомплексы, понимая под ними определенный комплекс видов, резко отличающийся от
такового в ниже и выше лежащих отложениях качественным или Количественным
составом представителей и точно устанавливающий не только возраст, но и фацию
отложений рассматриваемого подразделения (сам термин «возрастной комплекс» нельзя
признать особенно удачным).
При этом, как один из важных стратиграфических индикаторов, учитывалась массовая
встречаемость представителей определенных видов, отражающая стадию их расцвета. Эта
стадия, с одной стороны, обеспечивает наиболее частую встречаемость представителей
определенного вида в ископаемом состоянии, с другой стороны, вследствие
относительной ограниченности вертикального распространения вида во время расцвета,
по сравнению с его общим вертикальным распространением, обеспечивает четкую
датировку отдельных горизонтов. Таким образом, горизонты с характерными
ракушняками или наиболее часто встречаемыми видами приобретают вполне
определенное стратиграфическое значение.
Помимо транзитных видов, Мордвилко различает виды контролирующие,
приближающиеся по своему значению к руководящим, и маркирующие, наиболее часто
встречаемые. Последние две категории и составляют «возрастной комплекс» (некоторые
виды могут одновременно быть и контролирующими, и маркирующими). Виды, только
появившиеся в данном горизонте, нехарактерные для него и незначительно
представленные, не будут входить в его «возрастной комплекс». Однако расцвет данных
назначение спорово-пыльцевых спектров древних толщ в различного рода
палеогеографических реконструкциях [Гричук В. П., Заклинская Е. Д., 1948].
Действительно, интегральный характер спорово-пыльцевых комплексов, бесспорно,
может дать весьма большую информацию о палеоландшафтах [Палеофитогеография...,
1975].
Иногда появление определенных групп позволяет сделать чрезвычайно важные
выводы о палеоклиматических изменениях.
Например, появление пыльцы Glossopolis в верхнеюрских отложениях на юге Западной
Сибири [Палеозойские и мезозойские флоры..., 1972] позволило установить здесь
аридизацию климата и т. д.
Для целей же стратиграфии более перспективен переход к выявлению видовых
таксонов, изучению их распространения в разрезах и обычной биостратиграфической
интерпретации спорово-пыльцевых комплексов. Несмотря на большие сложности в
выделении видов, сама методика определения образцов станет при этом существенно
менее трудоемкой, а результаты анализов сразу могут быть использованы при конкретных
стратиграфических корреляциях.
Эволюционный метод. Под этим термином понимается корреляция основанная не на
присутствии сравнимых фаунистических комплексов одинаковых форм, а на уровне
эволюционного развития, характеризующего рассматриваемый комплекс Лии какую-либо
группу фауны. Корреляция проводится по сходным стадиям развития организма. Этот
метод имеет особое значение для предварительной прикидочной корреляции разрезов;
при сопоставлении сходных или подобных по составу фаун далеко от стоящих регионов.
Метод основан на установлении закономерностей филогенеза той, или иной групп.
Определяется время появление ряда морфологических признаков, а сами признаки
приобретают биостратиграфическое значение. Примером может служить соответствие
крупных этапов развития аммоноидей определенным эротемам и системам: гониатитов –
палеозою, цератитов – триасу, аммонитов – юре и мелу. В данном случае расчлененность
лопастных линий может быть использован для установления возраста отложений в
хорошо изученных филогенетических ветвях морфологические признаки могут служить
основанием для установления и более коротких временных интервалов и может
привлекаться для конкретных сопоставлений частных разрезов. Например, эволюция
киммериджских аммонитов подсемейства Aulaostephaninae идеен в направление
постепенного понижения ребер на вентральной стороне с образованием в конце концов
гладкой полоски. В этом случае возможно установить подъярусы и даже отдельные зоны
киммериджа. Использование эволюционного метода в стратиграфии требует весьма
детальных и тщательных филогенетических построений.
