Страйер Л. Биохимия. Том 3
Подождите немного. Документ загружается.

Вопросы и задачи
1. Образование Ile-тРНК происхо-
дит через связанный с фермен-
том промежуточный продукт Ilе-АМР. Бу-
дет ли, по вашему мнению, образовываться
32
Р-меченный АТР из
32
РР
i
, если инкубиро-
вать каждый из следующих наборов компо-
нентов в присутствии специфического акти-
вирующего фермента?
а) АТР и
32
PP
i
.
б) тРНК, АТР и
32
РР
i
.
в) Изолейцин, АТР и
32
РР
i
.
2. Из бактерий, выращенных на
«тяжелой» (содержащей
13
С и
15
N) и на «легкой» (
12
С и
l4
N) среде, получи-
ли рибосомы. Эти 70S-рибосомы добавили
в систему с активным синтезом белка in
vitro. Через несколько часов из смеси ото-
брали пробу и проанализировали ее центри-
фугированием в градиенте плотности.
Сколько полос 70S-рибосом вы ожидаете
увидеть в градиенте плотности?
3. Сколько богатых энергией фос-
фатных связей затрачивается на
синтез белка из 200 остатков, если исходить
из готовых аминокислот?
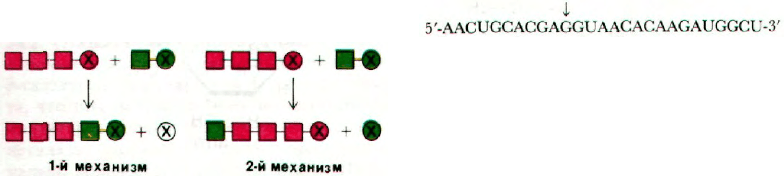
4. Существует два основных меха-
низма элонгации биологических
молекул (рис. 27.26). 1-й механизм основан
Рис. 27.26. Два механизма элонгации.
на отщеплении активирующей группы (по-
меченной на рисунке крестиком) от расту-
щей цепи. При синтезе по 2-му механизму
активирующая группа отщепляется от при-
соединяющегося к растущей цепи мономе-
ра. Укажите, по какому из механизмов, 1-му
или 2-му, протекают следующие реакции
биосинтеза.
а) Синтез гликогена.
б) Синтез жирных кислот.
в) С
5
—> С
10
—> С
15
при синтезе
холестерола.
г) Синтез ДНК.
д) Синтез РНК.
е) Синтез белка.
5. Мутации, в результате которых
возникают терминирующие ко-
доны, называются нонсенс-мутациями. Эти
мутации могут быть супрессированы изме-
ненными тРНК. Например, мутантная РНК
считывает кодон UGA как триптофановый
кодон. Какое наиболее вероятное замеще-
ние основания произошло в этой мутантной
тРНК?
6. Придумайте реактив для кова-
лентного мечения по сродству
участков связывания тРНК в рибосоме. Как
бы вы синтезировали такой реактив?
7. мРНК-транскрипт одного гена
фага Т7 содержит следующую
последовательность оснований:
Предскажите, к какому результату приведет
мутация отмеченного стрелкой G при его
замене на А.
8. Что общего между коррекцией
ошибок при синтезе белка
и ДНК?
Дополнительные вопросы см.: Wood W. В.,
Wilson J.H., Benbow R.M., Hood L. E.,
Biochemistry: A Problems Approach,
Benjamin, 1974, ch. 18.
ГЛАВА 28
Регуляция выражения
гена в фенотипе
Мы уже видели, что активность многих бел-
ков регулируется с помощью различных ме-
ханизмов, например протеолитической ак-
тивации, аллостерических взаимодействий
и ковалентной модификации. В этой главе
рассматривается регуляция скорости синте-
за белка, которая также играет принци-
пиально важную роль в обшей картине ме-
таболизма клетки. У бактерий активность
гена регулируется в основном на уровне
транскрипции, а не трансляции. Мы сосредо-
точим внимание на лактозном и триптофа-
новом оперонах Е. coli и на регуляторных
аспектах цикла развития бактериофага λ,
так как молекулярные механизмы регуля-
ции в этих системах хорошо изучены. Более
того, именно интенсивное исследование
этих систем позволило сформулировать не-
которые общие принципы регуляции выра-
жения гена в фенотипе (экспрессии гена)
у прокариот и вирусов. Экспрессия гена у эу-
кариот регулируется иначе, как это станет
очевидно из следующей главы.
28.1. β-Галактозидаза -
индуцибельный фермент
Е. coli может использовать лактозу в каче-
стве единственного источника углерода.
112
Часть IV.
Информация
Главный фермент в метаболизме этого са-
хара - β-галактозидаза, гидролизующая
лактозу на галактозу и глюкозу (рис. 28.1).
При выращивании на лактозе клетка Е. coli
содержит несколько тысяч молекул β-галак-
тозидазы. Если же выращивать Е. coli на
других источниках углерода, например на
глюкозе или глицероле, то число молекул
β-галактозидазы на клетку не достигает де-
сяти. Лактоза индуцирует значительное уве-
личение количества β-галактозидазы в клет-
ке Е. coli, причем она вызывает синтез новых
молекул фермента, а не активирует профер-
мент (рис. 28.2). Следовательно, β-галакто-
зидаза - индуцибельный фермент. Одновре-
менно и согласованно с β-галактозидазой
синтезируются еще два белка - галактозид-
пермеаза и тиогалактозид-трансацетилаза.
