Сиволоб А.В., Рушковський С.Р., Кир’яченко С.С. та ін. Генетика. Підручник
Подождите немного. Документ загружается.

Розділ 6. Генетика багатоклітинних еукаріотів
181
Природу межі, яка відділяє гетерохроматин у центромерній зоні, не
з'ясовано, але гранична ділянка містить свій специфічний маркер –
метильований Lys4 гістона Н3 (і деметильований Lys9).
HD
HMT
HD
HMT
HD
HMT
Ацетильовані
х
вости
Me-Lys9
Д
еацетильовані
х
вости
Нуклеосоми
НР1
Гістон-
д
еацетилаза
Гістон-
метилтрансфераза
Межа
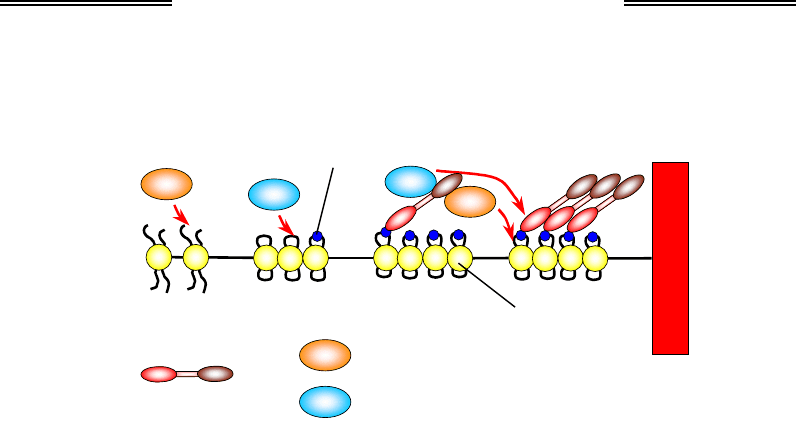
Рис. 6.9. Розповсюдження та самопідтримання
гетерохроматинового стану в центромерах
У процесі реплікації, що здійснюється нерівномірно в різних діля-
нках (гетерохроматин реплікується в останню чергу), білки тимчасово
знімаються з ДНК у реплікативній вилці (розділ 1). За точкою реплі-
кації гістони батьківського хроматину (котрі несуть на собі гетеро-
хроматинові маркери) повертаються на дочірні молекули ДНК разом
із гістонами, синтезованими de novo. НР1 також повертається на той
самий локус, де він був присутній, і відновлює патерни модифікацій
нових гістонів та компактний (репресований) стан гетерохроматино-
вої ділянки. Тобто гетерохроматиновий стан даного локусу відтворю-
ється в дочірніх клітинах. При цьому НР1 має спорідненість до білків
ламіни (розділ 1), що зумовлює розташування гетерохроматину в пе-
риферичних зонах клітинного ядра.
Подібна система репресії за участю НР1 широко використовується
в інших ділянках гетерохроматину, а також для гарантованого блоку-
вання генів в еухроматинових зонах. Метилювання Lys9 гістона Н3
і деацетильований стан лізинів гістонів Н3/Н4 є найхарактернішою
ознакою таких ділянок. Обидві модифікації, як описано вище, само-
підримуються та підтримують одна одну через опосередковану дію
НР1. При цьому важливу роль у забезпеченні репресованого стану ві-
діграє метилювання цитозинів ДНК – інша ковалентна модифікація,
яка також відновлюється при клітинному поділі й замикає своєрідне
"коло репресії" (рис. 6.10).