Палеоэкологический метод. Биостратономия. В предшествующих разделах мы
неоднократно отмечали существенную зависимость большинства фаунистических ком-
плексов от фаций. Это широко распространенное явление — едва ли не основное
осложнение при биостратиграфических корреляциях. В конечном итоге зависимость
организмов от фациальных обстановок обусловлена их образом жизни, реконструкцией
которого занимается палеоэкология. В задачи палеоэкологии входит выявление физико-
географических (соленость, глубина, температура воды бассейнов, степень аэрации и т. п.)
и биотических (трофические связи, конкуренция, структура биоценозов) параметров
существования, древних организмов. Эти задачи решаются с помощью комплексного
использования результатов литологических, геохимических и палеонтологических
исследований. В числе последних большое значение имеют помимо работ по систематике
тафономические наблюдения, количественная оценка представителей отдельных видов и
экологических групп, изучение явлений комменсализма, выявление симбионтов, анализ
эпифаун и т. п. Методика палеоэкологических исследований детально рассмотрена в
современной геологической литературе [Геккер Р. Ф., 1957;. Захаров В. А., Юдовный Е.
П., 1967, 1974; Красилов В. А., 19'72; Рауп Д., Стэнли С., 1974; Ager, 1963; Ziegler, 1972;
Николов Т., 1977, и др.].
Палеоэкологический метод стратиграфических исследований был предложен Р. Ф.
Геккером (1940, 1948 г.) при изучении верхнедевонских отложений Главного девонского
поля, а затем нижнекаменноугольных отложений Московской синеклизы. Позднее Р. Ф.
Геккер следующим образом охарактеризовал выделенные им подразделения типа
шелонского горизонта: «...вы-. деление этих единиц основано не на присутствии форм,
свойственных только данной единице, а на изменении количественного состава фауны, а
также осадков, отражавших изменения условий осадкообразования и вместе с тем
обитания фауны в определенные этапы геологической истории» [Геккер Р. Ф., Осипова А.
И., Вельская Т. Н., 1962, кн. 1, с. 105].
Метод корреляции разрезов по палеоэкологическим и биостратономическим
признакам. Этот метод используется для дробной корреляции разрезов в пределах
довольно ограниченных районов. Основой для расчленения служит не видовой состав
органических комплексов (который, конечно, фиксируется, но не дает в узких
вертикальных интервалах заметных изменений), а палеоэкологические и
биостратономические признаки.
К палеоэкологическим признакам относятся: а) прижизненные количественные
отношения видов; б) приуроченность определенных видов к определенным уровням,
соответствующим месту и времени их жизни; в) различные следы животных, а также
приуроченность их к определенным уровням; г) прижизненная ориентировка раковин и
корневых образований растений.
Биостратономические признаки, отражающие посмертные особенности захоронения,
включают: а) вторичные, посмертные количественные отношения разных видов; б)
сохранность раковин; в) ориентировку и скопление органических остатков, которые
обусловлены механическими факторами при гибели или после отмирания организмов и т.
д.
Отсюда при сборе материала необходимо обращать внимание на количественные
отношения различных форм в последовательно сменяющих друг друга слоях; на
спорадическое нахождение этих форм или в виде скоплений на определенных уровнях в
слое; на цельность раковин, разрозненность их створок, сохранность всех органических
остатков, на прижизненную или посмертную ориентировку органических остатков в
разрезе; на присутствие в слоях следов ползания, зарывания и т. д., на частоту их
нахождения и приуроченность к различным уровням слоя.
Принимая во внимание все эти факторы, можно выявить своеобразную
палеоэкологическую характеристику каждого из похожих в общем плане друг на друга
слоев, которая прослеживается на всем протяжении отдельного слоя в пределах исследу-
емого узкого района. На основании полученных данных проводится дробное послойное
расчленение разрезов и их послойная корреляция. Этот метод особенно хорошо применим
к толщам со сменой органических комплексов в пространстве и их частой сменой с
неоднократным повторением во времени.