Пермеаза необходима для переноса лак-
тозы через бактериальную клеточную мем-
брану, трансацетилаза же не имеет суще-
ственного значения для метаболизма лак-
тозы. Физиологическая роль трансацети-
лазы пока не установлена, in vitro она ката-
лизирует перенос ацетильной группы аце-
тил-СоА на гидроксильную группу при С-6
тиогалактозида.
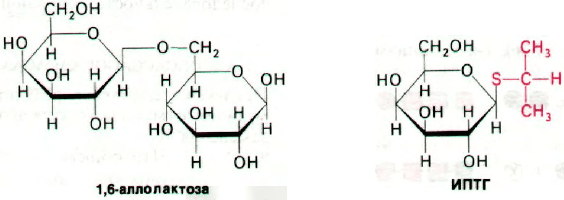
Физиологическим индуктором β-галакто-
зидазы является аллолактоза, которая обра-
зуется из лактозы в результате реакции
трансгликозилирования. Синтез аллолак-
тозы катализируется теми несколькими мо-
лекулами β-галактозидазы, которые имеют-
ся в клетке еще до индукции. Изучение
природы индукторов показало, что неко-
торые β-галактозиды служат индукторами,
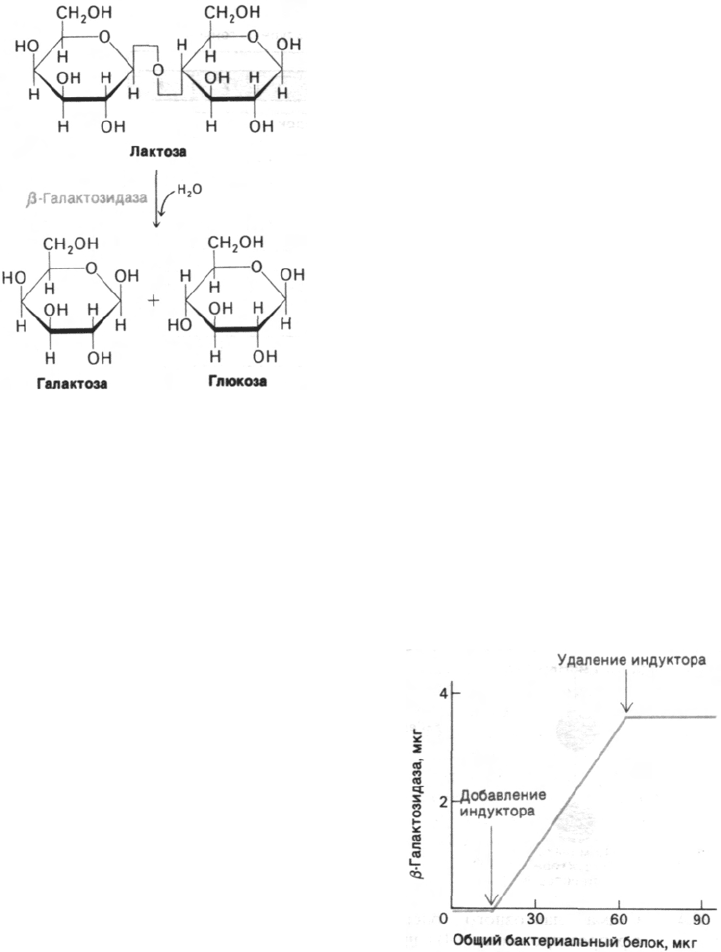
Рис. 28.1. β-Галактозидаза гидролизует
лактозу.
не являясь при этом субстратами β-галакто-
зидазы, тогда как другие соединения ведут
себя как субстраты, не будучи при этом ин-
дукторами. Например, изопропилтиогалак-
тозид (ИПТГ) неметаболизируемый индук-
тор (его также называют холостым индук-
тором).
28.2. Открытие регуляторного гена
Ключом к изучению механизма индукции
β-галактозидазы стало открытие того фак-
та, что под действием всех исследованных
индукторов количество пермеазы и транса-
цетилазы возрастало прямо пропорцио-
нально количеству β-галактозидазы. Даль-
нейший прогресс был достигнут при изуче-
нии мутантов. Оно показало, что β-галакто-
зидаза, пермеаза и трансацетилаза коди-
руются тремя генами - z, у и а соответствен-
но - которые расположены друг за другом.
Были получены мутанты, утратившие спо-
собность к синтезу одного из этих белков.
Например, генотип z
--
y
+
a
+
обозначает, что
у данного мутанта нет β-галактозидазы, но
имеются в нормальном количестве пермеа-
за и трансацетилаза. Наибольший интерес
представляет класс мутантов, у которого
мутацией затронуты все три белка. Такие
конститутивные мутанты синтезируют без
всякого индуктора большие количества
β-галактозидазы, пермеазы и трансацети-
лазы. Франсуа Жакоб и Жак Моно (Francois
Jacob, Jacques Monod) пришли к выводу, что
скорость синтеза этих трех белков зависит
от какого-то общего элемента, отличного
от генов, кодирующих их последовательно-
сти. Ген этого общего регуляторного эле-
мента был обозначен i. Индуцибельные бак-
терии дикого типа имеют генотип
i
+
z
+
y
+
a
+
,
а
конститутивные мутанты
по
лактозным генам - генотип i
--
z
+
y
+
a
+
.