Генетика
182
H3/H4
Lys+
H3 Me-Lys9
ДНК 5mC
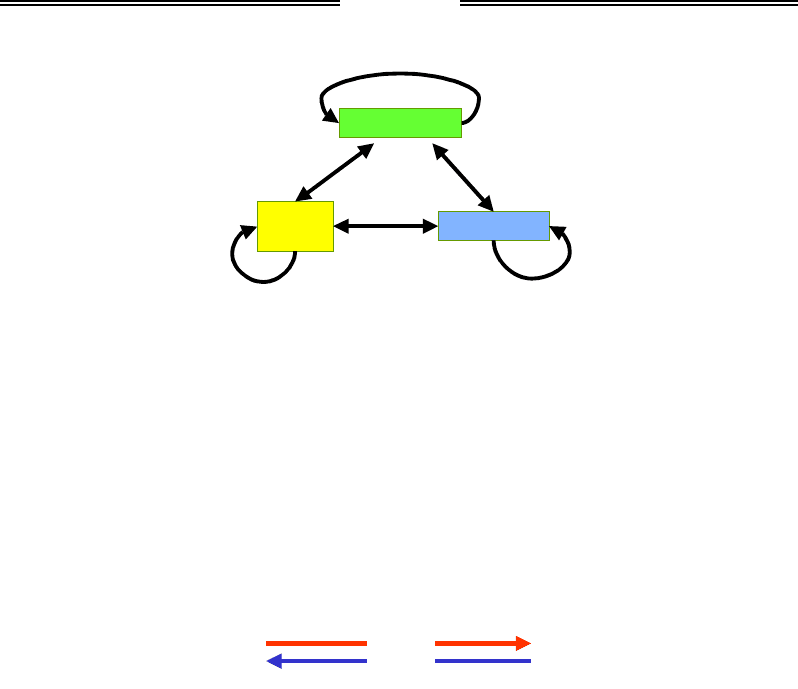
Рис. 6.10. Взаємний вплив деацетилювання гістонів,
метилювання Lys9 гістона Н3 і метилювання ДНК
у гетерохроматині. Lys+ – деацетильовані лізини
Метилювання ДНК
Патерн тканиноспецифічного метилювання ДНК (утворення
5mC у складі динуклеотидів CpG, рис. 6.11) є результатом двох
процесів: підтримання метильованого статусу після реплікації та
метилювання de novo.
pCpG
GpCp
5'
5'
3'
3'
Рис. 6.11. Динуклеотид CpG у ДНК – субстрат метилювання
Підтримуюча ДНК-метилтрансфераза спрацьовує протягом 1–2 хв
після реплікації: дві дочірні молекули ДНК містять батьківский лан-
цюг ДНК (із 5mС у складі CpG) і синтезований ланцюг, де цитозин не
є метильованим. Метилтрансфераза впізнає такі напівметильовані
динуклеотидні контакти й відновлює симетрію щодо метилювання С.
За рахунок цього процесу патерн метилювання відтворюється в дочір-
ніх клітинах, що є, поряд із відновленням модифікацій гістонів, одним
із важливих механізмів епігенетичного спадкування.
Інші ДНК-метилтрансферази здійснюють метилювання ДНК de novo.
Особливо важливим цей процес є на ранніх стадіях ембріонального роз-
витку, коли ДНК тотально деметильована. У процесі розвитку здійс-
нюється метилювання, яке визначає специфічне вимикання певних

Розділ 6. Генетика багатоклітинних еукаріотів
183
груп генів при диференціюванні клітин. Крім того, деметилювання є
можливим і в диференційованих клітинах, де метилтрансферази ви-
користовуються для відновлення метильованого статусу.
Залучення 5mС до репресії пов'язане з наявністю у складі певних
білків особливих структурних модулів – MBD (Methyl Binding D
omain),
які мають специфічну спорідненість до метильованих динуклеотидів
CpG. Білки, що містять MBD, є компонентами різноманітних репре-
суючих комплексів. Зокрема, такі білки рекрутують до метильованих
ділянок хроматину гістон-деацетилази та гістон-метилтрансферази.
І навпаки: Me-Lys9 розпізнається білком, котрий містить хромодомен
і рекрутує ДНК-метилтрансферазу (рис. 6.10).
Хоча Me-Lys9 і 5mС є загальними маркерами конститутивно реп-
ресованих ділянок хроматину, не завжди репресія та додаткова ком-
пактизація залежить від НР1. Прикладом іншої системи є інактивація
однієї з Х-хромосом у клітинах самок ссавців (див. підрозділ, присвя-
чений генетиці статі). У складі Х-хромосоми, яка буде інактивованою
(обирається випадково на ранніх стадіях розвитку), спрацьовує ген
Xist, що продукує велику некодуючу молекулу РНК. Ці РНК укривають
собою хромосому і взаємодіють з низкою білків, серед яких – гістон-
метилтрансфераза (здійснює метилювання Lys9 гістона Н3). Метил-
ювання Lys9 зумовлює метилювання ДНК, що приводить до епігене-
тичного спадкування. Крім того, до Х-хромосоми рекрутуються стру-
ктурні компактизуючі білки (але не НР1).