Примером успешного применения этого метода является расчленение и сопоставление
разрезов «толщи переслаивания» окской свиты нижнего карбона на р. Мете, выше
Боровичей (рис. 13, Р. Ф. Геккер, 1938). Отличия отдельных слоев, кажущихся с первого
взгляда тождественными, могут оказаться довольно резкими. Например, два слоя
известняка, одинаковые по мощности, с границей размыва в кровле, различаются по
характеру подошвы. У одного она ровная, у другого изобилует следами
жизнедеятельности червей. В одних слоях встречаются разрозненные крупные раковины
брахиопод, в других есть и двустворчатые. В некоторых слоях раковины не
ориентированы, в других встречаются с прижизненной ориентировкой.
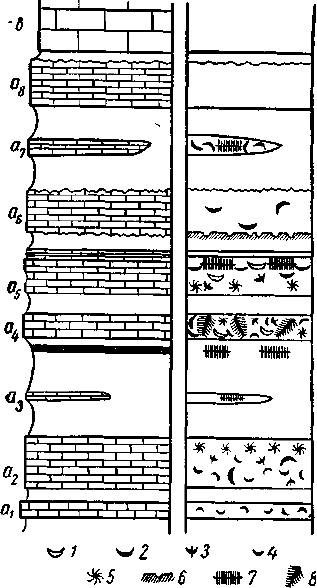
Рис. 9. Различная палеоэкологическая и
биостратономическая характеристика
отдельных слоев известняков (а\ — С8) толщи
переслаивания «а» нижнего карбона у г.
Боровичей при сохранении видового состава
фауны (Р. Ф. Геккер, 1956):
/ — раковины Gigantoproductus с обеими
створками; 2 — то же. отдельные створки; 3 —
Productus semiplanus Schwez.; 4 — мелкие
брахиоподы; 5 — Taonurus ' (Spi-. rophyton); 6 —
петлеобразные и мелкие простые ходы червей; 7
— горизонтальные стигмарии; « — вертикальные
и косые стиг-иарии. Значки брахиопод
одновременно показывают ориентировку
раковин, а также их одно- или двустворчатость.
Недостатком этого метода является то, что используемые им признаки имеют узкое,
чисто местное значение, не выдерживаются на большом протяжении, и радиус их
действия обычно ограничивается несколькими километрами.
Метод корреляции разрезов на основании закономерной смены экологических
комплексов форм в пространстве. Этот метод особенно важен для сопоставления разрезов
разнофациальных отложений, сменяющих друг друга в определенных направлениях. При
разнообразии фациального состава, сравнительно быстрой смене фаций в пространстве и
быстрой смене их с многократным повторением в вертикальном разрезе необходимо
начинать с выяснения закономерностей смены (радиальных обстановок в различных
направлениях и связанных с ними комплексов органических форм.
В силу этого в основу данного метода закладывается принцип, диаметрально
противоположный принципу, заложенному в методе руководящих форм и комплексов:
изменение, различие, непостоянство органических форм в сопоставляемых по времени
разрезах.
При этом методе для Отдельных горизонтов устанавливаются ряды, или «гаммы»,
экологических комплексов форм и осадков, закономерно сменяющих друг друга в
интересующем исследователей направлении. Выделяются типы осадков, и выясняется на-
личие в каждом типе определенного комплекса органических форм, находящегося с ним в
неразрывной связи. Затем прослеживается изменение типа осадка и изменение комплекса
форм, при котором новый комплекс также находится в тесной связи со своим типом
осадка (рис. 9).
Восстановление закономерной смены осадков и присущих им комплексов в заданном
направлении позволяет последовательно сопоставлять быстро изменяющиеся в
фациальном отношении отложения. Кроме того, отмечаются и количественные отношения
отдельных организмов в комплексах. Иногда виды, играющие в данном экологическом
комплексе второстепенную роль, приобретают одну из главных ролей в соседних фациях,
перекидывая к ним как бы мосты, подсказывая облик основных фаунистических и
флористических компонентов в соседних комплексах и помогая тем самым проводимому
сопоставлению. Хорошим примером применения этого метода является сопоставление
разрезов восточной и западной половины Главного девонского поля, проведенное Р. Ф.
Геккером (1954).