Каким образом осуществляется влияние
гена i на скорость синтеза белков, коди-
руемых генами z, у и а? Проще всего было
предположить, что ген i кодирует синтез не-
коего компонента цитоплазмы, названного
репрессором, которого либо совсем нет в
i
--
-клетках, либо он там неактивен. Эта идея
была проверена в ряде изящных генетиче-
ских экспериментов с частично диплоидны-
ми бактериями, имевшими два набора генов
лактозной области. Один набор содержался
в бактериальной хромосоме, а второй - в по-
ловом факторе F', введенном в клетку при
конъюгации. Например, был получен ди-
плоид i
+
z
--
/Fi
--
z
+
. У этого диплоида гены
i
+
z
--
находятся в хромосоме, а гены i
--
z
+
- в
эписоме. Является ли этот диплоид индуци-
бельным или конститутивным в отношении
β-галактозидазы? Другими словами, может
ли ген i
+
бактериальной хромосомы подав-
лять экспрессию гена z
+
, расположенного
в эписоме? Эксперимент дал совершенно
Рис. 28.2. Увеличение количества β-га-
лактозидазы идет параллельно
увеличению числа клеток в рас-
тущей культуре E.coli. Наклон
этого графика указывает, что
β-галактозидаза составляет
6,6% всего синтезируемого бел-
ка.
28. Регуляция выражения
гена в фенотипе
113
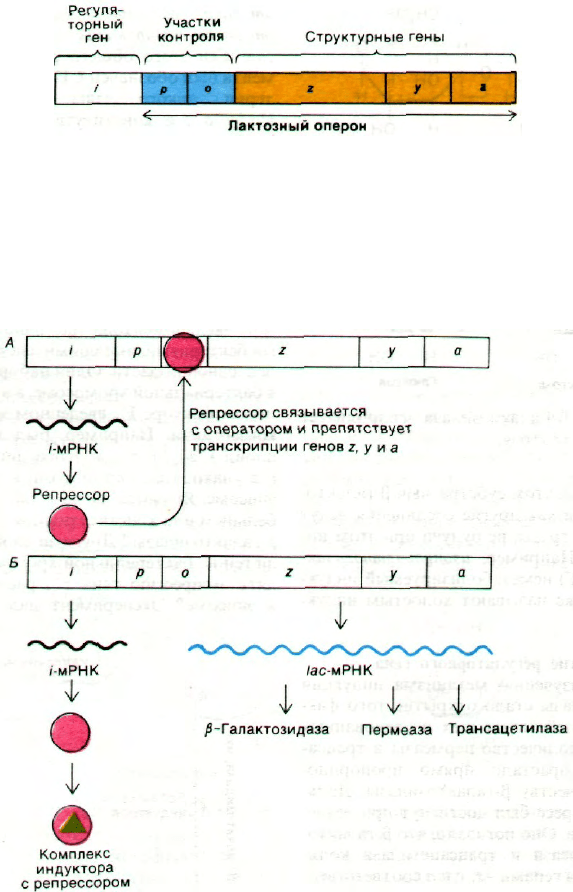
Рис. 28.3. Карта лактозного оперона
и его регуляторного гена.
В карте масштаб не соблюден:
участки р и о в действительно-
сти гораздо меньше, чем коди-
рующие участки генов.
Рис. 28.4. Схема лактозного оперона
в репрессированном (А) и инду-
цированном (Б) состояниях.
четкий результат: диплоид индуцибелен, а не
конститутивен. Такой же результат был по-
лучен и в отношении диплоида i
--
z
+
/Fi
+
z
--
.
Следовательно, ген i кодирует способный
к диффузии репрессор.
114
Часть IV.
Информация
28.3. Оперон - единица координированной
генетической экспрессии
На основе только что описанных экспери-
ментов Жакоб и Моно постулировали мо-
дель оперона, объясняющую регуляцию бел-
кового синтеза. Генетические элементы этой
модели - регуляторный ген, операторный
ген и набор структурных генов (рис. 28.3).
Регуляторный ген продуцирует репрессор,
который может взаимодействовать с опера-
торным геном. В дальнейшем выяснилось,
что репрессор представляет собой белок.
Операторный ген расположен рядом по со-
седству со структурными генами, которые
он контролирует. Связывание репрессора
с операторным геном препятствует тран-
скрипции структурных генов. Операторный
ген в совокупности со структурными гена-
ми, рядом с которыми он расположен, назы-
вается опероном. В случае лактозного оперо-
на ген i - регуляторный ген, ген о - ген-опе-
ратор, а гены z, у и а - структурные гены.
Кроме того, существует еще промоторный
участок (обозначаемый символом р) для
связывания РНК-полимеразы. Этот участок
инициирования транскрипции расположен
перед операторным геном. Индуктор, на-
пример изопропилтиогалактозид (ИПТГ),
связывается с репрессором и тем самым на-
рушает его взаимодействие с операторным
геном. Теперь гены z, у и а могут транскри-
бироваться. При этом образуется одна
длинная молекула РНК, кодирующая все
три белка (рис. 28.4). Молекулу мРНК, ко-
дирующую более одного белка, называют
полицистронным (или полигенным) тран-
скриптом.