Гетерохроматин і РНК-інтерференція
Гетерохроматин утворюється в першу чергу на послідовностях
ДНК, що повторюються. Причому має значення не послідовність як
така, а саме наявність повторів: штучне введення повторів до ге -
номної ділянки викликає утворення гетерохроматинової зони.
У формуванні й підтриманні гетерохроматинового стану повторів
(принаймні в центромерах) важливу роль відіграє процес РНК-інтер-
ференції (див. розділ 2).
При порушенні компактизації на центромерних повторах може від-
буватися спонтанна транскрипція в різних напрямках. Оскільки матри-
цею є повтори, існує висока ймовірність синтезу комплементарних мо-
лекул РНК: утвориться дволанцюгова РНК, яка запустить процес інтер-
ференції – нуклеаза Dicer розрізає дволанцюгову РНК на фрагменти
(можуть бути далі ампліфіковані РНК-залежною РНК-полімеразою), які
залучаються до комплексу RISC. У складі комплексу РНК взаємодіють

Генетика
184
із транскриптами, і відбувається деградація останніх. Крім деградації,
інтерференція має інший наслідок: RISC, який опиняється в зоні повто-
рів, рекрутує до хроматину гістон-метилтрансферазу, яка здійснює
метилювання Lys9 гістона Н3. За вже відомою схемою (рис. 6.9, 6.10)
відбувається зв'язування НР1 і компактизація гетерохроматину.
Гетерохроматин формується також у зонах скупчення мобільних
елементів. Активність мобільних елементів може контролюватися ор-
ганізмом – відповідну систему пригнічення транскрипції мобільних
елементів, котрі використовують РНК як проміжну стадію, знайдено
як у дрозофіли, так і в мишей. Система вмикається при сперматоге-
незі та складається з білків родини Piwi й коротких регуляторних
РНК, комплементарних ділянкам мРНК, яка синтезується на мобільних
елементах. Ці РНК і білки Piwi об'єднуються в комплекс. РНК спрямо-
вує цей комплекс на мРНК мобільного елемента, білки комплексу здій-
снюють деградацію мРНК, що й запобігає переміщенню – загалом та-
ка система є аналогічною процесу РНК-інтерференції. Крім того, від
присутності згаданих регуляторних РНК залежить також інтенсивне
метилювання ДНК мобільних елементів, яке зумовлює конститутивну
репресію їхньої транскрипції.
Ефект положення гена
Описаний досить давно ефект залежності прояву гена від його по-
ложення у хромосомі має безпосереднє відношення до епігенетичного
спадкування гетерохроматинового стану.
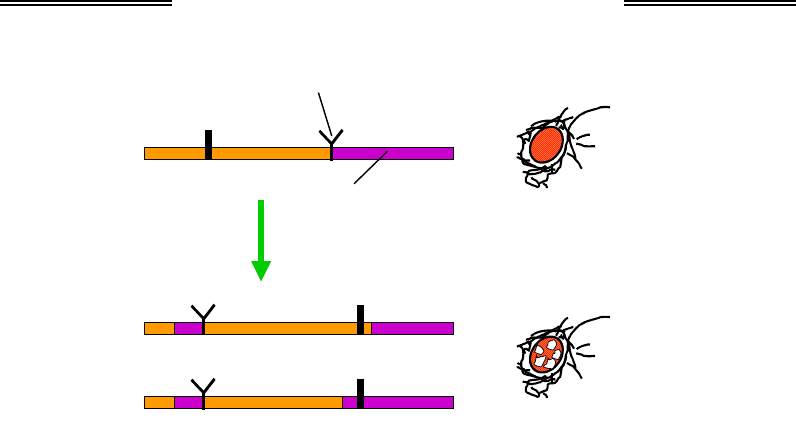
Один із прикладів ефекту положення демонструє рис. 6.12: штучно
індукована у статевих клітинах дрозофіли хромосомна перебудова з ін-
версією ділянки, яка на одному кінці містить ген w
+
(червоні очі дикого
типу), а на іншому – частину перицентромерної гетерохроматинової зо-
ни з межею, що не дозволяє гетерохроматину розповсюджуватися. Після
такої інверсії на ранніх стадіях ембріонального розвитку гетерохрома-
тин (який тепер не має межі) іноді поширюється на ген w
+
, що опинився
поряд. Це приводить до вимкнення гена, і такий залежний від структу-
ри гетерохроматину репресований стан спадкується при мітозі. У ре-
зультаті утворюються групи клітин білого кольору – мозаїчне око.