Метод выделения стратиграфических единиц и корреляции разрезов по смене
экологических комплексов форм во времени. В основу этого метода положено
выявление смены во времени органических комплексов одного экологического облика
комплексами другого экологического облика (т. е. в вертикальном, а не в горизонтальном
направлении, как у предыдущего метода). При методе же руководящих комплексов
прослеживается смена во времени комплекса одного видового состава комплексом
другого видового состава при в общем сходном экологическом облике обоих комплексов.
Если немые слои и горизонты при корреляции по руководящим комплексам являются
белыми пятнами в сопоставляемых разрезах, то при данном методе они имеют столь же
важное значение, как и слои, охарактеризованные экологическими комплексами форм.
Немые слои отражают определенные важные события, определенную обстановку в ис-
тории развития исследуемого района на соответствующем этапе, обусловившую
отсутствие в них органических остатков.
Многие стратиграфические границы приурочены к границам отличных в фациальном
отношении толщ, к границам между разными последовательно сменяющими друг друга
циклами осадконакопления, так как эти циклы обычно знаменуют завершение
определенных этапов в истории развития исследуемых территорий.
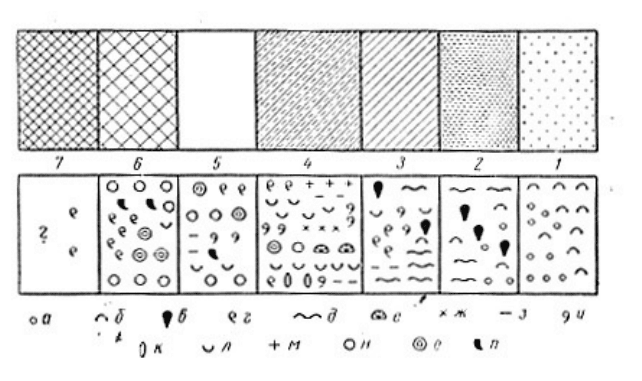
Рис. 10. Закономерная смена («гамма») пород и связанных с ними комплексов форм в морских
отложениях верхнего девона Главного девонского поля в направлении с северо-востока на юго-запад
(от берега моря в глубь водоема). (Р. Ф. Геккер, 1956):
Породы: / — красноцветные терригенные породы; 2 — белые кварцевые пески; 3—глины; 4 —
глинистые пески и мергели; 5 — чистые известняки; 6 — доломитовые известняки; 7 — доломиты.
Фауна и флора. Обитатели пресных вод: а — трохилиски; б — рыбы красноцветной фации. Обитатели
моря. Эвригалинные формы: в — лингулы; г —1 Platyschisma; д — черви (следы). Стеногалинные формы: я)
обитатели нормально-соленых вод: е—.табуляты; ж — Sptrorbis; з — пелециподы; и — гастроподы
(большинство); к — цефалоподы; л —замковые брахиоподы (большинство); м -~ иглокожие; б) обитатели
вод нормальной и повышенной солености; н — сине-зеленые водоросли (Girvanella — Pycnostroma); о —
строматопороидеи; л — ругозы
Палеоэкологические методы, наиболее тонко прослеживающие как изменения физико-
географических условий прошлого (среды обитавших тогда организмов), так и самих
организмов в зависимости от изменений среды, могут значительно помочь уточнению и
обоснованию стратиграфических границ. Мелководные отложения будут допускать при
этих методах более дробное деление, чем глубоководные, так как на них отражаются даже
сравнительно небольшие колебания уровня моря и перемещения береговой линии. Но это
будет чисто местным делением. Вообще же деление необходимо давать настолько дробно,
насколько позволяет материал, иногда даже больше требуемой узко практическими
запросами точности, так как чем детальнее проведена разбивка толщи, тем глубже понята
геологическая история района и лучше освещены связанные с ней насущные вопросы
практики.
Примером применения рассматриваемого метода является корректирование
стратиграфического расчленения верхнедевонских отложений в восточной половине
Главного девонского поля.