28.4. lас-Репрессор - тетрамерный белок
При выделении репрессора лактозного опе-
рона (lас-репрессор) была использована его
Рис. 28.5. Электронная микрофотогра-
фия lас-репрессора, связанного
с ДНК, содержащей lас-опера-
тор. (Печатается с любезного
разрешения д-ра Jack Griffith.)
способность связываться с ИПТГ. Уолтер
Гилберт и Бенно Мюллер-Хилл (Walter
Gilbert, Benno Muller-Hill) установили, что
lас-репрессор - белок, который связывается
с ДНК, содержащей lас-оперон, но не связы-
вается ни с одной другой ДНК. Как и пред-
полагалось, ИПТГ подавляет связывание
lас-репрессора с lас-операторной ДНК.
Клетка Е. coli дикого типа содержит всего
около десяти молекул lас-репрессора. При
очистке репрессора возникли трудности,
связанные с тем, что он составляет всего
0,001% общего содержания белка. Однако
существуют i
sq
-мутанты, имеющие, по-види-
мому, более эффективный промотор гена i.
У этих мутантов образуется гораздо боль-
ше lас-репрессора. Количество lас-репрессо-
ра увеличивается еще сильнее, если исполь-
зовать трансдуцирующие фаги, несущие
область lас. Зараженные таким фагом клет-
ки Е. coli содержат около 20 000 молекул ре-
прессора (примерно 2% всего белка). Это
прекрасный исходный материал для выделе-
ния lас-репрессора.
Репрессор - тетрамер из идентичных
субъединиц с массой 37 кДа, каждая из ко-
торых имеет один участок связывания с ин-
дуктором. Константа диссоциации ИПТГ
составляет примерно 10
-6
М. Репрессор
очень прочно и быстро связывается с опера-
тором. Константа диссоциации комплекса
репрессор-оператор составляет примерно
10
-13
М. Такое высокое сродство обуслов-
лено тем, что в клетке Е. coli дикого типа со-
держится всего несколько молекул реп-
рессора. Константа скорости ассоциации
поразительно высока - 7•10
9
м
-1
•с
-1
. Из
этого следует, что репрессор находит опера-
торный участок, диффундируя вдоль моле-
кулы ДНК (одномерный поиск), а не ищет
его, находясь в водной среде (трехмерный
поиск)
1
.
28.5. Последовательность оснований в
lас-операторе симметрична
Наличие чистого lас-репрессора сделало
возможным выделение lас-оператора
1
Недавно проведенные исследования показали,
что ситуация намного сложнее: поиск оператора
репрессором включает оба указанных механиз-
ма и в очень большой степени зависит от ион-
ного состава среды.- Прим. перев.
28. Регуляция выражения
гена в фенотипе
115
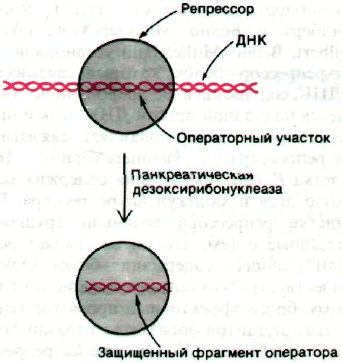
Рис. 28.6. lас-Репрессор защищает lас-
оператор от расщепления пан-
креатической дезоксирибону-
клеазой.
и определение его последовательности ос-
нований. Гилберт и его сотрудники, воздей-
ствовав ультразвуком на ДНК фага, содер-
жащего lас-область, получили фрагменты
длиной примерно 1000 пар оснований.
К смеси фрагментов добавили lас-рспрессор
и затем профильтровали раствор через ни-
троцеллюлозную мембрану. Фрагменты
ДНК, не связанные с lас-репрессором, про-
шли через фильтр, а комплексы ДНК-ре-
прессор прочно связались с ним. Связанную
ДНК сняли с фильтра с помощью ИПТГ.
Эти элюированные фрагменты ДНК обра-
ботали панкреатической дезоксирибону-
клеазой в присутствии репрессора. Идея
этого этапа эксперимента состояла в том,
что операторный участок, связанный в ком-
плексе с репрессором, должен быть защи-
щен от расщепления дезоксирибонуклеазой
(рис. 28.6). Затем установили последова-
тельность оснований этих операторных
фрагментов, определив последовательность
транскрибированной с этих фрагментов
РНК. Последовательность оснований этого
участка оказалась очень интересной: 28 пар
оснований расположены симметрично отно-
сительно оси второго порядка (рис. 28.7).
Симметрия репрессора, по-видимому, со-
ответствует симметрии оператора. Этот
принцип узнавания приложим также к сиг-
116
Часть IV.
Информация
налам терминирования транскрипции
(разд. 28.8) и к связыванию пептидного
антибиотика актиномицина D с ДНК
(разд. 25.18)
1
.