Якщо внаслідок хромосомних перебудов, нерівного кросинговеру
або активності мобільних елементів поряд із гетерохроматиновою зо-
ною опиняється група генів, то частота (імовірність) їхнього вимкнен-
ня залежить від порядку розташування – ген, що знаходиться ближче
до гетерохроматинової ділянки, вимикається частіше.
Розділ 6. Генетика багатоклітинних еукаріотів
185
w
+
w
+
w
+
Межа
Перицентромерний
гетерохроматин
Інверсія ділянки
Д
ики
й
тип
Мозаїчне
око
Рис. 6.12. Приклад ефекту положення гена –
утворення мозаїчного ока дрозофіли
ЦИТОПЛАЗМАТИЧНА СПАДКОВІСТЬ:
ГЕНЕТИКА МІТОХОНДРІЙ І ХЛОРОПЛАСТІВ
Переважна більшість еукаріотичних генів міститься в клітинному
ядрі. Крім ядра, свій власний генетичний матеріал мають цитоплаз-
матичні органели еукаріотичної клітини: мітохондрії та хлоропласти.
Геноми цих органел містять гени білків і РНК, необхідні для їхнього
функціонування. За своєю організацією геноми органел значно відріз-
няються від ядерного геному й нагадують геноми прокаріотів. Моле-
кулярна машинерія, яка забезпечує експресію генетичної інформації
в органелах, також подібна до прокаріотичної. Ці факти лягли в осно-
ву ендосимбіотичної теорії походження мітохондрій і пластид, відпо-
відно до якої органели виникли в результаті незалежних ендосимбіо-
тичних подій: вільно існуючі α-протобактерії та ціанобактерії були за-
хоплені протоеукаріотичною клітиною-хазяїном і еволюціонували
у специфічні органели (мітохондрії та хлоропласти відповідно), які
відповідають за дихання та фотосинтез. Під час коеволюції клітини-
хазяїна й ендосимбіонта частина мітохондріальних і хлоропластних
генів (інколи досить значна частина) була перенесена в ядерний ге-

Генетика
186
ном. Таким чином, наявність власного генетичного апарату забезпе -
чує мітохондріям і хлоропластам своєрідну автономність від ядерного
геному, але велика кількість компонентів, потрібних для виконання
їхніх функцій, кодується ядерним геномом.
Геноми мітохондрій
Мітохондрії, які присутні практично в усіх еукаріотів за деякими
винятками, відіграють центральну роль у синтезі АТР і деяких інших
важливих фізіологічних процесах. Кількість мітохондрій на одну клі-
тину може варіювати від декількох штук (наприклад, у сперміях)
до декількох тисяч (у гепатоцитах). Кожна мітохондрія може нести
декілька копій ДНК (мтДНК), які в комплексі з білками утворюють
структуру, подібну до нуклеоїдів прокаріотів.
Генетичний код, що використовується власною системою транс-
ляції мітохондрій, характеризується деякими відхиленнями від універ-
сальної таблиці відповідності кодонів амінокислотам (див. рис. 2.1).
Зокрема, універсальний стоп-кодон UGA в мітохондіях більшості ви-
дів кодує триптофан, у мітохондіях дріжджів кодон CUG відповідає
треоніну замість лейцину, у мітохондріях ссавців AUA відповідає ме-
тіоніну замість ізолейцину тощо.