В свое время в разрезе этих отложений был выделен шелонский горизонт, включающий
отложения как регрессивной фазы предыдущего цикла осадконакопления внизу, так и
трансгрессивной фазы последующего цикла. Общий облик нижних и верхних слоев
обусловливался принадлежностью их к отложениям бассейна ненормальной солености:
нижнешелонские слои даже включали гипсоносные отложения лагунного характера; в
верхнешелонских попадались представители обедненной морской фауны и
проблематично морской. Так как внутри шелонского горизонта прошла граница между
циклами осадконакопления, то впоследствии этот горизонт стремились ликвидировать
совсем, отнеся нижние слои к подстилающему чудовскому горизонту, а верхние — к
свинордскому. Палеоэкологический метод, использующий смену экологических
комплексов форм во времени, показывает, что этого нельзя делать.
Нижнешелонские слои, помимо резко обедненных форм чудовского комплекса,
содержат немногочисленные, но свои органические формы, а главная их особенность
состоит в том, что их фаунистический комплекс имеет свой специфический экологиче-
ский облик. Этим он резко отличается от фаунистического комплекса чудовского
горизонта. Верхи нижнешелонских слоев абсолютно немые, но содержат полный
комплекс лагунных осадков, представляющих характернейший маркирующий горизонт.
Поэтому нижнешелонские и верхнешелонские слои, хотя и относимые к разным циклам
осадконакопления, не могут быть слиты с соседними горизонтами, а должны сохранить
подчиненную им стратиграфическую самостоятельность.
Особенности использования микропалеонтологических объектов для
биостратиграфической корреляции. Широкое использование для целей
биостратиграфии микропалеонтологических объектов — в первую очередь фораминифер,
радиолярий, остракод, кокколитофорид, тинтинид, спор и пыльцы и т. д. — началось
сравнительно недавно, немногим более полувека назад. Однако за этот сравнительно
короткий промежуток времени биостратиграфические выводы, полученные на основании
изучения перечисленных групп, послужили основой для корреляции осадочных толщ
многих регионов земного шара. В первую очередь здесь следует отметить закрытые
территории, и в особенности нефтегазоносные бассейны Америки, Ближнего Востока,
Волго-Уральской области, Западной Сибири и т. д., а также осадочный чехол океанов.
Такое быстрое и плодотворное внедрение микропалеонтологии в стратиграфию прежде
всего связано с размерами объектов исследования, которые обеспечивают возможность их
находок практически по всему разрезу (для морских организмов — по всему разрезу
морской толщи), что имеет особенно важное значение для кернового материала. Поэтому
для сопоставления разрезов скважин микропалеонтологический материал представляет
наибольшую ценность, ^так как раковины крупных организмов встречаются в керне
сравнительно редко и имеют обычно плохую сохранность, что весьма затрудняет точное
их определение. Наконец, часто отдельные находки крупномерных организмов не
позволяют использовать их для сопоставления осадочных толщ. Имеется еще одно важное
достоинство микропалеонтологических объектов для целей стратиграфии: во многих
случаях они непрерывно распределены по всему разрезу и поэтому биостратиграфические
границы, устанавливаемые по изменению комплексов фораминифер, остракод и т. д.,
являются полностью обоснованными самим палеонтологическим материалом, в то время
как дискретное распределение в разрезе крупномерных остатков фауны и флоры зачастую
вынуждает исследователей приурочивать биостратиграфические границы к ли-
тологическим разделам, конкреционным горизонтам или к тем биостратиграфическим
уровням, которые установлены по микрофауне.