28.6. Циклический AMP стимулирует
транскрипцию многих индуцибельных
катаболических оперонов
Давно уже известно, что в клетках Е. coli,
растущих на глюкозе, активность катаболи-
ческих ферментов, таких, как β-галактозида-
за, галактокиназа, арабинозо-изомераза
и триптофаназа, находится на очень низком
уровне. Очевидно, было бы совершенно из-
лишним синтезировать эти ферменты в ус-
ловиях изобилия глюкозы
2
. Молекулярный
механизм такого ингибирующего действия
глюкозы, получивший название катаболит-
ная репрессия, был расшифрован. Ключом
к выяснению этого вопроса явились данные
о том, что глюкоза понижает концентрацию
циклического AMP в клетках Е. coli. Затем
обнаружилось, что экзогенный циклический
AMP снимает состояние репрессии, обусло-
вленное присутствием глюкозы. Последую-
щие биохимические и генетические исследо-
вания показали, что циклический AMP
стимулирует инициирование транскрипции
многих индуцибельных оперонов.
Циклический AMP, синтезированный
в отсутствие глюкозы, связывается с БАК
(белковый активатор катаболизма) - бел-
ком, представляющим собой димер из
субъединиц массой 22 кДа. Комплекс БАК
с циклическим AMP стимулирует тран-
скрипцию, связываясь вблизи от многих
промоторных участков; БАК без AMP та-
кой способностью не обладает. Опытами по
расщеплению дезоксирибонуклеазой было
установлено, что в случае lас-оперона БАК
связывается рядом с участком связывания
РНК-полимеразы. Точнее, БАК защищает
от расщепления нуклеотиды от —87 до
—49, а РНК-полимераза - нуклеотиды от
1
При детальном изучении механизма воздей-
ствия laс-репрессора с оператором оказалось,
что вазимодействию не свойственна такая сим-
метрия, так что пока остается не совсем ясным,
почему последовательности laс-оператора (и
многих других операторов) обладают элемента-
ми симметрии,- Прим. перев.
2
Далеко не очевидно, почему клетки E.coli, как
и многие другие клетки, предпочитают именно
глюкозу всем остальным источникам углерода
и энергии и используют ее до тех пор, пока ее
содержание не будет исчерпано даже в присут-
ствии значительного избытка других сахаров.—
Прим. перев.
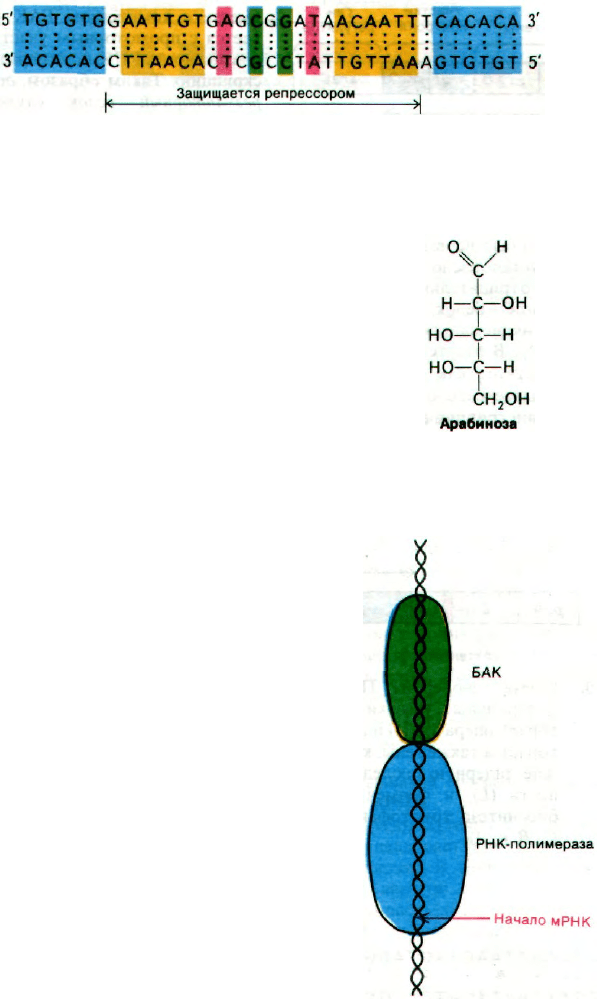
Рис. 28.7. Нуклеотидная последователь-
ность lac-оператора. Симме-
тричные участки закрашены
одинаковым цветом.
—48 до +5 (рис. 28.8). При этой системе
нумерации первый транскрибируемый ну-
клеотид обозначается +1. Последователь-
ность оснований ДНК, узнаваемая БАК,
обладает симметрией второго порядка, с ко-
торой мы постоянно сталкиваемся при рас-
смотрении взаимодействий белков с ДНК.
Каким образом БАК стимулирует инициа-
цию синтеза лактозной мРНК в 50 раз? По-
следовательное расположение неперекры-
вающихся участков связывания БАК
и РНК-полимеразы дает основание думать,
что связывание БАК с ДНК порождает
новые участки взаимодействия РНК-поли-
меразы с ДНК. Лактозный репрессор, на-
против, связывается с нуклеотидами от —3
до +21, которые в значительной степени
перекрываются с участком РНК-полиме-
разы (нуклеотиды от —48 до +5). Таким
образом, репрессор препятствует инициа-
ции, блокируя связывание РНК-полимеразы.