Популярною є думка, що більшість еукаріотів мають кільцеву
молекулу мтДНК. Часто так воно і є, проте останнім часом накопи-
чилися дані про те, що у великої кількості еукаріотів мтДНК є ліній-
ною (малярійний плазмодій, гідра, деякі гриби та одноклітинні водо-
рості). Іноді, наприклад у дріжджів, лінійна молекула мтДНК являє
собою так званий конкатомер – складається з великих однакових
ділянок послідовності, що тандемно повторюються. Лінійні мтДНК
характеризуються наявністю специфічних кінцевих структур: кова-
лентно замкнені на кінцях комплементарні ланцюги, приєднані до
кінців молекули специфічні термінальні білки або теломероподібні
кінцеві повтори різноманітної довжини.
Зазвичай мітохондріальні геноми (мітохондріоми) представлені
однією "хромосомою", однак є винятки. Так, мітохондрії гриба
Spizellomyces punctatus містять три різні кільцеві молекули ДНК.
У найпростішого Amoebidium parasiticum мітохондріом складається
з декількох сотень лінійних молекул, які різняться за розмірами й по-
слідовністю. Мітохондріальний геном рослин, як правило, складається
з декількох молекул різного розміру. Одна з них, "основна хромосома",
містить більшу частину генів, а кільцеві форми меншого розміру, які
Розділ 6. Генетика багатоклітинних еукаріотів
18
7
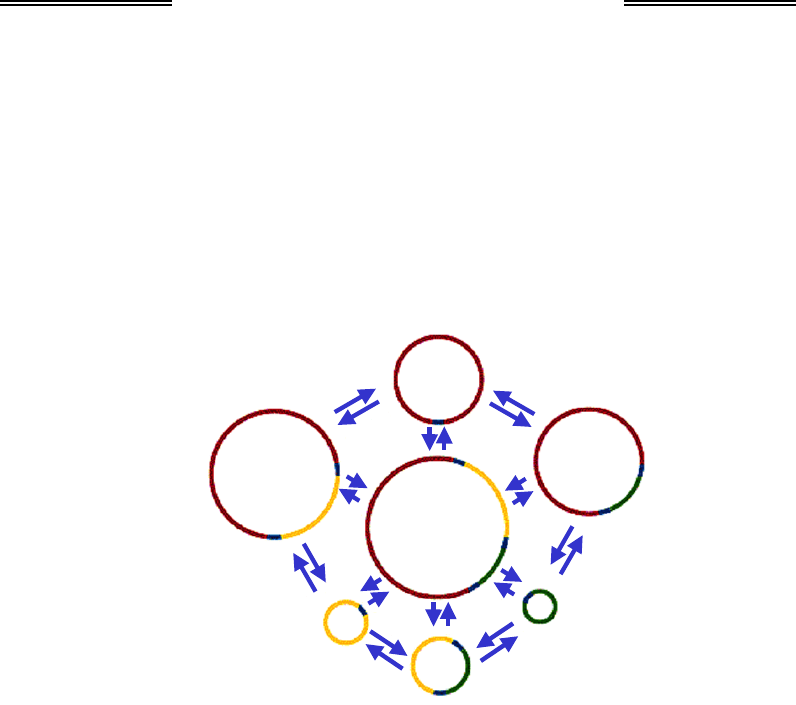
перебувають в динамічній рівновазі як між собою, так і з основною
хромосомою, утворюються внаслідок внутрішньо- та міжмолекулярної
рекомбінації завдяки наявності гомологічних ділянок (рис. 6.13). Пе-
ребудови геномів мітохондрій у результаті рекомбінаційних подій, ве-
дуть до делецій, дуплікацій, інверсій або інсерцій певних нуклеотид-
них послідовностей або цілих генів. Такі зміни можуть викликати не
тільки ушкодження наявних генів, але й появу нових працюючих ге-
нів. Вважають, що ця зміна структури мітохондріального геному кон-
тролюється ядром і є одним із механізмів регуляції ефективності екс-
пресії мітохондріальних генів.