Наибольшее значение в биостратиграфии получило сопоставление разрезов по
комплексам микрофауны, в первую очередь по комплексам фораминифер. Обычно
комплексы фораминифер устанавливаются путем выявления как стратиграфических
диапазонов отдельных видов, так и изменения количества экземпляров этих видов по
разрезу (рис. 10). Очень часто в состав комплекса входит несколько десятков и даже
сотни видов. Естественно, учитывать всю эту массу таксонов при биостратиграфических
корреляциях невозможно. Обычно для этих целей используется 10—20 наиболее харак-
терных, т. е. наиболее часто встречающихся и обладающих наиболее узким
стратиграфическим диапазоном, форм. Однако для общей характеристики комплекса
имеет большое значение и степень его разнообразия, и степень видового разнообразия от
дельных семейств. Так, например, комплекс фораминифер, большинство характерных
видов которого представлено трудноопределимыми агглютинирующими формами, может
содержать разное число видов с известковистой стенкой, причем иногда существенные
колебания количества видов, например лентикулин, может явиться указанием на
изменение стратиграфического по ложения комплекса. Для политаксонных комплексов
фораминифер, радиолярий, остракод четкий диагноз, содержащий общую характеристику
комплекса и обоснование его отличий от выше- и нижележащего комплексов, является
требованием со вершенно необходимым. Без выполнения этого требования не возможно
ни обоснованное прослеживание комплекса, ни воспроизведение результатов
проведенного исследования другими биостратиграфами.
Комплексы микрофауны имеют весьма различное распространение. Ассоциации
планктонных фораминифер и радиолярий кайнозоя часто прослеживаются в виде поясов,
охватывающих весь земной шар. Комплексы бентоса, как правило, имеют значительно
меньшую протяженность и приурочены обычно к определенным фациальным зонам (см.
рис. 10). Однако известны и чрезвычайно широко распространенные комплексы
бентосных фораминифер. Так, установленный Л. Г. Дайн на восточном склоне
Приполярного Урала позднекимериджский комплекс с Pseudolamarckina lopsiensis
прослежен на Тимане, в Приуралье, в Западной Сибири и на Южном Таймыре, т. е. на
расстоянии почти 2500 км. При этом если на восточном склоне Урала этот комплекс"
установлен в глинах; то в Притиманье он встречен в глинистых алевритах, в Приуралье —
в алевролитах, а на Таймыре — в глауконитовых песках. Иногда, правда, оказывается, что
широко прослеживаются не комплекс, а только отдельные его виды. В этом случае
широкие корреляции часто приводят к очень чувствительным ошибкам. Так, например, в
начале 50-х годов, когда началось интенсивное изучение разрезов Западной Сибири,
средняя часть меловой толщи была отнесена к апту на основании находок нескольких
видов, близких к аптским формам Поволжья и Прикаспия. Впоследствии, однако,
оказалось, что эта часть разреза датируется от альба до нижнего турона, а нормально-
морские аптские отложения в Западной Сибири отсутствуют вообще. В целом наибольшее
значение комплексы микрофауны бентоса имеют для сопоставления разрезов в пределах
одного бассейна или его части, и здесь они оказываются наиболее эффективными.
Как мы видим, общая процедура выделения и прослеживания комплексов
фораминифер, радиолярий, нанопланктона, тинтлннид, остракод, конодонтов и т. д. не
имеет особой специфики. Разве что необходимость четкой диагностики комплексов здесь
выступает более отчетливо. Несколько иное положение сложилось со стратиграфическим
использованием спорово-пыльцевого анализа. Как справедливо отметил В. В. Меннер
[1962], палинология имеет самое большое потенциальное значение при био-
стратиграфическом анализе, поскольку только использование спор и пыльцы в
принципе позволяет производить непосредственное сопоставление морских,
солоноватоводных и континентальных отложений.
В то же время именно благодаря этой возможности, т. е. благодаря широкому разносу
спор и пыльцы ветром, водными потоками, морскими течениями и волнениями,
интерпретация данных палинологического анализа имеет свою специфику. Эта специфика
возрастает еще более, если учесть многочисленные трудности, возникающие при
установлении и определении видовых таксонов спор и пыльцы древних растений...