По всей вероятности, комплекс циклический
AMP-БАК действует таким же образом
и на другие индуцибельные опероны. Итак,
индуцибелъные опероны контролируются
с помощью комплементарных механизмов,
которые используют в качестве сигнальных
молекул циклический AMP и специфические
индукторы.
28.7. Различные формы одного и того же бел-
ка активируют и ингибируют
транскрипцию арабинозного оперона
Бактерии могут использовать арабинозу
в качестве источника энергии, превра-
щая ее в ксилулозо-5-фосфат - промежуточ-
ный продукт пентозофосфатного пути
(разд. 15.4). Превращение арабинозы в кси-
лулозо-5-фосфат происходит в результате
последовательного действия арабинозо-
изомеразы, рибулокиназы и рибулозо-5-
фосфат-эпимеразы - ферментов, кодируе-
мых генами araА, araВ и araD соответ-
ственно. Эти структурные гены вместе
с промотором (araI) и оператором (araО)
образуют арабинозный оперон (рис. 28.9).
Этот оперон регулируется геном (araС), рас-
положенным рядом с операторным участ-
ком. Подобно лактозному оперону, араби-
нозный оперон может быть активирован
комплексом БАК с циклическим AMP. Ара-
Рис. 28.8. На схеме показаны БАК
и РНК-полимераза, сидящие
на ДНК-матрице. Положение
этих белков было определено
с помощью расщепления ну-
клеазой.
28. Регуляция выражения
гена в фенотипе
117
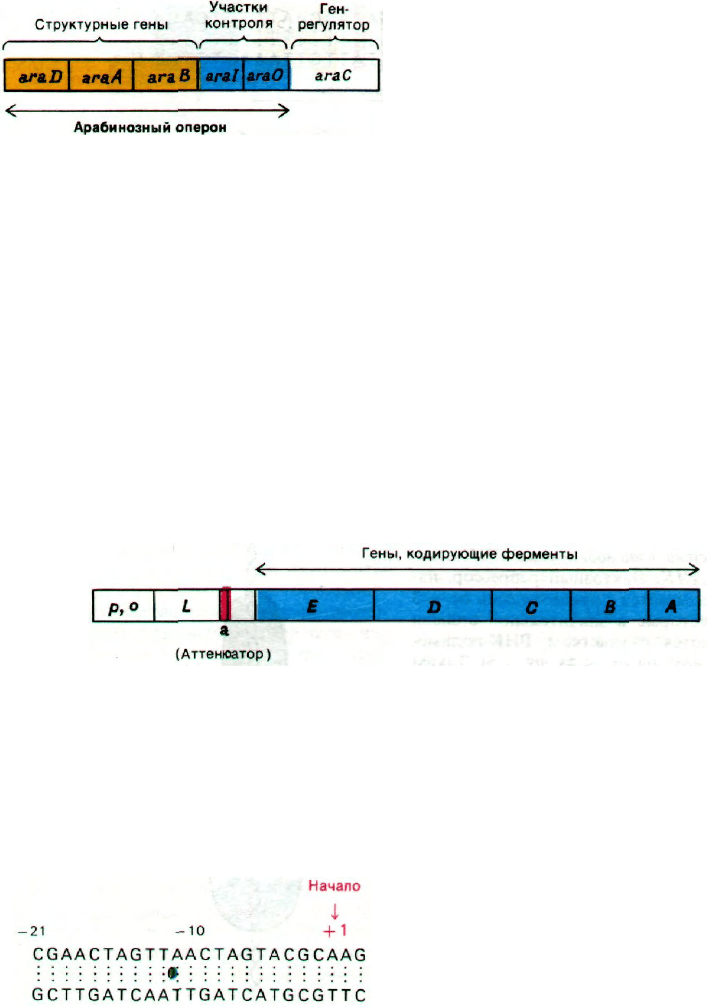
Рис. 28.9. Карта арабинозного оперона
и его регуляторного гена.
бинозный оперон был первым, в отношении
которого было установлено наличие и поло-
жительной, и отрицательной регуляций.
Продукт гена аrаС - белок, который суще-
ствует в двух функционально различных со-
стояниях Р
1
и Р
2
. В отсутствие арабинозы
этот белок действует как репрессор (Р
1
).
Форма P
1
связывается с оператором, что
препятствует транскрипции оперона. Араби-
ноза снимает Р
1
с оператора и смещает кон-
формационное равновесие в сторону Р
2
(ак-
тивирующей формы). Затем форма Р
2
Рис. 28.10. Схема trp-оперона. Показаны
контрольные участки - промо-
тор (р), оператор (о) и аттенюа-
тор (а), а также гены, кодирую-
щие лидерную последователь-
ность (L), и ферменты пути
биосинтеза триптофана (E, D,
С, В и А).
Рис. 28.11. Последовательность основа-
ний trp-оператора. Ось симме-
трии второго порядка обозна-
чена зеленым. Пара оснований,
отмеченная +1,- начало тран-
скрибируемой части оперона.
118
Часть IV.
Информация
вместе с комплексом БАК-сАМР связы-
вается с промоторным участком, что дает
возможность РНК-полимеразе начать тран-
скрипцию. Таким образом, один и тот же
регуляторный белок служит положи-
тельным и отрицательным регулятором.