Рис. 6.13. Схема утворення кільцевих молекул різного розміру
в мітохондріях рослин. Рекомбінація відбувається по гомологічних ділянках,
забарвлених однаковим кольором
Мітохондріальні геноми значно варіюють за розмірами: від
6 тис. пар основ у плазмодіїв до 2 млн 400 тис. пар основ у сітчас-
тої дині. У тварин мтДНК має в основному невеликі розміри – у се-
редньому 13–19 тис. пар основ. Відносно великі мітохондріоми (від
20 до 42 тис. пар основ) знайдені в молюсків, нематод і деяких ко-
мах. Мітохондріоми вищих рослини мають великі розміри (від 180
до 2 млн 500 тис. пар основ), містять значну кількість послідовнос-
тей, що повторюються, і відкритих рамок зчитування з невідомими
функціями. Характерним для мітохондріомів рослин є наявність
вбудованих ділянок хлоропластної ДНК.

Генетика
188
Набір генів у мітохондріальних геномах варіює від 5 у плазмодіїв
до 100 у джгутикового Reclinomonas americana. Для більшості еукаріо-
тів середня кількість генів, які кодує мітохондріом, становить 40–50,
з яких 12–20 є білковими. Геноми мітохондрій усіх еукаріотів кодують
рРНК великої та малої субодиниць мітохондріальної рибосоми, а та-
кож частковий (інколи повний) набір власних тРНК. Білки, що коду-
ються мітохондріомами, в основному залучені до процесів перенесен-
ня електрона та синтезу АТР: субодиниця АТР-синтази (ген atp), ком-
поненти комплексів дихального ланцюга (гени nad, sdh, cob, cox).
У мтДНК рослин і деяких одноклітинних містяться гени рибосомних
білків. Дещо атиповим за складом генів є мітохондріом дріжджів – він
містить, зокрема, гени ендонуклеаз і зворотної транскриптази.
Різниця в розмірах мітохондріомів викликана некодуючими послі-
довностями – кореляція між розміром мтДНК і кількістю генів практич-
но відсутня. Особливо велика кількість некодуючих послідовностей
мтДНК (50–70 %) є характерною для вищих рослин. Частина таких по-
слідовностей входить до складу інтронів (в основному зустрічаються в
мітохондріальних генах грибів і вищих рослин, у генах ссавців інтрони
відсутні), які видаляються з транскриптів під час сплайсингу. Більша
частина некодуючих ділянок представлена міжгенними регіонами.
Усі компоненти реплікативного, транскрипційного та частина транс-
ляційного апаратів мітохондрій кодуються ядерним геномом. Отже,
експресія мітохондріальних генів перебуває під повним контролем ядра.
Геном хлоропластів
Хлоропласти вищих рослин містять багато ідентичних кільцевих
дволанцюгових молекул ДНК, розміри яких коливаються від 120 до
220 тис. пар основ. Характерною особливістю хлоропластної ДНК
(хлДНК) вищих рослин є наявність інвертованого повтору (ІП), дов-
жина якого в середньому становить 20–30 тис. пар основ (варіює
в різних видів від 5 до 76 тис. пар основ). У результаті гени, локалізо-
вані в ІП, є дуплікованими в геномі хлоропластів. Відмінності за роз-
мірами хлДНК різних видів в основному визначаються довжиною ІП.
Винятком є хлДНК деяких бобових і хвойних, які не містять ІП. Вва-
жають, що ІП був у загального предка вищих рослин.
Як правило, ІП представлений двома сегментами (ІП-1 та ІП-2), що
розділяють хлДНК на велику й малу унікальні ділянки (рис. 6.14). Ве-
лика унікальна ділянка є найбільш варіабельною (для різних видів)
частиною молекули. У деяких видів у процесі еволюції один сегмент
ІП був повністю або частково втрачений.
Розділ 6. Генетика багатоклітинних еукаріотів
189
Мала унікальна
послідовність
ІП1
ІП2
Велика унікальна
послідовність
Рис. 6.14. Схема організації хлоропластного геному тютюну,
ІП – інвертований повтор.