Палинологический анализ впервые был использован Л. Постом в 1916 г. для
изучения торфяников. Анализ пыльцевых спектров позволил ему показать смену во
времени лесных сообществ и на этом основании сопоставить отдельные горизонты
торфяников. Идеи Л. Поста и В. Н. Сукачева были использованы И. П. Герасимовым
[1923], К. К. Марковым [1931] и другими исследователями для анализа четвертичных
осадков. Неоднократная смена похолоданий и потеплений, происходившая в
четвертичном периоде, обусловила неоднократную же смену растительных сообществ,
которая достаточно четко фиксировалась в изменении количественных соотношений
различных группировок, составляющих спорово-пыльцевые спектры. Таким образом,
использование палинологии для стратиграфии четвертичных отложений было основано, в
сущности, на палеогеографическом (климатическом) принципе. Незначительная продол-
жительность четвертичного периода, контрастность климатических изменений,
происходивших за последние 500 тыс. лет, большая вероятность захоронения пыльцы и
спор в Подмосковье в незначительном удалении от продуцирующих их растений,
возможность надежного экологического контроля и непе-реработанность четвертичной
толщи обусловили корреляцию отдельных горизонтов четвертичной системы не на
основании эволюционной смены видов, а путем прослеживания экологических изменений
в составе растительности. При таком подходе основная задача палинологов сводилась не к
определению отдельных - видов, а к подсчету количества зерен по отдельным группам,
палеогеографической (климатической) интерпретации полученных данных, выявлению
спектров, соответствующих моментам похолоданий и потеплений, и, наконец, к
сопоставлению этих спектров по площади.
При переходе к анализу более древних толщ палинологи пошли двумя путями.
Специалисты, изучавшие палеозой и верхний докембрий, вынуждены были разработать
свою систематику и сразу стали оперировать при биостратиграфичсском анализе
видовыми и родовыми таксонами. Для мезо-кайнозоя возможности сопоставления со
спорами и пыльцой современных растений, по крайней мере на уровне семейств и
классов, были значительно большими. По-видимому, именно поэтому вместе со
стремлением выделять естественные, пусть даже крупные, таксоны на мезо-кайнозой
были перенесены и методы стратиграфического анализа, разработанного для уникальных
в колонке мезо-кайнозоя четвертичных отложений. Между тем и по геологическим, и по
палеогеографическим условиям дочетвертичного времени такой перенос нельзя считать
оправданным. Прежде всего продолжительность не только отделов и ярусов, но даже зон
неогена—триаса значительно больше всего четвертичного периода. В течение всего
времени происходили неоднократные изменения ландшафта и неоднократное
перераспределение осадков.
Поскольку в подавляющем большинстве случаев древние толщи являются
неоднократно переотложенными и поскольку коррелируются самые разные их фации (от
сугубо континентальных до псевдоабиссальных), споры и пыльца выступают в роли
мельчайших осадочных частиц. Неудивительно - поэтому, что неоднократно отмечалось
изменение содержания пыльцевых зерен в разного типа породах (рис. ).
Наконец, различная продуктивность растений [Гричук В. П., Заклинская Е. Д., 1948],
различная способность спор и пыльцы к переносу ветром и водными потоками [Красилов
В. А., 1972], а также морскими течениями и волнениями [Чибрикова Е. В., 1963; Панов Д.
Г., Вронский В. А., 1964] настолько искажают первичный состав спорово-пыльцевых
комплексов, что их соотнесение с конкретными ландшафтами, а главное, прослеживание
на значительные расстояния вызывает большие сомнения. Неудивительно поэтому, что
многие палинологи видят главное назначение спорово-пыльцевых спектров древних толщ
в различного рода палеогеографических реконструкциях [Гри-чукВ. П., Заклинская Е. Д.,
1948]. Действительно, интегральный характер спорово-пыльцевых комплексов,
бесспорно, может дать весьма большую информацию о палеоландшафтах [Палеофито-
география..., 1975].
Иногда появление определенных групп позволяет сделать чрезвычайно важные
выводы о палеоклиматических изменениях.
Например, появление пыльцы Glossopolis в верхнеюрских отложениях на юге Западной
Сибири [Палеозойские и мезозойские флоры..., 1972] позволило установить здесь
аридизацию климата и т. д.
Для целей же стратиграфии более перспективен переход к выявлению видовых
таксонов, изучению их распространения в разрезах и обычной биостратиграфйческой
интерпретации спорово-пыльцевых комплексов. Несмотря на большие сложности в
выделении видов, сама методика определения образцов станет при этом существенно