28.8. Транскрипция триптофанового оперона
регулируется и аттенюатором, и оператором
Еще один регуляторный элемент был от-
крыт Чарльзом Янофски (Charles Yanofsky)
и его коллегами при изучении триптофано-
вого оперона E. coli. Транскрибируемая
с этого оперона мРНК длиной 7kb кодирует
пять ферментов, превращающих хоризмат
в триптофан (разд. 21.9). Эти пять белков
синтезируются при трансляции полици-
стронной trp-мРНК последовательно, коор-
динированно и в эквимолярных количе-
ствах. Трансляция начинается раньше, чем
заканчивается транскрипция. trp-мРНК син-
тезируется примерно за 4 мин и затем бы-
стро разрушается. Короткое время жизни
trp-мРНК, составляющее всего около 3 мин,
позволяет бактериям быстро реагировать
на изменяющуюся потребность в триптофа-
не. Е. coli может менять скорость образова-
ния ферментов биосинтеза триптофана бо-
лее чем в 700
раз.
Как осуществляется эта регуляция?
Первый уровень регуляции достигается пу-
тем взаимодействия специфического репрес-
сора с trp-операторным участком ДНК. trp-
репрессор - белок с массой 58 кДа, коди-
руемый геном trpR, удаленным от trp-оперо-
на на довольно большое расстояние. Ком-
плекс этого репрессора и триптофана прочно
связывается с оператором, тогда как сам по
себе репрессор с ним не связывается. Други-
ми словами, триптофан является корепрессо-
ром. Мишень, на которую действует ком-
плекс триптофана с репрессором,- участок
ДНК, обладающий симметрией второго по-
рядка (рис. 28.11); и в этом случае симме-
трия играет важную роль во взаимодей-
ствии белка с ДНК. Этот операторный
участок перекрывается с промоторным
участком инициирования транскрипции. Та-
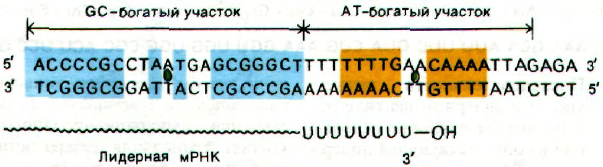
Рис. 28.12. Последовательность основа-
ний аттенюаторного участка
trp. Связанные симметрией
второго порядка пары основа-
ний GC-богатой области пока-
заны синим цветом; такие же
пары AT-богатой области по-
казаны желтым цветом.
ким образом, связывание trp-penpeccopa
с оператором препятствует связыванию
РНК-полимеразы с trp-промотором, и гены
trp не транскрибируются.
В течение некоторого времени считалось,
что ингибирование конечным продуктом ка-
талитической активности первого фермен-
тативного комплекса в пути биосинтеза
триптофана (разд. 21.11) и система ингиби-
рования транскрипции с участием репрессо-
ра и оператора - основные регуляторные си-
стемы биосинтеза триптофана. Эта точка
зрения была неожиданно опровергнута, ког-
да было обнаружено, что у некоторых му-
тантов с делециями между оператором и ге-
ном первого фермента (trpE) в опероне
происходит повышенное образование
trp-мРНК. К тому же анализ 5'-концевой по-
следовательности trp-мРНК показал, что
там имеется лидерная последовательность
длиной 162 нуклеотида, расположенная
перед инициирующим кодоном trpE. Затем
оказалось, что делеции, повышающие со-
держание trp-мРНК, картируются в этой ли-
дерной области, примерно в 30-60 нуклеоти-
дах от начала гена trpE. Следующее
поразительное наблюдение состояло в том,
что при высокой концентрации триптофана
образовывался транскрипт, содержащий
всего 130 нуклеотидов лидерной последова-
тельности ; при нехватке же триптофана син-
тезировалась trp-мРНК длиной 7000 ну-
клеотидов, включающая полную лидерную
последовательность. Отсюда Янофски сде-
лал вывод, что транскрипция trp-оперона
должна регулироваться участком контро-
лируемой терминации, называемым атте-
нюатором. Он локализован между операто-
ром и геном первого фермента пути
биосинтеза триптофана. Этот участок
терминации, регулируемый физиологиче-
скими условиями, подобен участкам тер-
минации в конце других оперонов
(разд. 25.15); он содержит GC-богатую по-
следовательность и следом за ней АТ-бо-
гатый участок. Каждый из этих участков ат-
тенюатора обладает симметрией второго
порядка (рис. 28.12). Кроме того, термини-
рованный лидерный транскрипт заканчи-
вается несколькими U подряд.
Аттенюаторный участок дополняет опе-
ратор в регуляции транскрипции trp-генов.
При изобилии триптофана инициация тран-
скрипции блокируется в результате связыва-
ния комплекса триптофан-репрессор с опе-
ратором. По мере снижения концентрации
триптофана в клетке репрессия снижается
и начинается транскрипция. Однако неко-
торые молекулы РНК-полимеразы поки-
дают матрицу, дойдя до аттенюатора, тогда
как другие продолжают синтезировать пол-
ную trp-матрицу. По мере исчерпания трип-
тофана увеличивается доля молекул РНК-
полимеразы, проходящих через аттенюа-
торный участок.