Вражає подібність організації хлоропластного й бактеріального ге-
номів. Основні регуляторні послідовності, такі як промотори і терміна-
тори, в обох геномах фактично ідентичні. Білки, що кодуються хлоро-
пластними генами, дуже схожі на бактеріальні, а деякі групи генів із
близькими функціями (скажімо, гени рибосомних білків) організовані
однаково в геномах хлоропластів, Е. coli та ціанобактерій. Разом із тим,
на відміну від прокаріотів, у генах хлДНК присутні інтрони.
Велика кількість генів локалізована в кластерах, що експресуються
у вигляді великих поліцистронних первинних транскриптів. Останні
процесуються до оліго- і моноцистронних мРНК, що піддаються сплай-
сингу та редагуванню. Редагування транскриптів хлоропластних генів
виявлено тільки у вищих рослин; найчастіше спостерігаються заміни
С → U шляхом дезамінування. Аналогічний процес, що викликає замі-
ни амінокислотних залишків у відповідних білках, відбувається і в мі-
тохондріях деяких еукаріотів (найпростіших, грибів і рослин).
За складом і порядком розміщення генів хлоропластні геноми
вищих рослин є високо консервативними. У вивчених геномах хло-
ропластів вищих рослин виявлено від 108 до 122 генів, 95 з яких
є однаковими й зустрічаються в усіх видів. За функціональністю гени
хлоропластів можна розділити на три групи: гени апаратів транс-
крипції та трансляції; гени, що пов'язані з фотосинтезом; гени фото-

Генетика
190
синтетичного метаболізму (біосинтезу амінокислот, жирних кислот,
пігментів тощо). До генів першої групи відносять гени рРНК, тРНК
(близько 30), деяких субодиниць хлоропластної РНК-полімерази,
приблизно 20 генів рибосомних білків і кількох факторів трансляції.
До другої групи – гени, що кодують деякі білки, які є компонентами
фотосистем I і II, комплексу цитохромів b/f, АТР-синтази, субоди-
ниць NADH-дегідрогеназного комплексу дихального ланцюга та ве-
лику субодиницю ключового ферменту фотосинтезу рибулозобісфо-
сфаткарбоксилази. Гени третьої групи найменш вивчені. До неї
відносять ген accD, що кодує β-субодиницю ацетил-СоА-карбокси-
лази прокаріотичного типу, яка бере участь у біосинтезі жирних
кислот. Геноми хлоропластів злаків не мають гена accD. У мохопо-
дібних і голонасінних виявлено гени, пов'язані з біосинтезом хло-
рофілу. Їх було ідентифіковано за схожістю з відповідними генами
фотосинтезуючих бактерій.
Усі відомі білки, які кодуються в хлоропластах, входять до складу
великих ферментативних комплексів. Ці комплекси також містять одну
або декілька субодиниць, що кодуються ядерним геномом. Цікаво, що
субодиниці, котрі кодуються ядерним геномом, є регуляторними, а ті,
що кодуються хлДНК, – каталітичними. Усі найважливіші білки, які бе-
руть участь у реплікації, транскрипції та трансляції також кодуються
ядерним геномом. Тобто, як і для мітохондрій, усі процеси, що відбува-
ються у хлоропластах, перебувають під жорстким контролем ядра.
Основні характеристики спадкування генів органел
У диплоїдних видів еукаріотів ядерні гени зумовлюють успадку-
вання ознак за законами Менделя в результаті того, що, за винятком
зчеплених зі статтю ознак, нащадки отримують по одній копії кожно-
го гена від обох батьків. Мітохондрії та пластиди не розподіляються по
гаметах і дочірніх клітинах аналогічно хромосомам ядра, тому зако-
номірності спадкування генів органел і ознак, які вони зумовлюють,
значно відхиляються від менделівських. Оскільки мітохондрії та плас-
тиди знаходяться в цитоплазмі клітин, такий тип спадкування нази-
вається позаядерним (позахромосомним), або цитоплазматичним.
Особливості позаядерного спадкування зумовлені в першу чергу тим,
що гамети різних статей мають непорівнянні кількості мітохондрій і пла-
стид. Найчастіше жіночі гамети містять тисячі мітохондрій (і пластид
у рослин), а чоловічі – одиниці. У результаті в зиготі переважають міто-