28.9. Аттенюация опосредуется трансляцией
лидерной мРНК
Каким образом аттенюаторный участок trp-
оперона улавливает концентрацию трипто-
фана в клетке? В решении этого вопроса
особенно важную роль сыграли данные
о том, что часть лидерной мРНК трансли-
руется. Весьма существенно, что в 14-член-
ном лидерном полипептиде (рис. 28.13)
имеются остатки триптофана в положениях
10 и 11. Когда триптофан находится в избы-
тке, синтезируется полный лидерный пеп-
тид. Но если триптофана не хватает, рибо-
сома задерживается на двух располо-
женных тандемом кодонах UGG, поскольку
в этом случае оказывается недостаточным
содержание триптофанил-тРНК. Застряв-
28. Регуляция выражения
гена в фенотипе
119
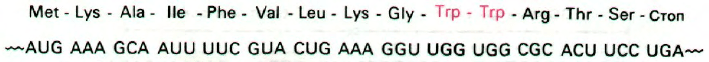
Рис. 28.13. Последовательность амино-
кислот в лидерном пептиде trp
и последовательность основа-
ний в соответствующей лидер-
ной мРНК.
шая рибосома каким-то образом меняет
структуру мРНК, так что РНК-полимераза
транскрибирует оперон за пределами атте-
нюаторного участка. Ключевой аспект это-
го регуляторного механизма состоит в том,
что трансляция и транскрипция тесно сопря-
жены между собой. Рибосомы, транслирую-
щие лидерную trp-мРНК, следуют непос-
редственно за молекулой РНК-полимеразы,
транскрибирующей ДНК-матрицу. Иссле-
дования, проведенные в последние годы, по-
казали, что застрявшая рибосома изменяет
вторичную структуру мРНК: конформация,
при которой основания спариваются, благо-
приятствуя тем самым терминированию
транскрипции, изменяется таким образом,
что РНК-полимераза проскакивает атте-
нюатор (рис. 28.14). Мы начинаем пони-
мать, что молекулы нуклеиновых кислот,
как и белковые молекулы, могут принимать
различные конформации и что изменения
конформации регулируются и имеют дале-
ко идущие физиологические последствия.
28.10. Аттенюаторный участок
гистидинового оперона содержит семь
гистидиновых кодонов подряд
В настоящее время известны еще два оперо-
на биосинтеза аминокислот у E. coli, содер-
жащих аттенюаторные участки. Фенилала-
ниновый оперон и гистидиновый оперон,
подобно триптофановому оперону, содер-
жат регулируемые участки терминации
перед первым геном, кодирующим фермент.
И в этих случаях лидерная область перед
участком терминации транслируется. Уди-
вительна последовательность аминокислот
в лидерном пептиде фенилаланинового опе-
рона: 7 из 15 остатков - фенилаланины
(рис. 28.15). Еще поразительнее лидерный
пептид гистидинового оперона: он содер-
жит семь остатков гистидина подряд. Оче-
видно, что эти лидерные мРНК предназна-
120
Часть IV.
Информация
чены, чтобы улавливать концентрации фе-
нилаланина и гистидина. Если соответ-
ствующих аминоацилированных тРНК не
хватает, трансляция лидера останавливает-
ся. Как уже обсуждалось выше на примере
trp-оперона, считается, что застрявшая ри-
босома таким образом изменяет конформа-
цию мРНК, что у нее происходит спарива-
ние оснований. Это дает возможность
РНК-полимеразе проскакивать аттенюа-
торный участок
1
. Присутствие семи после-
довательно расположенных кодонов ги-
стидина в лидерной мРНК гистидинового
оперона существенно увеличивает чувстви-
тельность этого детектора. Действительно,
падение концентрации гистидил-тРНК на
15% вызывает троекратное увеличение чис-
ла молекул мРНК, транскрибируемых с это-
го оперона.
28.11. Репрессоры и активаторы
детерминируют развитие умеренных фагов
Обратимся теперь к роли репрессоров и ак-
тиваторов транскрипции в регуляции жиз-
ненного цикла бактериофага лямбда (λ). Зре-
лая вирусная частица состоит из линейной
двухспиральной молекулы ДНК (48 kb), упа-
кованной в белковую оболочку. Существует
два пути развития вируса: он может разру-
шить клетку-хозяина или он может стать ее
компонентом (отсюда и название - уме-
ренный). При литическом пути развития
происходит полное выражение (экспрессия)
фаговых генов, что приводит к лизису бакте-
рии и образованию примерно 100 вирусных
частиц потомства. В другом случае развитие
фага λ может пойти по пути лизогенизации
клетки, когда его ДНК становится кова-
лентно связанной с ДНК клетки-хозяина
в строго определенном месте (сайт-специфи-
ческая интеграция). Этот процесс рекомби-
нации, в котором участвует кольцевая моле-
кула ДНК фага λ, мы обсудим ниже
(разд. 30.16). Когда ДНК фага интегрирует
с ДНК клетки-хозяина, большинство фа-
говых функций выключается. Фаговая ДНК
в таком состоянии называется профагом,
а клетка-хозяин, содержащая профаг - лизо-
генной бактерией. Профаг реплицируется
1
Автор противоречит собственному утвержде-
нию, что застрявшая рибосома разворачивает
мРНК (см. предыдущий разд.); рибосома дей-
ствительно способствует именно разворачива-
нию мРНК.- Прим. перев.